
牛源犬新孢子虫NcGRA9基因原核表达及免疫原性分析
2023-03-08 05:01金卫东唐泽宇李作臣王海军贾立军
中国兽医学报 2023年2期
金卫东,唐泽宇,李作臣,王海军,陈 健,贾立军
(1.延边大学 东北寒区肉牛科技创新教育部工程研究中心,吉林 延吉 133002;2.延边特色产业发展中心,吉林 延吉 133002;3.延边州畜牧总站,吉林 延吉 133002;4.延边州动物疫病预防控制中心,吉林 延吉 133002)
犬新孢子虫(Neosporacaninum)是引起孕畜流产、死胎及运动神经系统障碍的胞内寄生性原虫,主要寄生于动物的脑、肝脏、脾脏等脏器组织以及生殖系统[1-2]。新孢子虫病(Neosporosis)呈世界性分布,我国北京、山东、吉林、新疆、青海等多个省、市(区)都有该病的报道[3]。犬新孢子虫对牛的危害最为严重,羊、马、驴、鹿、狐、鸽、水貂等多种动物均有该病感染的报道[4-5]。目前尚无用于预防新孢子虫病的商品化疫苗[6]。
致密颗粒蛋白(GRA)是新孢子虫致密颗粒细胞器分泌的蛋白,致密颗粒蛋白被释放到寄生液泡(pv)中,在维持寄主-寄生关系和获取营养物质方面发挥重要作用[7]。目前鉴定得到的新孢子虫GRA9蛋白与弓形虫GRA9蛋白同源性达到60%,且NcGRA9主要是亲水蛋白质,但也包含较短的疏水区。而NcGRA9蛋白作为新孢子虫疫苗的候选抗原之一,国内尚未有该蛋白研究的相关报道,本试验PCR扩增新孢子虫NcGRA9基因,构建NcGRA9重组表达载体,诱导表达重组蛋白并进行纯化,通过Western blot鉴定和小鼠免疫来验证NcGRA9蛋白的抗原性,以期为新孢子虫疫苗的研究奠定基础。
1 材料与方法
1.1 虫株和实验动物牛源犬新孢子虫、Vero细胞、pGEX-4T-1表达载体均由延边大学预防兽医实验室提供;BALB/c小鼠购自延边大学实验动物中心。
1.2 主要试剂BamHⅠ 酶、Not Ⅰ酶、GST纯化试剂盒、BCA试剂盒均购自生工生物工程(上海)股份有限公司;HRP标记的山羊抗小鼠IgG、蛋白质Marker均购自北京索莱宝科技有限公司;其他试剂均为国产分析纯。
1.3 引物的设计与合成根据ToxoDB网站发布的NcGRA9(NCLIV_066630)基因序列,应用Oligo 7.0软件设计特异性引物,引物序列P1:5′-TCTTATCGGGATCCATGCAGGGCGTGAC-3′,P2:5′-ATAAGAATGCGGCCGCTATTTCTCCGTT-ATGGTTCG-3′。
1.4 NcGRA9基因PCR扩增以提取的新孢子虫基因组DNA作为模板,进行PCR扩增。反应条件:94℃ 5 min;94℃ 45 s,56℃ 45 s,72℃ 45 s,35个循环;72℃ 7 min。PCR产物进行1%琼脂糖凝胶电泳。
1.5 NcGRA9基因重组克隆质粒的构建及测序将PCR扩增正确的条带回收,连接pMD18-T克隆载体,转化到DH5α中,提取重组质粒进行PCR和双酶切鉴定,并进行测序分析。
1.6 NcGRA9基因原核表达质粒的构建双酶切pGEX-4T-1载体,琼脂糖凝胶回收载体片段。将NcGRA9基因与pGEX-4T载体4℃过夜连接。
1.7 表达蛋白SDS-PAGE电泳和Western blot分析将pGEX-4T-NcGRA9重组质粒转化至BL21感受态细胞中IPTG诱导表达,GST标签蛋白纯化试剂盒纯化表达蛋白。SDS-PAGE电泳后,进行Western blot分析。
1.8 重组蛋白的小鼠免疫将纯化后的重组蛋白与弗氏佐剂(完全佐剂和不完全佐剂)混合。将30只BALB/c小鼠随机分为3组,每组10只,分为免疫组(重组蛋白+佐剂)、质粒组(pGEX-4T-NcGRA9)及对照组(PBS)。免疫组的重组蛋白含量200 μg/只,质粒组为等量pGEX-4T-NcGRA9,对照组为等体积PBS。每次接种间隔14 d,共接种3次,每次接种前及第3次接种后14 d采集血液,分离血清并保存待检。
1.9 免疫小鼠血清中抗体水平和细胞因子水平检测应用间接ELISA方法[8]检测免疫小鼠血清中IgG抗体水平,按照ELISA试剂盒说明书检测免疫小鼠血清中抗体亚类IgG1、IgG2a水平和细胞因子IFN-γ、IL-4水平。
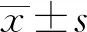
2 结果
2.1 牛源犬新孢子虫NcGRA9基因的克隆与测序分析以牛源犬新孢子虫的DNA为模板,经PCR扩增出大小为522 bp的条带,扩增条带与预期大小相符。构建pMD18-T-NcGRA9重组克隆质粒,经PCR和双酶切鉴定,PCR扩增出522 bp片段;经NotⅠ与BamHⅠ双酶切后,获得522 bp的目的条带与2 692 bp的载体片段。经测序表明,获得的NcGRA9基因序列与ToxoDB网站发布NcGRA9(NCLIV_066630)基因序列同源性为100%。
2.2 重组表达质粒pGEX-4T-NcGRA9鉴定pGEX-4T-NcGRA9进行PCR和双酶切鉴定,PCR扩增出522 bp目的片段;经NotⅠ、BamHⅠ双酶切后,获得522 bp的目的条带与4 969 bp的载体片段(图1)。
M.DL2000 DNA Marker;1.pGEX-4T-NcGRA9双酶切产物
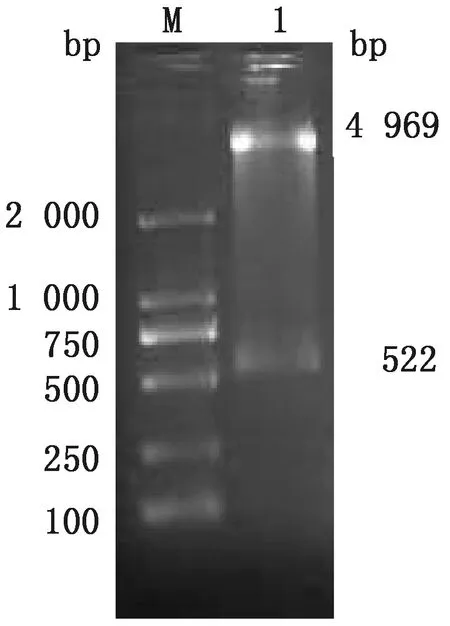
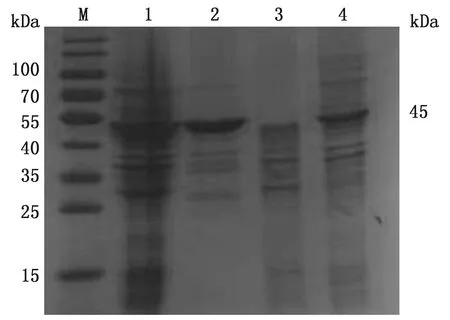
2.3 蛋白的表达与纯化取诱导3 h后的重组蛋白和纯化后的重组蛋白,进行SDS-PAGE电泳,在约45 kDa处出现目的条带,并以可溶性蛋白表达(图2)。Western blot分析表明,该蛋白能被小鼠抗新孢子虫阳性血清所识别,具有较好的反应原性(图3)。
M.蛋白质Marker;1.诱导后总蛋白;2.纯化后的蛋白;3.沉淀样本;4.上清样本
M.蛋白质Marker;1.NcGRA9重组蛋白;2.阴性对照
2.4 免疫小鼠血清中IgG、IgG1及IgG2a抗体水平检测结果ELISA检测免疫小鼠血清中IgG、IgG1、IgG2a抗体水平(图4),经统计学分析结果显示,免疫重组蛋白后,免疫组小鼠IgG抗体及其亚类IgG1、IgG2a水平均极显著高于质粒组与PBS对照组(P<0.01)。

**P<0.01表示差异极显著;*P<0.05表示差异显著;P>0.05表示差异不显著。下同
2.5 免疫小鼠血清中IFN-γ、IL-4细胞因子水平检测结果ELISA试剂盒检测免疫小鼠血清中细胞因子IFN-γ和IL-4水平(图5),经统计学分析,免疫组小鼠IFN-γ与IL-4水平均极显著高于质粒组与PBS对照组(P<0.01)。
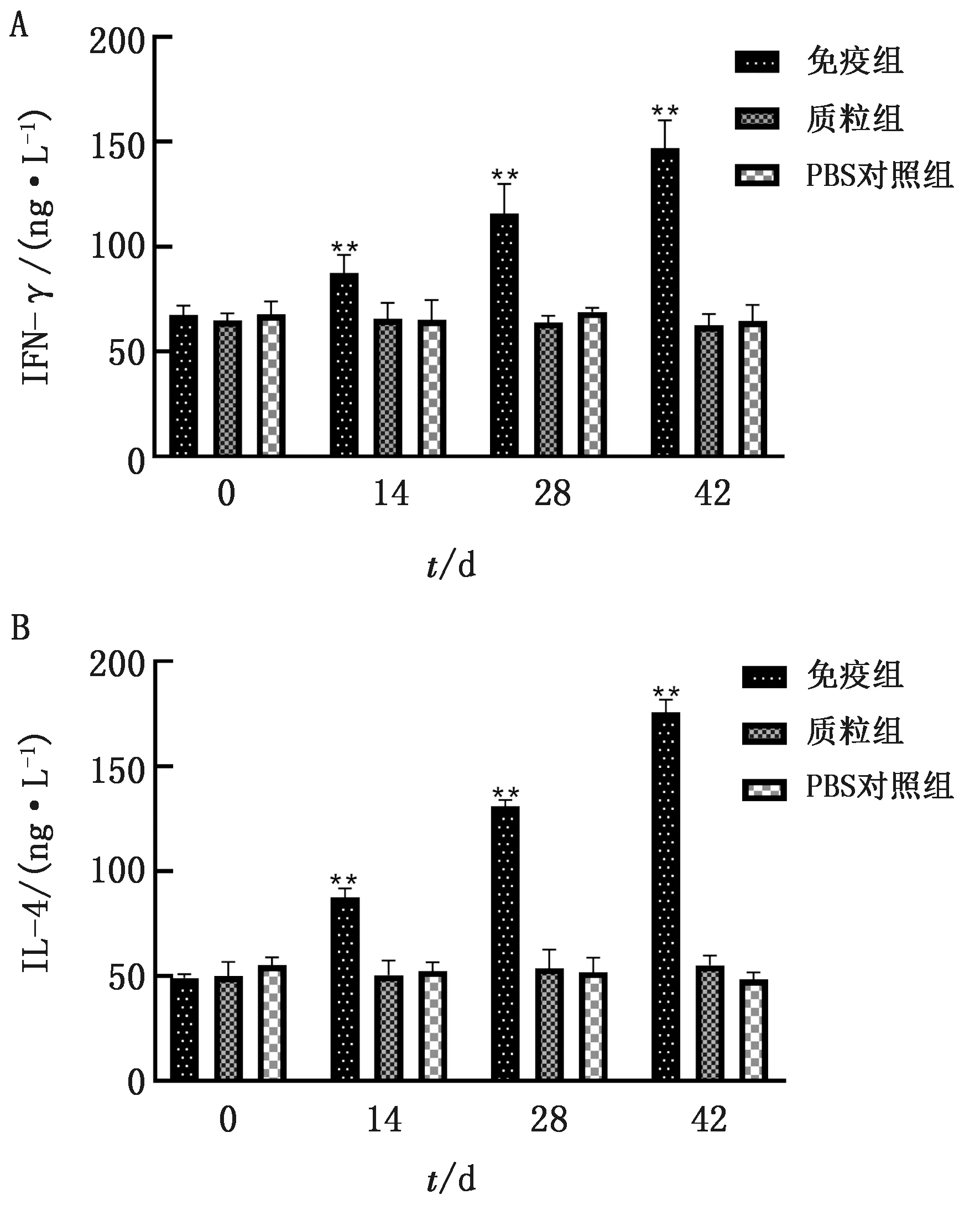
图5 不同免疫组小鼠血清IFN-γ(A)、IL-4(B)的水平检测
3 讨论
新孢子虫病严重影响我国的畜牧业发展,而目前尚无有效药物和疫苗防控新孢子虫病。国内外对NcGRA2、NcGRA6、NcGRA7、NcGRA16等均有不同程度的研究报道,而对NcGRA9的研究较少[9]。国内金春梅等[10]对NcGRA2进行了表达与鉴定;刑明等[11]对NcGRA6基因进行表达,并对其免疫效果进行初步分析;贾立军等[12]对NcGRA7进行了表达;杜博亚等[13]对NcGRA16进行了初步分析;但尚未见对NcGRA9的相关研究报道。
致密颗粒蛋白质结构和功能多样,在宿主细胞入侵后被释放到寄生液泡中,使虫体获得营养以满足生长增殖的需要[14]。近年来研究发现,致密颗粒蛋白在参与虫体入侵宿主细胞后以高水平分泌,在其后处于较低水平。NcGRA9作为可溶性蛋白存在于液泡中,在宿主细胞入侵中起重要作用,并且C末端截短的NcGRA9蛋白保留在这些细胞器中因为折叠不当,随后可能会退化。
以往的研究证实,GRA2、GRA6和GRA7的抗原分子是新孢子虫有效的候选疫苗蛋白基因[15]。而关于NcGRA9疫苗方面的研究甚少,仅有YU等[16]将新孢子虫致密颗粒蛋白NcGRA1、NcGRA4、NcGRA9、NcGRA14、NcGRA17及NcGRA23构建的pcNcGRA DNA疫苗接种BALB/c小鼠,这些真核表达质粒可引起强烈的Th1型免疫反应,诱导机体产生IgG2a抗体和IFN-γ细胞因子水平,免疫保护试验表明DNA疫苗均能够延长小鼠的存活时间和减少脑内虫荷量。由此可见,NcGRA9基因作为疫苗抗原的可行性。
为了解NcGRA9基因的表达形式及作为疫苗候选抗原的免疫原性,本试验对NcGRA9基因进行了原核表达,确定了NcGRA9基因表达重组蛋白为可溶性蛋白,Western blot分析具有良好的反应原性;接种BALB/c小鼠能够刺激机体产生细胞免疫应答与体液免疫应答,具有良好的免疫原性。本试验为NcGRA9基因蛋白作为新孢子虫病诊断的候选抗原和疫苗候选抗原奠定了基础。
猜你喜欢
山西地震(2019年1期)2019-03-20
食品科学(2018年10期)2018-05-23
渔业致富指南(2016年12期)2016-11-11
系统工程与电子技术(2016年2期)2016-04-16
科学大众(中学)(2015年9期)2015-10-12
西南医科大学学报(2015年1期)2015-08-22
少儿科学周刊·少年版(2015年3期)2015-07-07
中国光学(2015年1期)2015-06-06
中国当代医药(2015年9期)2015-03-01
西南军医(2015年6期)2015-01-23
