
基于非靶向代谢组学分析黄桃果肉褐变过程中代谢产物的差异
2023-03-23 06:22龚意辉李丽梅田宗琼曾永贤陈致印
中国食品学报 2023年2期
龚意辉,李丽梅,黄 华,李 论,田宗琼,张 娟,曾永贤,陈致印*
(1 湖南人文科技学院 农业与生物技术学院 湖南娄底 417000 2 广东省农业科学院果树研究所 农业农村部南亚热带果树生物学与遗传资源利用重点实验室 广东省热带亚热带果树研究重点实验室 广州 510640)
黄桃(Amygdalus persica)是湖南省最重要的特色水果之一,富含大量的番茄红素、番茄黄素、胡萝卜素、维生素C、膳食纤维等营养物质[1],具有果实大、果型优美、色泽金黄、肉质鲜美、口味香甜等特点,其营养和经济价值都很高,深受国内外广大消费者的欢迎。近年来,黄桃种植已成为湖南省的重点支柱产业和扶贫产业,是促进各县市农民产业增收的重要手段。然而,黄桃采收时正处于高温盛夏季节,采摘后的黄桃果实因代谢旺盛而易发生果实软化,从而引发果肉褐变和异味现象的发生,导致黄桃果实品质和市场价值下降,严重制约了我省黄桃产业的进一步发展[2]。
代谢组学(Metabonomics)是生物组学中发展出来的一门新兴学科,是利用高通量化学分析技术对某一生物体细胞、组织中所有小分子代谢产物进行定性和定量研究的一门新学科,是解决诸多果蔬采后病害问题的有效工具之一[3-4]。其中,非靶向代谢组学(Untargeted metabolomics)能全面反映生物样本中的代谢物信息,有助于挖掘生物样本新的代谢通路等[5]。代谢组学技术广泛应用于植物代谢及逆境响应机理[6],营养科学[7],采后病害诊断[8],品种鉴定[9]等方面。Busatto 等[10]利用靶向代谢组学技术(Targeted metabolomics)鉴定了苹果虎皮病发生过程中的代谢物差异,发现表儿茶素、儿茶素、绿原酸等代谢物与虎皮病的形成有着密切的关系。Lee 等[11]利用LC-MS 技术研究了二苯胺(DPA)处理的苹果贮藏期间各代谢物含量的变化规律,表明乙醛、乙酸乙酯和乙醇等挥发性代谢物的积累与果肉褐变有着密切的关系。Pedreschi等[12]利用LC-MS 技术解析贵妃梨气调贮藏过程中果心褐变的发生机理,发现葡萄酸和GABA 可能是果心褐变和坍塌的代谢标志物。Gong 等[13]基于非靶向代谢组学技术在苹果果皮中共鉴定出833 种代谢产物,其中59 种显著差异代谢参与虎皮病的形成。目前,国内外学者从酶促褐变、乙烯合成等角度深入研究了桃果肉褐变机理[14-15],然而,利用代谢组学技术分析采后黄桃果肉褐变进程中的代谢物质目前尚无研究报道。本研究采用LC-MS/MS 技术,以黄桃果实为试验材料,利用非靶向代谢组学技术对采后黄桃果肉褐变进程中的代谢物进行研究,以探明黄桃果肉褐变过程中的关键代谢物,为进一步完善黄桃果肉褐变机理及其控制提供理论参考。
1 材料与方法
1.1 试验处理
以采自冷水江市水云峰基地的黄桃果实为试验材料,所采摘果实的生长环境和栽培管理一致,挑选无病虫害、果形和色泽一致的果实。随机挑选30 个黄桃果实为一组,装入PE 薄膜袋中,放入温度为(20±1)℃,且湿度为85%的恒温箱中贮藏12 d。以无处理的黄桃果实作为对照组(CK),在同样条件下贮藏12 d,并进行3 次生物学重复。分别在果实贮藏0,3,6,9,12 d 后切取1 cm 厚的果肉,用液氮速冻后立即放在-80 ℃的超低温冰箱中保存,用于后续样品的LC-MS/MS 分析。
1.2 主要试剂与仪器
水,德国Merck 公司,醋酸铵、甲酸、甲醇等质谱试剂,美国Thermo Fisher 公司,PE 薄膜袋(规格为200 mm×200 mm,厚度为0.05 mm),广州健阳生物科技有限公司。
D3024R 低温离心机,美国Scilogex 公司,Vanquish UHPLC 色谱仪、Q ExactiveTMHF-X 质谱仪,德国Thermo Fisher 公司,Hypesil Gold column 色谱柱(100 mm×2.1 mm,1.9 μm),美国Thermo Fisher 公司等。
1.3 方法
1.3.1 褐变指数的测定 参照李丽梅等[16]的方法。将黄桃果实中间部位纵切成两半,按照黄桃果肉褐变程度进行分级。0 级:果肉完全无褐变;1级:1%~10%面积果肉轻微褐变;2 级:10%~30%的褐变面积;3 级:30%~50%的褐变面积;4 级:果肉褐变面积>50%。
褐变指数=Σ(褐变级别×该级果实数占总黄桃数的百分比)。
1.3.2 黄桃果肉代谢物的提取 根据Want 等[17]的方法,称100 mg 黄桃果肉样品,立即加入液氮速冻后研磨成粉末,加500 μL 80%甲醇用振荡器混匀,将黄桃果肉代谢物提取液放在冰上静置5 min,4 ℃、15 000×g条件下离心20 min 取上清液,加质谱级水将甲醇体积分数稀释至53%;将样品在4 ℃、15 000×g条件下离心20 min 取100 μL 的上清液,全部用0.22 μm 的有机相过滤后转入LC进样瓶中,以在每个提取液中吸取30 μL 混匀作为质控QC 样本,以53%甲醇作为空白对照,与试验样品进行同样的操作,将各提取液记录好并放入超低温冰箱中保存,用于后续质谱数据的采集。
1.3.3 色谱条件 柱温:40 ℃;流速:0.2 mL/min;正离子模式:流动相A:0.1%甲酸,流动相B:甲醇;负离子模式:流动相A:5 mmol/L 醋酸铵(pH 9.0),流动相B:甲醇;进样体积:100 μL;按照体积比进行梯度洗脱,洗脱程序为:0~1.5 min,2%~2% B;1.5~14 min,2%~100% B;14~17 min,100%~2% B。
1.3.4 质谱条件 正负离子模式扫描范围m/z100~1 500;质谱电压3 200 V,离子源温度320℃,GSⅠ和GSⅡ分别为276 000 Pa 和69 000 Pa,采用数据依赖型串联质谱法进行MS/MS 二级扫描。
1.3.5 代谢物定性与定量 利用Compound Discoverer 3.1 软件对黄桃果肉样本的下机数据(.raw)文件进行预处理,首先依据每种代谢物的保留时间、分子质量、质荷比等进行初步筛选,然后按照质量偏差和保留时间偏差在不同果肉样本比较组中进行峰对齐,再与北京诺禾致源科技股份有限公司自建代谢物数据库、mzCloud(https://www.mzcloud.org/)、mzVault 等数据库进行匹配,鉴定黄桃果肉中的代谢物。最后对果肉样品中鉴定到的色谱峰进行积分,每个色谱峰对应一种代谢物,其相对含量用峰面积表示。
1.3.6 数据分析 使用Compound Discoverer 3.1软件对黄桃果肉褐变中所鉴定的代谢物数据进行相对定性和定量分析。采用主成分(PCA)、正交偏最小二乘判别分析(Orthogonal partial least squares discrimination analysis,OPLS-DA)等方法分析了黄桃各果肉样本中的显著差异代谢物。根据OPLS-DA 法判断模型稳定性和准确性。利用多维统计VIP 值、差异倍数(Fold change,FC)以及q值分析黄桃果肉褐变中的显著代谢物,显著代谢物满足FC≥1.2 或≤0.8333,且q<0.05 两个条件。通过Heml1.0 程序对黄桃果肉褐变进程中的显著差异代谢物进行聚类分析并绘制热图。利用HMDB 数据库(https://hmdb.ca/ metabolites)和KEGG 数据库(https://www.genome.jp/kegg/pathway.html)对黄桃果肉褐变过程中的代谢物进行通路分析。
2 结果分析
2.1 PE 包装对果肉褐变的影响
在采后黄桃果实贮藏期间,PE 包装和对照组的果肉褐变指数均不断增加,PE 包装处理的黄桃果肉在第3 天没有出现果肉褐变的现象,PE 包装的果肉褐变指数始终低于对照组(图1),说明PE包装处理明显抑制黄桃果肉褐变的发生。
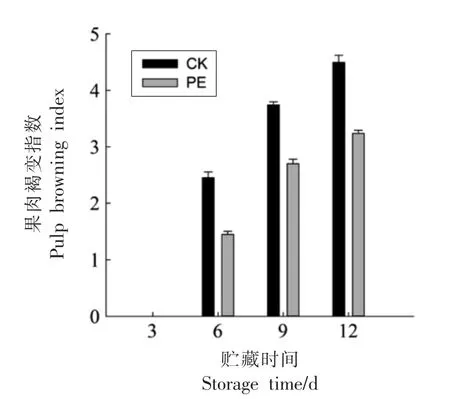
图1 PE 包装对果肉褐变指数的影响Fig.1 The effect of PE packaging on pulp browning index
2.2 差异代谢物筛选结果
基于北京诺禾致源科技股份有限公司自建代谢物数据库、mzCloud、mzVault 等数据库,对黄桃果肉褐变过程中的代谢物进行定性和定量分析。表1 显示,共在黄桃果肉中检测出991 种代谢化合物,在CK 12 d vs CK 0 d 样品中共筛选出232种显著差异代谢物,相对含量显著上调、下调的代谢物分别为206 种和26 种;在PE 12 d vs CK 0 d 样品中共筛选出203 种显著差异代谢物,相对含量显著上调、下调的代谢物分别为145 种和58种;而在其它果肉样本中,显著差异代谢物总数相对较少。
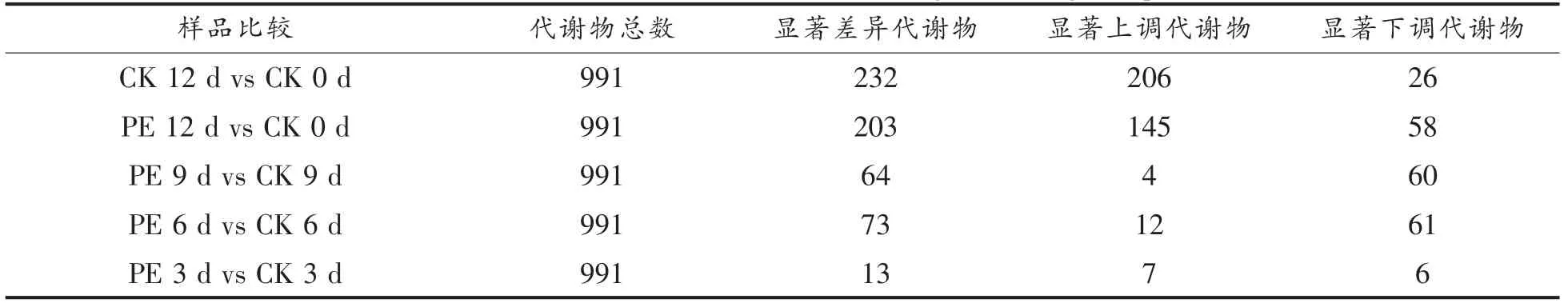
表1 黄桃果实贮藏过程中差异代谢物的筛选Table 1 Identification differential metabolites during the storage of peach fruit
2.3 PCA 分析
分别对CK 12 d vs CK 0 d 和PE 12 d vs CK 0 d 样本进行主成分(PCA)分析,以阐明褐变严重的果肉(CK 12 d)、褐变中等程度的果肉(PE 12 d)和无褐变(CK 0 d)各分间和组内样品之间的变异度大小。PCA 结果显示,在CK 12 d vs CK 0 d 样本中,PC1、PC2 的贡献率分别为46.69%,23.85%,CK 12 d vs CK 0 d 样本分离较为明显,组内之间褐变严重的果肉样品分离小于无褐变的果肉样品,PCA 结果说明在CK 12 d vs CK 0 d样品比较对中的代谢物差异较大(图2a)。而在PE 12 d vs CK 0 d 样本中,PC1、PC2 的贡献率分别为43.35%,23.23%,且PE 12 d vs CK 0 d 样本在二维图上呈现出明显的分离趋势,组内之间褐变中等程度的果肉样品分离小于无褐变的果肉样品,PCA 结果说明:在PE 12 d vs CK 0 d 样本中的代谢物存在较大的差异(图2b)。

图2 样品组间的PCA 得分图Fig.2 PCA results of samples
2.4 OPLS-DA 分析
本研究采用的偏最小二乘法判别分析(OPLS-DA)能实现黄桃果肉样品组间区分的最大化,便于筛选黄桃果肉褐变中的差异代谢物。利用OPLS-DA 模型分析了黄桃果肉褐变发生过程中991 种代谢物,没有褐变的果肉样品(CK 0 d)和褐变程度严重的果肉样品(CK 12 d)分别位于置信区间左侧和右侧,并且CK 0 d 和CK 12 d 样本间具有明显的区分度(图3a)。PC1 和PC2 的贡献率为56.28%和10.77%,R2Y=1、Q2Y=0.91,证实了OPLS-DA 模型比PCA 分析黄桃果肉褐变差异代谢物效果更佳(图3a)。进一步对该模型进行了200 次排列试验验证,根据200 次打乱并建模后的Q2值和R2值可以得到它们的回归线,R2=0.84大于Q2=-2.22,并且Q2回归线与Y轴截距小于0,表明该模型没有“过拟合”,能准确地描述样本的信息(图3b)。没有褐变果肉样品(CK 0 d)和中等褐变程度果肉(PE 12 d)分别位于置信区间左侧和右侧,且CK 0 d 和PE 12 d 区分明显。PC1、PC2 的贡献率分别为52.24%,17.82%,R2Y=0.99、Q2Y=0.81(图3c)。同样对该模型进行了200 次的排列试验验证,R2=0.88 大于Q2=-2.09,同样说明该模型未“过拟合” 且更好地分析果肉样品信息(图3d)。

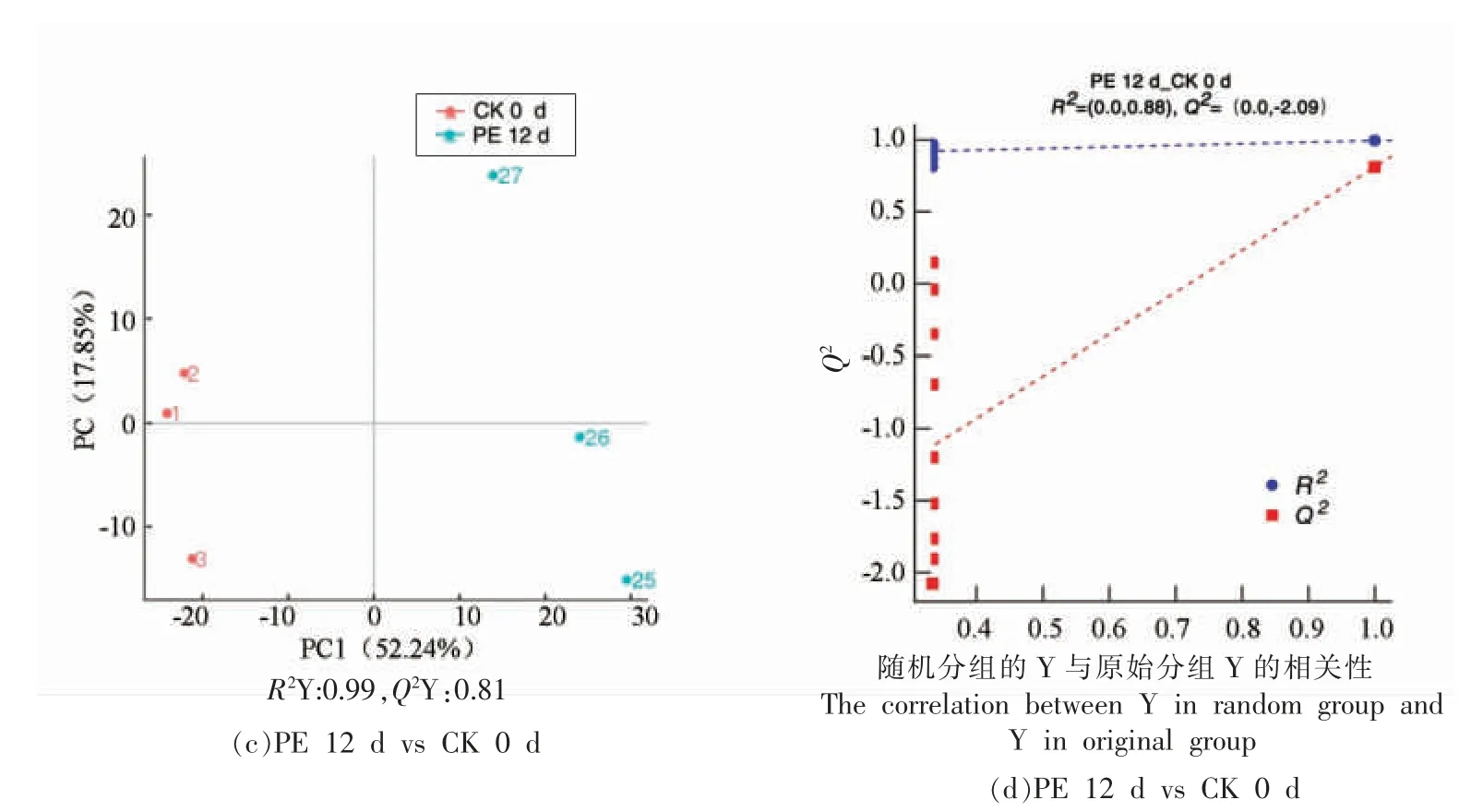
图3 不同黄桃果肉OPLS-DA 得分图及置换图Fig.3 OPLS-DA score chart and replacement chart of different yellow peach pulp
2.5 差异代谢物火山图
分别对CK 12 d vs CK 0 d 和PE 12 d vs CK 0 d 两组样本进行火山图分析(图4),红、绿和灰3 种颜色分别表示代谢物的相对含量呈显著上调、显著下调、无显著变化,表明CK 12 d vs CK 0 d 和PE 12 d vs CK 0 d 两组样本中大部分代谢物无显著变化,少数代谢物相对含量呈显著上调或下调。

图4 正离子模式下的组间火山图Fig.4 Volcano map of two groups of samples in positive ion mode
2.6 差异代谢物聚类分析
采用聚类分析方法对黄桃果肉贮藏过程中的差异代谢物进行分析,图中颜色表示果肉中代谢物的相对含量,颜色越红则表示果肉中代谢物的相对含量越高,越蓝则表示果肉中代谢物的相对含量越低。图5 和图6 形象说明了代谢物在黄桃果肉褐变过程中的变化规律。在CK 12 d vs CK 0 d 样品中,在232 种显著差异代谢物中,果肉褐变严重的(CK 12 d)相对无褐变的果肉样品(CK 0 d)有206 种代谢物显著上调,显著上调代谢物占232 种差异代谢物的88.79%,包括紫杉醇A、纳地西酮、4-苯基丁酸、2-苯基苯酚等206 种代谢物含量显著上调,然而儿茶素、表儿茶素、原花青素B2、黄芩素B、烟酸等26 种代谢物的含量呈显著下调,仅为显著代谢物总数的11.21%(图5)。而在PE 12 d vs CK 0 d 果肉样品中,共筛选出显著代谢物203 种,褐变程度中等的果肉样品(PE 12 d)相比无褐变的果肉样品(CK 0 d),有春日霉素、蝶呤B、青蒿素、脱落酸葡萄糖酯等145 种代谢物含量显著上调,占显著代谢物总数的71.43%,而哌可酸、醌、黄嘌呤酸等58 种化合物的含量呈显著下调,占总显著代谢化合物的28.57%(图6)。

图5 正离子模式CK 12 d 与CK 0 d 差异代谢物聚类分析热图Fig.5 The differential metabolites between CK 12 d and CK 0 d in positive ion mode performed in heat map
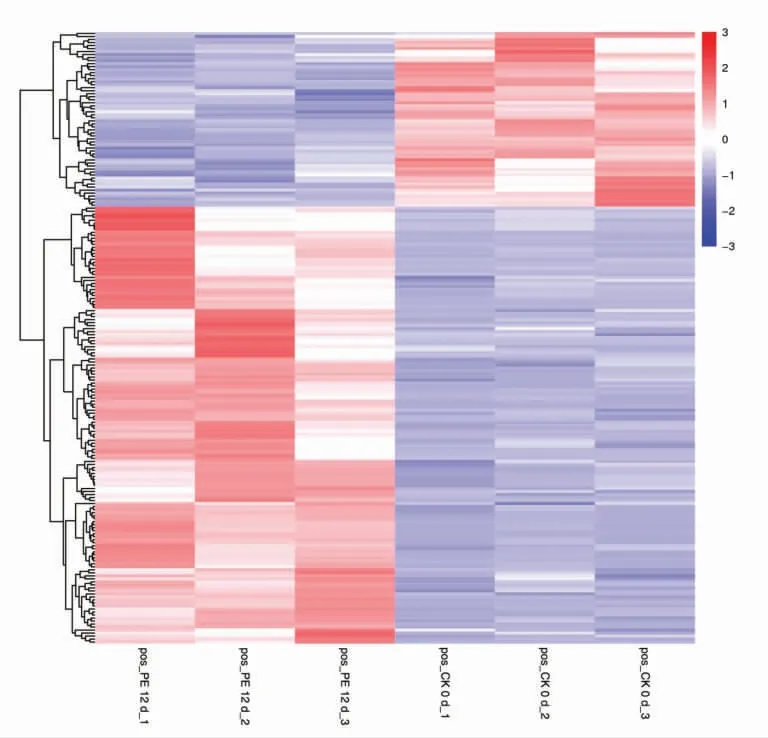
图6 正离子模式PE 12 d 与CK 0 d 差异代谢物聚类分析热图Fig.6 The differential metabolites between PE 12 d and CK 0 d in positive ion mode performed in heat map
2.7 差异代谢物通路分析
2.7.1 HMDB 通路分析 利用Human Metabolome Database 数据库对黄桃果肉褐变过程中的显著代谢物进行HMDB 通路分析(图7)。黄桃果肉褐变进程中的显著代谢物主要注释到13 条HMDB 通路中,其中显著代谢物数量分布排名前6 条代谢途径分别为:①苯丙烷和聚酮化合物代谢通路(Phenylpropanoids and polyketides),包括香豆素、槲皮素、芦丁、儿茶素、三甲氧基肉桂酸等75 种化合物;②脂质和类脂质分子(Lipids and lipid-like molecules),包括D-泛醇、皮质醇、香芹酮、十六酰胺、麦芽糖醇、硬脂酰胺等71 种化合物;③有机杂环化合物(Organoheterocyclic compounds),主要包括间苯三酚、烟酸、烟酰胺、鸟嘌呤、1-甲基腺嘌呤、3-吲哚丙酸、维生素B2、甾胆素等53 种化合物;④有机酸及其衍生物(Organic acids and derivatives),主要包括L-苯丙氨酸、脯氨酸、L-焦谷氨酸、哌啶酸、L-酪氨酸、L-苏氨酸、胱硫醚、肌酸、精氨琥珀酸等50 种化合物;⑤苯甲酸酯类(Benzenoids),主要包括香兰素、丁香酸、没食子酸、丁香酚、4-乙基苯酚、原儿茶酸等40 种化合物。⑥有机氧化合物(Organic oxygen compounds),主要包括葡萄糖1-磷酸、L-艾多醇、N-乙酰-D-半乳糖胺、胞壁酸、苯乙酮等25 种化合物。
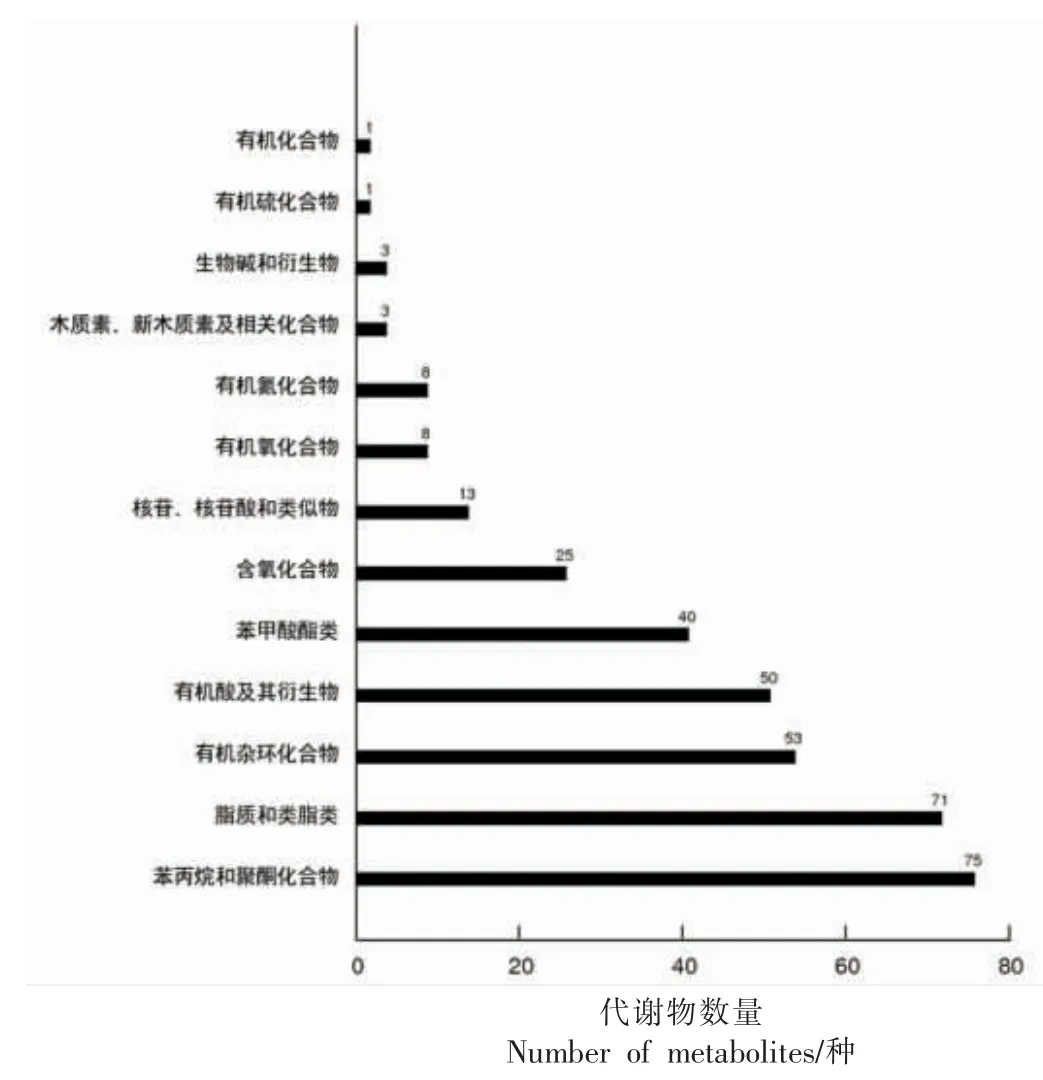
图7 黄桃果肉中差异代谢物的HMDB 富集分析Fig.7 Differential metabolites of yellow pulp in HMDB enrichment map
2.7.2 KEGG 通路分析 利用KEGG 数据库对黄桃果肉褐变中的差异代谢物质进行通路注释分析结果(图8)显示,将黄桃果肉褐变代谢物分为3大类:1)环境信息加工(Environmental information processing),包括信号转导(Signal transduction)和跨膜运输(Membrane transport)两个亚类,其中水杨酸、吲哚-3-乙酸、肌醇参与信号转导途径,维生素B1、L-苏氨酸、L-苯丙氨酸等10 种化合物参与跨膜运输代谢通路;2)遗传信息加工(Genetic information processing),包括翻译(Translation)和折叠、分类及降解(Folding, sorting and degradation)两个亚类,其中L-天冬酰胺、L-酪氨酸、L-谷氨酸等7 种化合物参与翻译通路,S-腺苷甲硫氨酸、维生素B1参与折叠、分类及降解通路;3)新陈代谢(Metabolism),包括其它次生代谢物的生物合成通路(Biosynthesis of other secondary metabolites)、氨基酸代谢通路(Amino acid metabolism)、辅因子和维生素的代谢通路(Metabolism of cofactors and vitamins)、其它氨基酸的代谢通路(Metabolism of other amino acids)、脂肪酸代谢(Lipid metabolism)等10 个二级分类,其中绿原酸、酪胺、羽扇豆碱等44 种化合物参与其它次生代谢物的生物合成通路,原儿茶酸、D-脯氨酸、酪胺香兰素等38 种化合物参与氨基酸代谢途径,泛酸、烟酰胺、生物素等28 种化合物参与辅因子和维生素代谢途径,L-焦谷氨酸、亚精胺、精胺等14种化合物参与其它氨基酸的代谢途径,花生四烯酸、亚油酸、磷胆碱等12 种化合物参与脂肪酸代谢途径。
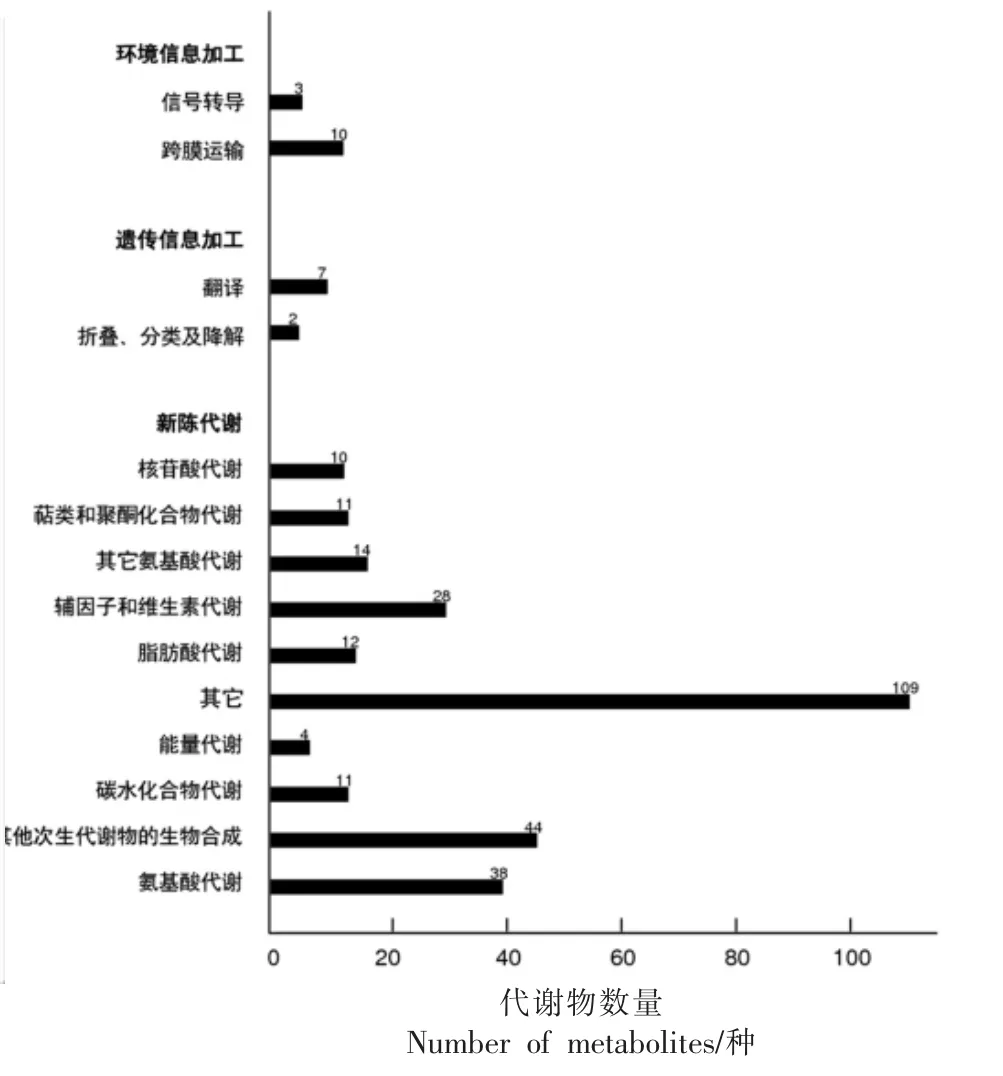
图8 黄桃果肉中差异代谢产物的KEGG 富集分析Fig.8 Differential metabolites of yellow pulp in KEGG enrichment map
3 讨论
近年来,国内外学者利用HPLC 技术对桃果实中的酚类物质鉴定进行了大量的研究,发现桃果肉中存在大量的酚类化合物,且种类较多[18-19]。Zhang 等[20]利用HPLC 技术鉴定出表儿茶素、芦丁、绿原酸、儿茶素、根皮苷等化合物是组成桃果肉主要的酚类物质。不同色泽桃果肉中富含酚类物质的种类和含量有较大的差别。表儿茶素、绿原酸、儿茶素和新绿原酸是红肉桃主要富含的酚类成分,表儿茶素、绿原酸和儿茶素是黄肉桃主要富含的酚类成分,芦丁、新绿原酸和儿茶素则组成了白肉桃主要的酚类成分[21-22]。这些研究表明,利用HPLC 鉴定果实中代谢化合物种类有限。本研究主要利用非靶向代谢组学技术对采后黄桃果肉褐变发生过程中的代谢物进行了分析,共在黄桃果肉中鉴定出991 种代谢物质,大多数代谢物的相对含量在果肉褐变过程中没有显著变化,只有少数代谢物的相对含量显著上调或下调。例如共在CK 12 d vs CK 0 d 样本中确定232 种显著代谢物,有206 种化合物显著上调和26 种化合物显著下调。方贤胜等[23]利用广靶代谢组学技术在浅黄色和紫色核桃内种皮中检测出680 种次生代谢物。Gong 等[13]利用非靶向代谢组学技术在蛇果和科特兰苹果虎皮病发生过程中共鉴定出833 种代谢化合物,共筛选出59 种差异代谢物。Doppler 等[24]利用非靶向代谢组学方法分别在小麦穗、茎、叶、根组织中检测出871,785,733,517 种代谢物。这些结果说明,非靶向代谢组学技术相比HPLC 技术具有鉴定代谢物种类多、准确度、高效等优势。
果蔬褐变与组织中酚类底物种类及其相对丰度关系密切。目前大多数学者认为,果实酚类底物的含量随着果实褐变症状的严重而表现出降低的趋势,是因为这些酚类底物被PPO 催化氧化聚合形成褐色物质,从而使得果实褐变的症状表现出来。本研究非靶向代谢组学技术在黄桃果肉褐变过程中发现儿茶素、表儿茶素、绿原酸、原花青素B2、黄芩素B、烟酸等26 种代谢物的相对含量显著下调,暗示这些物质可能作为黄桃果肉褐变的底物,被PPO 催化氧化消耗,加速了黄桃果肉褐变的进程。武倩[25]在深州蜜桃果肉中发现褐变指数与酚类物质的含量变化呈显著对应关系。有学者发现儿茶素、表儿茶素、绿原酸的含量均随着桃果肉褐变症状的加深而表现出不断下降的趋势[26-27]。Duan 等[28]提取和纯化了桃果肉中部分PPO,并利用HPLC 技术分离出PPO 的天然底物,将PPO 与底物混合后立即形成褐色,结果表明了PPO 能催化氧化酚类物质导致果肉褐变的发生。这些研究表明,引起黄桃果肉褐变的因素很多且复杂,跟果肉中代谢物的种类、相对含量、酶促褐变等因素密切有关。
4 结论
本文建立了基于LC-MS/MS 对黄桃果肉褐变过程中差异次生代谢物的非靶向代谢组学分析方法。在黄桃果肉中共鉴定出991 种代谢物,在CK 12 d vs CK 0 d 样品中,共有206 种代谢物显著上调,包括紫杉醇A、纳地西酮、4-苯基丁酸、2-苯基苯酚等206 种代谢物含量显著上调,而儿茶素、表儿茶素、原花青素B1、黄芩素B、烟酸等26 种代谢物的含量显著下调,这些代谢物质可能作为果肉褐变的底物,被逐渐氧化消耗,暗示这些代谢物可能参与黄桃果肉的褐变进程。本研究在黄桃果肉褐变过程中所鉴定的关键代谢物可为果肉褐变控制技术的研发提供理论支撑。
猜你喜欢
农业工程学报(2022年6期)2022-06-27
食品安全导刊(2021年20期)2021-08-30
小学生学习指导(低年级)(2021年5期)2021-07-21
小学生学习指导(中年级)(2021年5期)2021-05-18
小学生学习指导(高年级)(2021年5期)2021-05-18
农产品市场周刊(2019年15期)2019-09-24
江苏农业科学(2016年9期)2016-11-28
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
茶叶通讯(2014年2期)2014-02-27
食品科学(2013年6期)2013-03-11
