
栽培模式对稻茬小麦籽粒产量、氮素吸收利用和群体质量的影响
2023-03-09 08:40:48丁锦峰徐东忆丁永刚朱敏李春燕朱新开郭文善
中国农业科学 2023年4期
丁锦峰,徐东忆,丁永刚,朱敏,李春燕,朱新开,郭文善
扬州大学农学院/江苏省作物遗传生理国家重点实验室培育点/粮食作物现代产业技术协同创新中心/扬州大学小麦研究中心,江苏扬州 225009
0 引言
【研究意义】我国小麦生产能力不断提升,保障了粮食安全,但仍面临肥料施用过多、氮肥利用率偏低、籽粒品质不稳定、残留化肥污染环境等问题。加之,劳动力成本增加、种植效益下降,直接影响生产者的种粮积极性。因此,在保障小麦需求同时,如何实现籽粒品质、肥料利用率和经济效益的协同提升,形成绿色高效的小麦产业可持续发展局面,已成为亟需应对的现实问题。【前人研究进展】前人对小麦高产栽培机理开展了大量研究,但因生态条件、生产技术、品种类型等不尽一致。于振文等[1]就黄淮麦区小麦超高产栽培理论开展了大量研究,阐明旱茬小麦高产群体形成需要建立在合理群体结构的基础上,提出延长缓衰期、缩短速衰期、保持较长光合速率高值持续期,同步提高生物产量和经济系数的旱茬小麦高产途径。黄淮麦区高产旱茬小麦还表现出剑叶在展开前中期具有较高的光合速率,后期下降缓慢,光合时间长[2-3]。笔者所在研究小组针对稻茬小麦高产栽培机理,从群体质量、产量构成、光合和营养物质积累与分配、碳氮代谢特征等角度开展了系统研究,提出长江中下游稻茬小麦高产群体的实现关键在于获得适宜穗数的基础上,主攻每穗粒数与千粒重的协调增加,使群体在花前具有较高的光合面积和光合速率,花后光合面积衰减速率低,维持较高的光合面积,从而充分积累花后光合物质[4-8]。此外,卢百关等[9]研究了黄淮地区稻茬小麦高产群体特征;汤永禄等[10]和吴晓丽等[11]系统研究了四川盆地稻茬高产小麦产量结构和干物质积累特点、花后冠层结构、生理及同化物分配特性;李鸿伟等[12]阐明了稻茬高产小麦养分吸收与积累特征。小麦品种高效利用氮素机理和氮高效栽培机制亦有丰富报道。熊淑萍等[13]提出氮高效品种具有较高的根系生物量、根系活力、根系总吸收面积和根系活跃吸收面积,以促进其对氮素的吸收。李淑文等[14]认为氮高效品种具有高穗数和群体干物质积累量的特征。张洋等[15]亦指出氮高效品种具有较高的穗粒数和千粒重。丁永刚等[16]发现稻茬小麦氮吸收高效品种表现出生育前期较强的分蘖能力,生育后期旺盛的物质生产能力,尤其是花后有较多的光合物质向籽粒转运。丁锦峰等[17]和DING等[18]通过密肥措施构建氮效率差异的群体,分析认为稻茬小麦氮高效群体具有较高的花后和总的干物质积累量;发现氮素吸收效率决定氮素利用效率,提高氮素吸收量尤其是拔节至开花期氮素吸收能力是氮吸收高效的关键。密肥调控作为主要栽培技术,显著影响着小麦籽粒产量、品质和氮效率。DING等[19]提出稻茬小麦高产栽培的适宜施氮量为210 kg·hm-2,基肥、壮蘖肥、拔节肥和孕穗肥分施比例为 3∶1∶3∶3、孕穗肥追施时期为剑叶露尖;同时,研究表明配以密度 225 株/m2构成的密肥组合有助于提升小麦氮肥利用效率、蛋白质含量、湿面筋含量和出粉率[6]。LI等[20]研究认为增加种植密度是缩小高产与地产田间产量差的关键,并提出优化肥料管理能进一步促进产量提升、增加生产效率。DONG等[21]研究指示优化播种量和氮追施量能够通过促进氮素吸收和向籽粒转运,协同提升籽粒产量、蛋白质含量和面包品质。此外,MAHMMOD等[22]报道节氮生产条件下要实现丰产,减少1 kg·hm-2施氮量需要增加 0.6 kg·hm-2播种量,并提出江苏盱眙地区高产、氮高效的施氮量235 kg·hm-2、播种量 180 kg·hm-2的组合。【本研究切入点】前人对小麦高产或氮高效栽培技术和机理已有大量报道,但高产氮高效协同机制研究有待进一步深入。【拟解决的关键问题】本研究连续2年在江苏苏北(黄淮麦区南部)和苏中(长江中下游地区)稻茬麦区分别设置传统、高产、节肥和节肥增密4种栽培模式,研究不同模式间籽粒蛋白质含量、产量、氮效率、经济效益的差异,以期明确高产、高效模式及其产量构成、群体质量和氮素吸收转运特征,探明产量、氮效率与农艺生理性状间关系,揭示高产高效协同实现途径,为稻茬小麦优质、高产、高效协同栽培提供理论与技术支撑。
1 材料与方法
1.1 试验材料和地点
试验于 2017—2018和 2018—2019年分别在扬州大学江苏省作物遗传生理重点实验室试验田(邗江)、仪征市新集镇新集村试验田(仪征)和睢宁县睢宁现代农业示范区试验田(睢宁)进行。邗江与仪征点相距约30 km。邗江和仪征供试品种均为扬麦 25,睢宁为淮麦 33;前茬作物均为水稻。邗江试验田土壤为轻壤土,小麦播种前 0—20 cm土层土壤含有机质 21.0 g·kg-1、速效磷 97.5 mg·kg-1、速效钾 88.3 mg·kg-1和速效氮 76.1 mg·kg-1。仪征试验田土壤为淤泥土,小麦播种前 0—20 cm土层土壤含有机质 36.4 g·kg-1、速效磷 38.9 mg·kg-1、速效钾 87.0 mg·kg-1和速效氮 81.0 mg·kg-1。两年度,睢宁试验田土壤均为砂壤土,小麦播种前 0—20 cm土层土壤分别含有机质15.2和11.7 g·kg-1、速效磷44.3 和 43.2 mg·kg-1、速效钾 69.7 和 116.0 mg·kg-1、速效氮 66.1 和 76.0 mg·kg-1。
两年度试验点的气候条件见图 1。2018年度苗期干旱、越冬期温度较低、灌浆期降水多;2019年度苗期降水多、越冬期温度较高、生育中后期降水适量。

图1 试验点小麦生长季每月日均温和降水量Fig.1 Mean daily temperature and precipitation per month during wheat growth season in the experimental sites
1.2 试验设计
试验分别在江苏苏中地区的邗江(2018)、仪征(2019)和苏北地区的睢宁(2018、2019)设置4种不同栽培模式,采用单因素随机区组设计,3次重复。栽培模式具体措施详见表 1,其中传统模式(TCP)按江苏大面积生产中习惯的小麦栽培设定基本苗和施用氮、磷、钾肥;高产模式(HCP)按高产栽培设定,在TCP模式的基础上基本苗减少33.3%,氮肥施用量不变、氮肥后移、增施磷钾肥;减氮模式(RFCP)在HCP模式基础上减少氮、磷、钾肥施用量11.1%;减氮增密模式(IDCP)在RFCP模式基础上增加基本苗25%、减少氮、磷、钾肥 12.5%。同时,对应不同栽培模式设置4个不施氮处理对照。
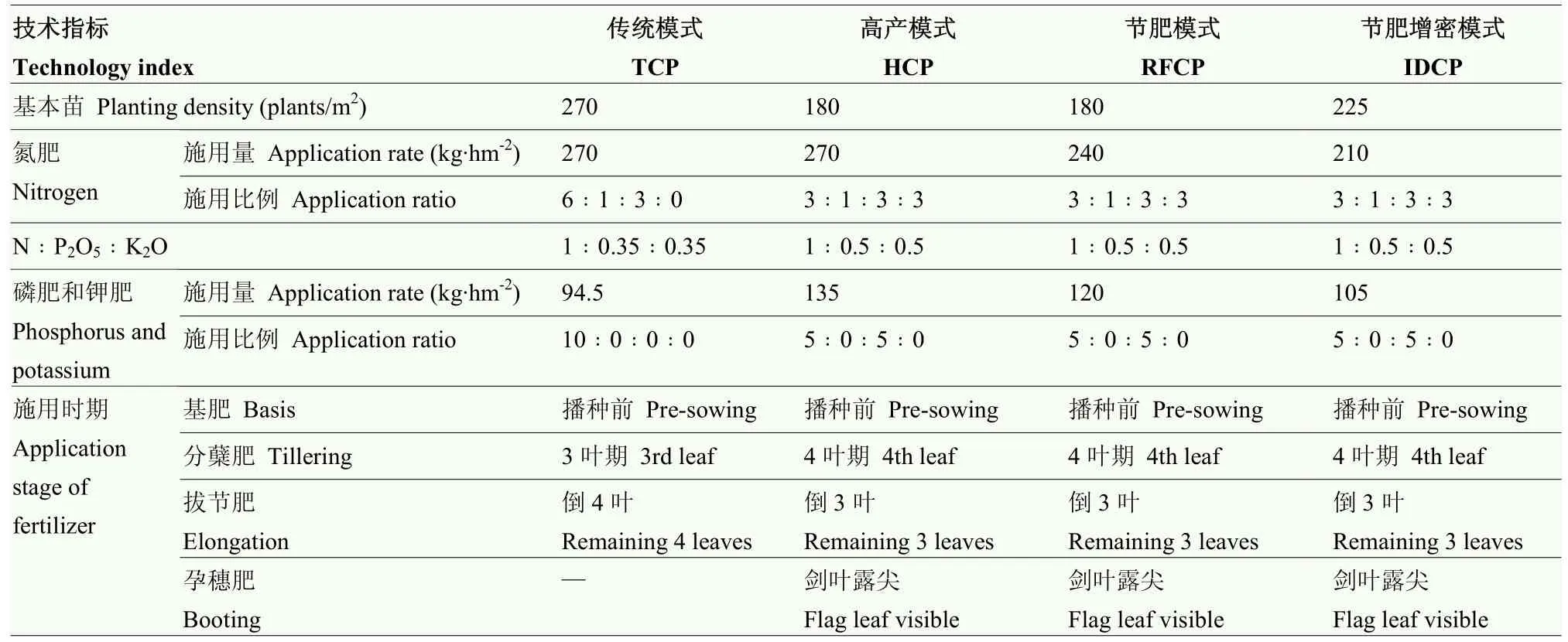
表1 不同栽培模式的基本苗和肥料运筹Table 1 Planting density and fertilizer management of different cultivation patterns
每个处理小区面积12.6 m2(3 m×4.2 m),人工条播,行距25 cm。邗江点于2017年11月2日播种,仪征点于2018年11月1日播种,睢宁点分别于2017年10月28日和2018年10月23日播种。2017年播种后因墒情不足进行了灌水以保证出苗;2018年苗期因极端降水排水沟有积水,在仪征较为严重。3叶期定苗,其余管理同当地大田栽培。
1.3 测定项目及方法
1.3.1 群体茎蘖数和叶面积指数(LAI) 于3叶期每个处理小区框定1 m2定苗区域,越冬始期和拔节期计数田间茎蘖数,成熟期计数最终穗数。最终穗数与拔节期茎蘖数之比即为茎蘖成穗率。于孕穗期、开花期和乳熟期分别在每个处理小区连续取30株小麦植株,用台式叶面积仪(LI-3100C,美国LI-COR公司)测定绿叶面积。LAI =样品叶面积(cm2)/株数×基本苗(株/m2)。
1.3.2 剑叶净光合速率(Pn) 于开花期和乳熟期,从各小区随机取长势均匀单茎的剑叶,采用便携式光合仪(LI-6400,美国LI-COR公司),设定人工光源900 μmol·m-2·s-1,于晴天 9:30—11:30 测定Pn,重复5次。
1.3.3 氮素积累、转运和利用 于越冬始期、拔节期、开花期和成熟期在每小区连续取样30株,分为茎鞘、叶片、穗轴(包括颖壳)和籽粒等不同器官,在 105℃下杀青60 min,然后在80℃下烘干至恒重,测定干重。样品粉碎后采用硫酸-靛酚蓝比色法测定含氮率,计算氮素积累量、转运量和氮效率。
植株氮素积累量=植株干重×含氮率;
氮素转运量=开花期营养器官氮素积累量-成熟期营养器官氮素积累量;
氮肥偏生产力(nitrogen partial factor productivity,NPFP)=施氮处理籽粒产量/施氮量;
氮肥利用效率(nitrogen use efficiency,NUE)=(施氮处理籽粒产量-不施氮处理籽粒产量)/施氮量;
氮素吸收效率(nitrogen uptake efficiency,NUpE)=(施氮处理氮素积累量-不施氮处理氮素积累量)/施氮量;
氮素生理利用效率(nitrogen utilization efficiency,NUtE)=(施氮处理籽粒产量-不施氮处理籽粒产量)/(施氮处理氮素积累量-不施氮处理氮素积累量)。
1.3.4 籽粒产量及其结构 乳熟期连续取 100个单穗,计数每穗粒数。成熟期从测定茎蘖数划定的1 m2区域人工收获后脱粒,自然晾干,测定籽粒含水率和重量,折算为13%含水率的籽粒产量。从脱粒籽粒中取样,数3个1 000粒测定重量和含水率,折算为13%含水率的千粒重。总结实粒数=最终穗数×每穗粒数;单穗产量=每穗粒数×千粒重/1 000。
1.3.5 粒叶比
粒叶比=单位面积籽粒干重/单位面积孕穗期绿叶面积。
1.3.6 籽粒蛋白质含量 取成熟期籽粒,采用硫酸-靛酚蓝比色法测定籽粒含氮量,含氮量×5.7即为籽粒蛋白质含量。
1.3.7 经济效益 经济效益按下式计算:总成本(元/hm2)=种子成本+肥料成本+作业成本+病虫草害防治成本;总收入(元/hm2)=籽粒产量×小麦单价;净效益(元/hm2)=总收入-总成本。为方便计算和比较,式中参数均为不变价,小麦收购价2.24元/kg,种子3.64元/kg,尿素2.25元/kg,磷酸二铵和氯化钾均为5.6元/kg,作业成本费用2 400元/hm2,病虫草防治费用420 元/hm2。
1.4 数据统计分析
采用 DPS 7.05进行相同年度和地点条件下不同栽培模式间指标的方差分析,以及不同生态条件和栽培模式下各指标间相关性分析。采用Excel 2016进行作图。
2 结果
2.1 栽培模式对籽粒蛋白质含量、籽粒产量和经济净效益的影响
栽培模式未显著影响籽粒蛋白质含量(表 2)。除2019年仪征点HCP模式下籽粒蛋白质含量显著高于 TCP和IDCP模式外,其他生态条件下不同模式间差异均不显著。不同栽培模式的籽粒蛋白质含量均达 12.5%以上,符合中筋小麦品质要求,其中 HCP和RFCP模式稳定达13%—14%,符合中强筋小麦品质要求。
籽粒产量和经济净效益仅在 2018年度受到栽培模式的显著影响,2019年度受影响不显著(表2)。2018年度,籽粒产量和经济净效益在邗江和睢宁点均以 IDCP模式下最高,相比 TCP分别提高 31.5%—33.5%和104.4%—239.1%;其次是HCP和RFCP模式,两者间差异均未达显著水平;以TCP显著最低。2019年度,籽粒产量以HCP模式最高,较TCP提高8.1%—13.2%;其次为RFCP和IDCP,较TCP提高1.0%—6.6%,差异不显著。因HCP和RFCP模式施肥次数多,增加了成本,经济净效益较TCP下降3.1%—7.7%;而IDCP施肥少,较TCP稳定或增加了经济净效益。

表2 栽培模式对籽粒蛋白质含量、籽粒产量和经济净效益的影响Table 2 Effects of cultivation patterns on grain protein content (GPC), grain yield, and economic benefit
综上所述,TCP模式相对低产、低效益;HCP模式可稳定高产,且蛋白质含量高;IDCP模式最具高产、高效益潜力,但蛋白质含量偏低。
2.2 栽培模式对氮肥利用率的影响
栽培模式显著影响不同生态条件下的氮肥偏生产力,表现为IDCP显著最高,TCP显著最低,HCP与RFCP间差异不显著(表 3)。不同栽培模式间氮肥利用效率、氮肥吸收效率和氮素生理利用效率表现因年度而异。2018年,上述3个氮肥利用率指标均表现为IDCP>RFCP、HCP>TCP,其中IDCP与TCP间差异显著,HCP与TCP间差异均不显著。2019年,氮肥利用效率表现为HCP、RFCP与IDCP间差异不显著,均显著高于TCP;氮素吸收效率以HCP和RFCP最高,其次为IDCP,3个模式间差异不显著,但TCP仅显著低于HCP和RFCP;氮素生理效率则以TCP和IDCP较高,仪征点不同模式间差异均不显著,而睢宁点TCP显著高于HCP和RFCP。总体而言,TCP模式氮肥利用效率最低;IDCP模式最易实现高氮肥利用效率。结果还表明,高氮肥利用效率的实现关键是较高氮素生理利用效率基础上提高氮素吸收效率。

表3 栽培模式对氮肥偏生产力、氮肥利用效率、氮素吸收效率和氮素生理利用效率的影响Table 3 Effects of cultivation patterns on nitrogen partial factor productivity (NPFP), nitrogen use efficiency (NUE), nitrogen uptake efficiency (NUpE), and nitrogen utilization efficiency (NUtE)
2.3 栽培模式对产量构成的影响
栽培模式未显著影响不同生态条件下的穗数和总结实粒数,但显著影响单穗产量(表4)。除2019年睢宁点TCP的穗数显著高于RFCP,以及2018年度邗江点HCP和IDCP的总结实粒数显著高于TCP,其他年度试验点不同模式间穗数和总结实粒数差异均不显著。栽培模式对单穗产量、每穗粒数和千粒重的影响效应因年度和地点而异。2018年,邗江和睢宁点单穗产量在HCP、RFCP与IDCP间差异不显著,均显著高于 TCP;每穗粒数表现为 HCP>RFCP、IDCP>TCP,HCP、IDCP均显著高于 TCP;千粒重表现为IDCP>RFCP>HCP>TCP,除邗江点IDCP与RFCP间差异不显著,其他模式间差异均显著。2019年,仪征和睢宁点单穗产量、每穗粒数和千粒重均表现HCP、RFCP>IDCP>TCP,其中HCP与RFCP间差异不显著,均显著高于TCP。结合籽粒产量表现进行分析,发现 HCP在获得高穗数基础上通过提高单穗产量尤其是每穗粒数实现高产;IDCP同样在获得高穗数基础上通过提高单穗产量实现高产,但依赖于高千粒重进行产量潜力提升。

表4 栽培模式对产量构成因素的影响Table 4 Effects of cultivation patterns on yield components
2.4 栽培模式对群体茎蘖数及其成穗率的影响
由表5可知,除2018年邗江点拔节期茎蘖数和2019年睢宁点茎蘖成穗率受栽培模式影响不显著外,两年度不同生态点条件下栽培模式均显著影响越冬始期和拔节期茎蘖数、茎蘖成穗率和冬前茎蘖数/最终穗数。越冬始期和拔节期茎蘖数在不同栽培模式间表现为 TCP>IDCP>HCP、RFCP,其中越冬始期茎蘖数在TCP下均显著高于其他模式,HCP与RFCP间差异不显著;拔节期茎蘖数在TCP下均显著高于HCP和RFCP,仅在 2018年睢宁和 2019年仪征显著高于IDCP,HCP与RFCP间差异不显著。茎蘖成穗率在不同栽培模式间均表现为TCP最低,在2018年显著低于其他模式,HCP、RFCP与IDCP间差异不显著;在2019年,TCP与IDCP间差异不显著,均低于RFCP,以HCP最高。结合最终穗数进行分析,表明不同栽培模式间实现高穗数的途径不一,TCP依赖于高茎蘖数,HCP主要通过高茎蘖成穗率,而IDCP通过协同高的茎蘖数和茎蘖成穗率。
冬前茎蘖数/最终穗数在不同模式间均以 TCP最高,2018年邗江和睢宁点以及2019年仪征点条件下显著高于其他模式,HCP、RFCP与IDCP间差异不显著;而2019年睢宁点条件下TCP与IDCP间差异不显著,显著高于HCP和RFCP。结合不同生态条件下群体的高产表现,分析得出高产群体的冬前茎蘖数/最终穗数存在适宜范围,在0.9—1.1。
2.5 栽培模式对孕穗期LAI和粒叶比的影响
孕穗期LAI可表示光合源水平,粒叶比可表示源库关系。栽培模式对孕穗期LAI和粒叶比的影响程度因年度和生态点而异(表5)。2018年邗江点和睢宁点,TCP的粒叶比均显著低于其他模式,HCP、RFCP与IDCP间差异不显著。孕穗期LAI在仪征点不同模式间表现为 IDCP显著高于 HCP,HCP显著高于 TCP和RFCP,但在睢宁不同模式间差异不显著。2019年两试验点不同模式间粒叶比差异均不显著;而孕穗期 LAI以HCP最高,其次为IDCP,TCP最低。结果表明HCP和IDCP模式可实现扩源基础上高水平协调源库关系。

表5 栽培模式对越冬始期和拔节期茎蘖数、茎蘖成穗率、冬前茎蘖数/最终穗数、孕穗期叶面积指数和粒叶比的影响Table 5 Effects of cultivation patterns on culm number at the stages of the beginning of wintering (BWCN) and stem elongation(SECN), culm fertility, BWCN/spike number, leaf area index (LAI) at booting, and the ratio of grain weight and leaf area(GLR)
2.6 栽培模式对绿叶光合特性的影响
由表6可知,栽培模式对单茎绿叶面积和剑叶净光合速率的影响程度因试验年度、地点和测定时期而有所差异。2018年度,邗江和睢宁点不同栽培模式间孕穗期和开花期单茎绿叶面积差异均不显著,乳熟期则表现为IDCP最高,仅在睢宁显著高于其他模式。剑叶净光合速率在开花期和乳熟期表现为 IDCP>HCP、RFCP>TCP,IDCP与TCP间差异显著,HCP与RFCP间差异均不显著。2019年度,仪征和睢宁点条件下孕穗期、开花期和乳熟期单茎绿叶面积表现为HCP>RFCP、IDCP>TCP,其中HCP与TCP间差异均达显著水平,HCP与RFCP间差异均不显著。开花期和乳熟期剑叶净光合速率在仪征点不同栽培模式间差异均不显著,在睢宁点表现为HCP最高,IDCP最低,两者间差异显著。上述结果表明,高产的模式(2018年IDCP和2019年HCP)可在维持高花后绿叶光合面积基础上协同提高绿叶净光合速率。

表6 栽培模式对单茎绿叶面积和剑叶净光合速率的影响Table 6 Effects of cultivation patterns on green leaf area per stem (GLA) and flag leaf net photosynthetic rate (Pn)
2.7 栽培模式对氮素积累和转运的影响
栽培模式对氮素积累和转运量的影响因年度、地点和时期而有所不同(表 7)。2018年度,邗江和睢宁点不同栽培模式间越冬始期氮素积累量表现为TCP显著最高,HCP、RFCP与IDCP间差异不显著;拔节期表现为TCP显著高于HCP和RFCP,与IDCP差异不显著;开花期和成熟期总体表现为 IDCP>HCP>RFCP>TCP,其中IDCP与HCP间差异均不显著;氮素转运量以IDCP显著最高,其次为HCP和RFCP,TCP最低,显著低于IDCP和HCP。2019年度,不同栽培模式间越冬始期氮素积累量表现 TCP、IDCP>RFCP、HCP,其中仪征点模式间差异均不显著,睢宁点TCP与IDCP间差异不显著,均显著高于其他两模式;拔节期表现为IDCP、TCP与HCP间差异不显著,以 RFCP最低;开花期和成熟期表现为HCP、RFCP>TCP、IDCP,HCP与RFCP差异不显著、显著高于TCP,TCP与IDCP间差异不显著;氮素转运量表现为 HCP>RFCP、IDCP>TCP,其中 HCP显著高于TCP,RFCP与IDCP间差异不显著。总体而言,TCP群体对于氮素在生育前期积累多、后期吸收弱、转运量低;HCP和RFCP群体在生育前期积累少、中后期吸收能力逐渐增强、转运量多;IDCP群体在生育中、前期吸收能力强、转运量足。

表7 栽培模式对氮素积累和转运量的影响Table 7 Effects of cultivation patterns on nitrogen (N) accumulation and transportation
2.8 茎蘖成穗率与籽粒产量、氮肥利用效率和农艺性状间关系
分析不同生态条件和栽培模式下籽粒产量与农艺性状间关系,发现茎蘖成穗率与籽粒产量、单穗产量、乳熟期单茎绿叶面积和剑叶净光合速率均呈显著线性正相关关系(图 2)。进一步分析茎蘖成穗率与氮肥利用效率、氮素吸收与转运间关系,显示茎蘖成穗率与氮肥利用效率、成熟期和开花期氮素积累量、氮素转运量呈显著线性正相关关系(图 3)。上述结果表明,提高群体茎蘖成穗率有助于维持灌浆期较高的单茎光合面积和光合速率,进而增加单穗和群体籽粒产量;促进了花前氮素吸收能力、增加了氮素转运量,提升了氮肥利用效率。

图2 不同生态条件和栽培模式下茎蘖成穗率与籽粒产量、单穗产量、乳熟期单茎绿叶面积和剑叶净光合速率的关系Fig.2 Relationship of culm fertility with grain yield, spike yield, green leaf area per stem (MRGLA), and flag leaf Pn at milk-ripening stage (MRPn) under different cultivation patterns and ecological conditions

图3 不同生态条件和栽培模式下茎蘖成穗率与氮肥利用效率、成熟期和开花期氮素积累量、氮素转运量间关系Fig.3 Relationship of culm fertility with nitrogen use efficiency (NUE), nitrogen accumulation at maturity (MANA) and anthesis stages (ANNA), and nitrogen transportation under different cultivation patterns and ecological conditions
3 讨论
3.1 不同栽培模式优质、高产、高效协同性及潜力的分析
本研究连续2个年度在2个生态区域进行试验,比较了4种栽培模式间籽粒蛋白质含量、籽粒产量、经济净效益、氮肥偏生产力和氮肥利用效率的差异,结果表明栽培模式对上述量、质、效指标的影响因生态条件存在差异。栽培模式间籽粒蛋白质含量差异较小,均达到中筋、中强筋小麦品质国家标准(GB/T 17320-2013),其中HCP和RFCP模式籽粒蛋白质含量较高,可稳定达 13%—14%。相比其他栽培模式,TCP模式籽粒产量、氮肥偏生产力、氮肥利用效率均最低,除2019年睢宁点籽粒产量外均显著最低;HCP模式籽粒产量次高或最高,氮肥偏生产力显著低于IDCP,氮肥利用效率在2018年低于IDCP、2019年度与IDCP差异较小;RFCP模式籽粒产量均低于HCP,氮肥偏生产力和氮肥利用效率与 HCP无显著差异;IDCP模式籽粒产量在2018年最高、2019年与最高产模式差异不显著,氮肥偏生产力显著最高,氮肥利用效率最高。就经济净效益表现而言,2018年度IDCP模式均最高,其次为HCP和RFCP,TCP最低,2019年度不同模式间差异均不显著。综上所述,通过综合措施构建的栽培模式可以实现高产、优质、高效协同提升目标,但协同性和提升潜力受生态条件制约。本研究构建的不同栽培模式中,TCP相对低产、低效益、低氮效率;HCP可稳定高产,且蛋白质含量高;IDCP最具高产、高效益、氮高效潜力,但蛋白质含量偏低。
3.2 高产、高效群体形成机理
小麦高产群体形成机理因生态条件、生产技术、品种类型等存在差异。对于稻茬小麦而言,实现9 000 kg·hm-2高产的关键在于适宜穗数基础上,协调增加每穗粒数和千粒重[5];形成适宜的穗数要求群体茎蘖数稳健发展,越冬始期茎蘖数达最终穗数的0.9—1.3倍,高峰苗1 250 株/m2,茎蘖成穗率约40%。本研究结果表明,相比TCP模式,HCP、IDCP模式均是在获得高穗数基础上提高单穗产量实现高产,这与前人研究结果基本一致。然而,HCP模式提高单穗产量更多地依赖每穗粒数,而IDCP则是千粒重。此外,不同栽培模式间实现高穗数的途径不一,HCP主要通过高茎蘖成穗率,而IDCP通过协同高的茎蘖数和茎蘖成穗率。这些结果与9 000 kg·hm-2高产群体产量结构和穗数形成要求不同,可能由于本研究模式实现的产量水平较低,穗数潜力还有较大空间。
高产群体需要源、库在高水平上实现协调[23]。粒叶比作为衡量群体源库协调水平的指标,其值高意味着群体质量高,表现为花后光合面积衰减慢、绿叶光合生产能力强且功能期长,有助于提升籽粒灌浆速率和粒重[24]。稻茬小麦9 000 kg·hm-2高产群体形成要求花前具有较高的光合面积和光合速率,花后光合面积衰减速率低,维持较高的光合面积,从而充分积累花后光合物质,在适宜库容基础上保障对库充实的需求[5]。本研究结果表明,HCP和IDCP模式可实现扩源(绿叶面积)基础上高水平协调源库关系(粒叶比)。此外,高产的模式(2018年IDCP和2019年HCP)可在维持高花后绿叶光合面积基础上协同提高绿叶净光合速率。这与前人研究结论基本一致,指示稻茬小麦高产的关键制约因素是花后光合源的不足。
提高氮肥利用效率可以通过氮肥吸收效率或氮肥生理利用效率或二者协同提高[13,25-26]。本研究表明,不同栽培模式下高氮肥利用效率的实现关键是较高氮素生理利用效率基础上提高氮素吸收效率。前人研究认为,小麦营养生长阶段是氮素吸收与积累的主要阶段,花前更多的积累氮素是决定氮肥利用效率的关键[27];但增加花后氮素积累可以延缓植株衰老,提高花后光合物质生产,亦可以提高氮肥吸收和利用效率[28]。本研究中不同栽培模式构建的群体在氮素积累特征上不尽相同,其中高产高效协同的IDCP群体在生育中、前期吸收能力强、转运量足;而高产的HCP群体生育前期积累氮素少、中后期吸收能力逐渐增强、转运量多。目前的栽培模式未有实现各生育阶段均高效吸收氮素,这可能给进一步提升高产、高效潜力提供了突破方向。
DING等[29]研究发现相同小麦品种种植于江苏北部相比种植于南部,能够提高茎蘖成穗率,有助于减缓花后光合面积的衰减,进而增强群体对生育后期自然资源的利用,协同提升穗数和单穗籽粒产量。本研究分析不同生态条件和栽培模式下茎蘖成穗率与籽粒产量、农艺性状间关系,也发现提高群体茎蘖成穗率有助于维持灌浆期较高的单茎光合面积和光合速率,进而增加单穗和群体籽粒产量。结果还表明,提升群体茎蘖成穗率还可促进花前氮素吸收能力、增加氮素转运量,提升氮肥利用效率。前人研究指出高茎蘖成穗率能够提升根系长度、表面积和体积等形态特征,进而促进氮吸收能力[30-31]。
3.3 高产、高效群体构建的技术途径
种植密度和肥料施用是小麦大面积生产中有效且实用的栽培技术。种植密度是群体形成的基础,其在适宜范围内能够使群体数量和最终穗数充足,群体质量和单茎生产力提升;而过高或过低均不利于后期群体光合生产[32-34]。适宜范围内较高的施氮量可促进分蘖发生,增加穗数和每穗粒数,提高粒重,进而提升单产和氮肥利用效率,但超过临界值,氮肥投入与产出比急剧降低[35-36]。较高的基施氮肥可促进小麦幼苗生长发育,而增加后期追肥施用量则促进中后期营养与生殖生长和产量构成各因素,但基肥与追肥应有一定比例,以实现充足穗数和较高单穗生产力的协同提升[37-38]。本研究不同栽培模式的基本苗、肥料施用量、时期和比例虽均有差异,但穗数差异较小,主要通过单穗产量提升增加最终产量。结果还表明,足穗的形成有3条途径:多茎蘖数但低成穗率(TCP)、低茎蘖数但高成穗率(HCP、RFCP)、协同高的茎蘖数和成穗率(IDCP)。此外,单穗生产力的提升主要通过维持高的花后绿叶光合面积基础上协同提高绿叶净光合速率,这就要求群体数量适宜且质量高以保证单茎健壮生长,加之适期、适量的追肥以延缓植株花后衰老。综合而言,IDCP模式以较低的氮肥施用量配以足够的基本苗,控制了群体数量、提升了单茎质量,加之中后期精准施肥,保障了营养需求、促进了单茎生产力,进而实现氮肥高效利用基础上的高产。
4 结论
通过基本苗、肥料施用量、时期和比例等栽培技术构建的不同栽培模式在实现小麦高产、氮肥高效的机制上有所差异。虽然高产的实现均是依赖于高穗数基础上高的单穗产量,但足穗的形成可分为多茎蘖数但低成穗率、低茎蘖数但高成穗率、协同高的茎蘖数和成穗率等3条途径。此外,相对高产的模式均可在维持高花后绿叶光合面积基础上协同提高绿叶净光合速率,实现扩源基础上高水平协调源库关系。高氮肥利用效率的实现关键是较高氮素生理利用效率基础上提高氮素吸收效率,但氮的吸收能力亦表现出生育前期吸收少、中后期吸收强、转运量多和生育中、前期吸收强、转运量足的差别。综合而言,稻茬小麦高产高效协同途径是获得充足穗数基础上,攻大穗,重点是增粒重;构建数量足、质量高的群体,越冬前群体茎蘖数满足预期穗数、重点提高茎蘖成穗率,花前高效吸收氮素、提升转运水平,花后维持较高单茎光合面积和强度、提升灌浆水平。为实现高产高效栽培,技术上应强调“适量增密、适度减肥、前氮后移、精准施肥”。
猜你喜欢
安徽农学通报(2024年12期)2024-07-13 20:26:34
江苏农业科学(2020年22期)2020-03-03 14:37:13
今日农业(2019年14期)2019-01-04 08:57:40
现代农业科技(2018年14期)2018-10-31 10:50:20
安徽农业科学(2017年28期)2017-05-30 14:55:41
水利规划与设计(2016年9期)2017-01-15 14:00:45
植物营养与肥料学报(2012年1期)2012-10-26 02:49:48
植物营养与肥料学报(2012年1期)2012-10-26 02:49:42
植物营养与肥料学报(2011年5期)2011-11-06 07:30:52
植物营养与肥料学报(2011年2期)2011-10-26 03:52:10
