
环形泰勒虫与牛无浆体双重PCR方法的建立
2023-04-18 03:20:24葛晓敏缪荣浩李才善陈宋琴温丽翠刘凯强郑会珍巴音查汗郭庆勇新疆农业大学动物医学学院新疆乌鲁木齐830052
中国兽医学报 2023年3期
葛晓敏,缪荣浩,李才善,陈宋琴,温丽翠,刘凯强,刘 燕,郑会珍,巴音查汗,郭庆勇 (新疆农业大学 动物医学学院,新疆 乌鲁木齐 830052)
环形泰勒虫病(theileriosis)与牛无浆体病(anaplasmosis)分别由环形泰勒虫(Theileriaannulata)和牛无浆体(Anaplasmabovis)引起,属于蜱传血液寄生虫病,这2种病原均能引起动物的高热、贫血、黄疸、消瘦及淋巴结肿大等临床症状,严重感染时可导致死亡。全球每年造成的损失估计在13.9亿~187.0亿美元之间[1]。由于环形泰勒虫病和牛无浆体病的高发病率和病死率对中国反刍动物养殖业发展造成威胁,因此,在实施适当的治疗和相关的控制措施之前,及时、具体地诊断病原体是非常重要的[2]。
传统显微镜检测法需要专业的技术人员和仪器且无法判断感染初期及携带动物是否出现感染情况[3-4]。血清学方法有较好的效果,能在寄生虫血症下降时检测疾病后期的循环抗体,然而该方法存在不同物种中抗体的交叉反应,其中ELISA检测时就可能出现假阳性[5]。基于核酸技术的检测方法即使在病原感染潜伏期也能够检测,且检测的灵敏度也高于一般检测方法,并且能够鉴定形态相似的血液寄生虫[4]。PCR是一种检测不同病原体的高灵敏度和特异性方法。目前,国内外有大量的常规PCR[6]、荧光定量PCR[7],以及二温式PCR[8]、双重PCR方法[9]等PCR方法检测蜱传病原体的报道。其中双重PCR(duplex PCR,dPCR)方法费用低且更高效省时能够在一次PCR反应体系中同时特异性地检测2种病原,可用于临床实践,便于操作。
研究表明,环形泰勒虫子孢子与裂殖体基因2(Spm2)存在于环形泰勒虫生命周期各个阶段且定位于虫体表面[10],国内外已将该抗原用于ELISA检测,且与其他表面抗原相比,Spm2ELISA对环形泰勒虫特异性更高[11-12]。此外,原核生物线粒体16Sr RNA基因高度保守,国内外也广泛应用病原LAMP和PCR等检测方法[13-15]。本研究通过环形泰勒虫Spm2和牛无浆体16S rRNA基因[15]保守序列设计并合成2对特异性引物,建立检测上述2种病原的dPCR方法。该方法有助于对病原感染初期或处于亚临床感染阶段的家畜进行检测、诊断及后续的流行病学调查研究,且有助于制定控制这2种病原感染及混合感染的策略与早期的治疗方法。
1 材料与方法
1.1 试验样品环形泰勒虫(T.annulata)、驽巴贝斯虫(Babesiacaballi)、马泰勒虫(T.equi)、牛无浆体(A.bovis)、绵羊无浆体(A.ovis)和伊氏锥虫(T.evansi)等阳性血液由新疆农业大学动物医学学院实验室保存。此外2021年4-5月在吐鲁番托克逊县采集疑似患病牛全血样品60份。
1.2 主要试剂与仪器血液DNA提取试剂盒、pEASY-T1购于北京全式金生物技术有限公司;TaqDNA聚合酶、脱氧核糖核苷三磷酸(dNTPs),DL-2 000 Marker均购于天根试剂公司;胶回收试剂盒、质粒DNA提取试剂盒,购自美国Axygen公司;DH5α感受态细胞购于TaKaRa公司。电热恒温水浴锅(型号为DK-600A)购自上海一恒科学仪器有限公司,紫外凝胶成像仪(型号为 Gel Doc2 000)、PCR仪购自美国Bio-Rad公司,紫外光度计(Nano Drop 2 000c)为美国Thermo公司产品,台式高速离心机(型号为RADIAL20)购自西班牙ORTO ALRESA公司,琼脂糖电泳仪(型号为 DYCP-31DN)购自北京六一公司。
1.3 引物设计通过DNAMAN及Premier 5.0软件设计1对引物,根据NCBI登录的环形泰勒虫Spm2基因保守序列(Y15795.1)设计引物;上、下游分别为sF:5′-AACAAGTGACCATGAGAC-3′,sR:5′-TTGTGAGCATAAGAGTTC-3′,预期扩增片段大小为853 bp,此外,合成实验室基于Tams1基因设计的环形泰勒虫引物,上、下游分别为TA-F:5′-GAGGATGAAAAGAAAAAGG-3′,TA-R:5′-GAAGTAAAGGACTG-3′;并参考文献[15]合成牛无浆体16S rRNA基因序列(MH255941.1)特异性引物,上、下游分别为rF:5′-GCGGGGGAAAGATTTATCGCTAC-3′,rR:5′-CCCTTTACGCCCAATAATTCCGA-3′,预期扩增片段大小为351 bp;引物均有上海生工生物公司合成。
1.4 全血DNA提取按照全式金血液/细胞/组织基因组DNA提取试剂盒说明书提取牛全血基因组DNA,并保存于-20℃冰箱,备用。
1.5 环形泰勒虫和牛无浆体PCR扩增及序列测定环形泰勒虫和牛无浆体PCR的反应体系为25 μL:2×EcoTaqPCR Super Mix 12.5 μL,上、下游引物(10 μmol/L)各1 μL,环形泰勒虫或牛无浆体阳性血样DNA 1.5 μL,灭菌双蒸水补足至25 μL,其反应体系见表1。
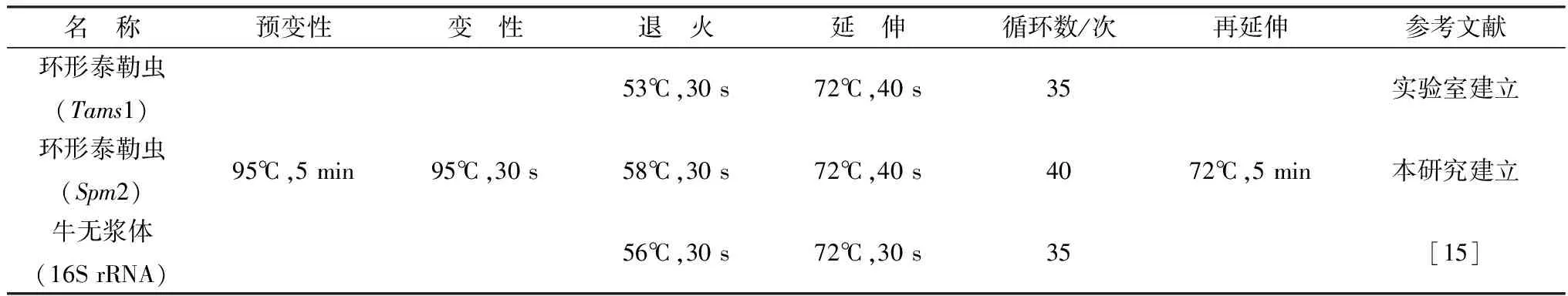
表1 PCR反应条件
扩增完成的PCR产物经琼脂糖凝胶电泳后,切下特异性条带,胶回收后连接pMD19-T载体,转化至感受态细胞大肠埃希菌(E.coli)DH5α中,将其置于含氨苄青霉素的LB固体培养基中过夜培养,挑取单菌落接种于含氨苄青霉素的LB液体培养基中培养,提取阳性质粒DNA进行PCR鉴定,并送至上海生工生物工程有限公司测序分析。
1.6 测序结果的遗传进化分析将所有测定的环形泰勒虫和牛无浆体2种病原靶基因序列在NCBI中Nucleotide Blast在线比对分析,从GenBank中下载其他已上传的相关基因序列。由于环形泰勒虫Spm2基因目前在NCBI数据库中数量较少且仅有3条上传的序列,仅能确定是否为环形泰勒虫但无法与其他种属进行种间比对,因此,本研究使用环形泰勒虫Tams1测定序列在NCBI中进行筛选同源物种基因序列构建系统进化树。采用MEGA7.0软件系统,使用Clustal W算法,设置基本参数:邻接法(Neighbor joining),Bootstrap值为1 000,以统一模型分析并分别计算以上2种病原种间遗传距离。
1.7 dPCR扩增反应体系为:2×EcoTaqPCR Super Mix 12.5 μL,上、下游引物(10 μmol/L)各1 μL,环形泰勒虫和牛无浆体阳性DNA 1.5 μL,灭菌双蒸水补足至25 μL。反应条件为:95℃ 5 min;94℃ 30 s;56℃ 30 s;72℃ 40 s,共35个循环;72℃ 5 min。
1.8 dPCR条件优化对反应体系和反应条件中的引物浓度、退火温度、循环次数进行调整。引物浓度的优化,加入引物浓度分别为环形泰勒虫1~5 μmol/L、牛无浆体2~10 μmol/L,PCR体系为25 μL,其他条件及成分不变;退火温度的优化,保持反应体系和条件不变,设置梯度退火温度51~59℃,每次递增1℃;循环次数的优化,选择优化后体系,其余条件不变,循环次数为20,25,30和35次。配制4份体系,以ddH2O作为阴性对照,为了减少误差,同时使用4台PCR仪进行扩增。
1.9 特异性试验使用方法1.1中提及的病原阳性DNA样品作为特异性试验的模板,以最佳反应体系和条件进行dPCR扩增,用于检验PCR方法的特异性。
1.10 灵敏性试验提取上述1.5的环形泰勒虫Spm2和牛无浆体16S rRNA重组质粒DNA,使用紫外光度计测定D260值,环形泰勒虫质粒DNA浓度为2.9×10-7g/μL,牛无浆体质粒DNA浓度为1.8×10-7g/μL。将2种质粒DNA按10倍梯度稀释,以此评价该方法的灵敏性。
1.11 重复性试验以环形泰勒虫和牛无浆体阳性样品为模板,用优化的dPCR反应体系和条件进行不同批次、批间重复性试验,检验该方法的重复性和稳定性。
1.12 临床样本检测从吐鲁番托克逊县随机采集经临床诊断疑似患病牛全血样品60份并提取血液DNA备用,利用该研究所建立的dPCR法、环形泰勒虫和牛无浆体已建立的单一PCR法进行检测,并对其阳性率、符合率进行统计分析。
2 结果
2.1 环形泰勒虫和牛无浆体PCR扩增对环形泰勒虫的Spm2基因保守序列和牛无浆体16S rRNA基因序列扩增产物进行凝胶电泳,结果显示,环形泰勒虫DNA扩增出853 bp的特异性条带;牛无浆体扩增出351 bp的特异性条带,与预期结果一致(图1)。
A.环形泰勒虫;B.牛无浆体;M.DL2000 DNA Marker;1~2.扩增产物
2.2 系统进化分析
2.2.1环形泰勒虫Spm2和Tams1基因的系统进化分析 结果显示,环形泰勒虫Spm2种内比对结果与印度和英国地区已上传的Spm2基因序列(Y15795、MH844677)分在一支,瑟氏泰勒虫(T.sergenti)作为外群(图2)。环形泰勒虫Tams1基因与突尼斯地区环形泰勒虫株(登录号:AF214899)分在一支,且与苏丹地区的莱氏泰勒虫株(T.lestoquardi)(KY965145、KY965146)同源关系也较近,其中马巴贝斯虫株(B.equi)作为外群(图3)。

●.本研究测定的环形泰勒虫Spm2基因

图3 环形泰勒虫Tams1基因的系统进化树
2.2.2牛无浆体16S rRNA基因的系统进化分析 通过牛无浆体16S rRNA基因同源物种基因序列进化分析,结果所示,本研究中牛无浆体16S rRNA与突尼斯和中国地区已上传的牛无浆体序列(KY655808、KM401903、KM285225、MW008784)分布在同一支[16],种间同源性100%,且与嗜吞噬细胞无浆体、绵羊无浆体、中央无浆体和边缘无浆体分在不同支上(图4)。

图4 牛无浆体16S rRNA基因的系统进化树
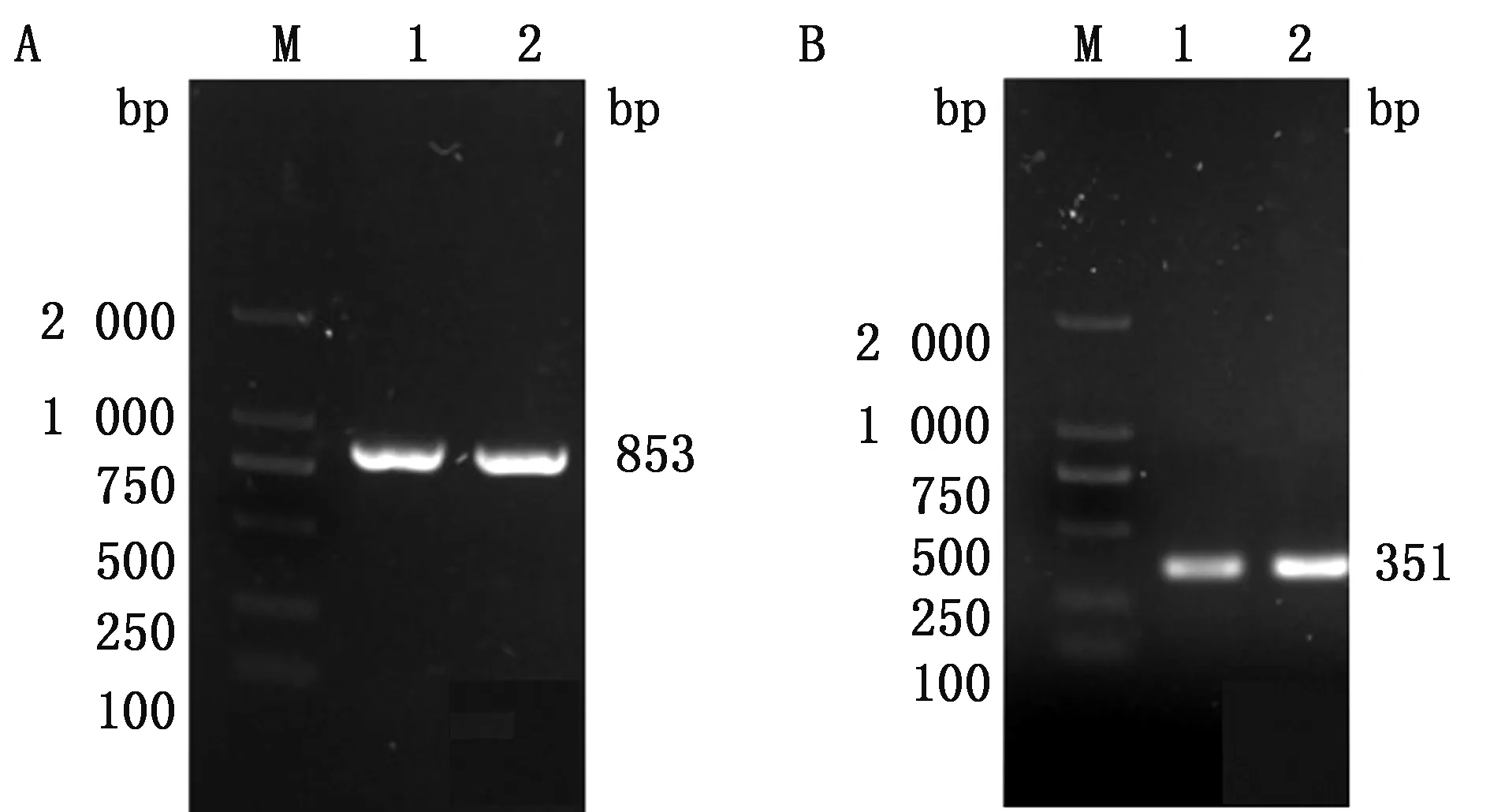
2.3 dPCR扩增同时采用环形泰勒虫sF/sR和牛无浆体rF/rR 2对引物进行PCR扩增,凝胶电泳结果显示出现853,351 bp 2条特异性条带且与预期片段大小一致(图5)。
M.DL2000 DNA Marker;1~4.扩增产物;5.阴性对照
2.4 dPCR条件优化本研究仅对影响dPCR扩增效果的引物浓度、退火温度和循环次数进行优化。
2.4.1最适引物浓度优化 通过从采用不同引物浓度(环形泰勒虫1~5 μmol/L、牛无浆体2~10 μmol/L)进行dPCR试验,结果显示,当环形泰勒虫引物浓度为5 μmol/L时、牛无浆体引物浓度为10 μmol/L时,2条目的条带最清晰(图6A)。
2.4.2最适退火温度优化 退火温度在51~59℃内进行dPCR扩增,结果显示56℃以下各个温度均能扩增2条特异性条带,当退火温度为56℃时2条带最亮(图6B)。
2.4.3最适循环次数优化 设置不同循环次数观察出现条带的亮度,以确定最适循环次数。研究结果显示循环次数为30次时2条特异性条带均最亮,所以该方法最适循环次数为30次(图6C)。

a.不同引物浓度;b.不同退火温度;c.不同循环次数;M.DL2000 DNA Marker;1~5.环形泰勒虫1~5 μmol/L和牛无浆体2~10 μmol/L;6.阴性对照;7~15.51~59℃;16.阴性对照;17~20.20,25,30和35次;21.阴性对照
2.5 优化后dPCR反应体系与条件通过对dPCR反应体系中引物剂量的优化及反应条件中退火温度、循环次数的优化,确定了双重PCR的最适反应体系与条件,见表2,3。

表2 环形泰勒虫和牛无浆体dPCR扩增体系
2.6 dPCR验证试验
2.6.1特异性试验 该研究结果显示,除混合感染阳性样品(环形泰勒虫和牛无浆体)与分别感染环形泰勒虫、牛无浆体的样品扩增出特异性条带,其他驽巴贝斯虫、马泰勒虫、绵羊无浆体和伊氏锥虫等阳性DNA样品的PCR结果均无条带,表明该方法的特异性良好(图7)。
2.6.2灵敏性试验 将环形泰勒虫Spm2质粒DNA浓度(2.9×10-7g/μL)和牛无浆体16S rRNA质粒DNA浓度(1.8×10-7g/μL)以10倍梯度稀释后进行双重PCR扩增试验,结果显示,随稀释梯度增加条带亮度减弱,其中环形泰勒虫最低检出浓度为2.9×10-16g/μL;牛无浆体最低检出浓度为1.8×10-19g/μL(图8)。

M.DL2000 DNA Marker;1~9.2.9×10-8~2.9×10-16g/μL;1~12.1.8×10-8~1.8×10-18g/μL

表3 环形泰勒虫和牛无浆体dPCR扩增条件
2.6.3重复性试验 使用本研究建立的dPCR方法对混合感染阳性样品进行3次重复试验,每次批间试验设置3个重复进行批内重复试验,重复性试验结果均一致,表明该方法具有良好的重复性与稳定性(图9)。

M.DL2000 DNA Marker;1~3.扩增产物;4.阴性对照
2.6.4临床样本检测 对60份疑似血样分别进行单一PCR法和本研究建立的dPCR法进行检测,结果显示单一PCR检测环形泰勒虫的阳性率为66.7%(40/60),牛无浆体阳性率为46.7%(28/60),环形泰勒虫和牛无浆体混合感染率为36.7%(22/60)。dPCR检测环形泰勒虫阳性率为66.7%(40/60),牛无浆体阳性率为53.3%(32/60),混合感染阳性率为43.3%(26/60);其中单一PCR和dPCR检测以上2种病原出现单一或混合感染的阳性符合率为93.3%(56/60),见表4。

表4 2种方法对临床样本检测阳性率统计%
3 讨论
环形泰勒虫病和无浆体病是世界热带和亚热带地区牛的两大蜱传疾病,其传播媒介均为璃眼蜱属,这些病原可在牛体内同时感染,使疾病诊断和治疗变得困难[17-19]。动物急性感染环形泰勒虫和/或牛无浆体恢复后仍持续感染,成为携带者并在蜱传播感染过程中发挥重要作用[3]。目前这2种病原在检测和诊断上一般都采用简便、敏感性和特异性较高的分子生物学方法,相比于普通血液涂片法和血清学检测来说减少了假阳性与操作的复杂性等问题[20,21]。
据文献报道,环形泰勒虫Spm2基因存在于虫体各个周期,是环形泰勒虫的保守基因[11,22]。因此本研究选取环形泰勒虫Spm2基因并参考文献选取牛无浆体16S rRNA基因[15],2种基因序列均是病原的保守序列,建立一种可以同时检测这2种病原的dPCR方法。根据环形泰勒虫Spm2、Tams1和牛无浆体16S rRNA基因构建了系统进化发育树,结果显示,本研究测序的环形泰勒虫Spm2基因序列与NCBI中印度和英国地区已上传的同基因序列(登录号:Y15795、MH844677)分在一支;环形泰勒虫Tams1基因序列与突尼斯地区已上传的同基因序列(登录号:AF214899)分在一支,据文献报道环形泰勒虫Tams1基因与莱氏泰勒虫裂殖子表面抗原(Ms1)基因具有显著相似性[23],因此本研究测定的Tams1基因序列与苏丹地区莱氏泰勒虫株(T.lestoquardi)同源性很近分在同支。牛无浆体16S rRNA基因与突尼斯和中国地区已报道的牛无浆体序列(KY655808、KM401903等)分在一支,同源性为100%,且与其他无浆体遗传距离较远。
dPCR仅扩增出2条目的条带,其他驽巴贝斯虫、马泰勒虫、绵羊无浆体和伊氏锥虫均无非特异性扩增及交叉反应情况,表明该检测方法的特异性良好。其中环形泰勒虫和牛无浆体最低检出浓度分别为2.9×10-16,1.8×10-19g/μL,其灵敏度较已发表基于16S rRNA基因的牛无浆体PCR检测方法略高[10]。对本研究建立的dPCR方法进行组内、组间3次重复性试验,结果表明该方法具有良好的稳定性,可用于临床检测试验。利用本研究建立的dPCR方法检测吐鲁番托克逊县60份牛全血DNA样品。结果显示,dPCR与本实验室建立的环形泰勒虫单一PCR和已发表的牛无浆体单一PCR[15]相比,dPCR检测牛无浆体的敏感性高于单一PCR,而与单一PCR检测环形泰勒虫敏感性相似。
目前,许多不同的PCR方法已被用于物种水平上的病原体鉴定,但在同一物种上检测超过一种病原时其操作过程就相对繁琐。本研究建立的dPCR方法可提高检测环形泰勒虫和牛无浆体的效率且能检测家畜出现2种病原混合感染情况。据报道,印度学者建立了牛巴贝斯虫(B.bovis)和环形泰勒虫(T.annulata)的双重PCR方法[3],证实了当地环形泰勒虫和牛巴贝斯虫存在混合感染情况,而在新疆地区尚未有报道这2种病原之间存在混合感染,今后在当地进行流行病学调查时可增加这方面的研究并探索建立一种可同时检测环形泰勒虫、无浆体和牛巴贝斯虫等多种病原的多重PCR方法。
本研究成功建立了具有快速、高效、简便及良好的特异性和灵敏性等特点的环形泰勒虫和牛无浆体dPCR检测方法。
