
猪瘟病毒E2蛋白猪源化单克隆抗体的制备及其中和活性鉴定
2023-03-08 05:01:52李淑红刘平黄孙慧敏仇华吉
中国兽医学报 2023年2期
李淑红,刘平黄,孙慧敏,仇华吉,李 素*
(1.中国农业科学院 哈尔滨兽医研究所 兽医生物技术国家重点实验室,黑龙江 哈尔滨150069;2.中国农业大学 动物医学院,北京 100193)
猪瘟(classical swine fever,CSF)是猪的一种急性、烈性、出血性传染病,给全球养猪业造成了巨大的经济损失,该病被世界动物卫生组织(world organization of animal health,OIE)列为必须报告的动物疫病[1-2]。CSF的病原是猪瘟病毒(classical swine fever virus,CSFV),为黄病毒科瘟病毒属成员。CSFV是单股正链RNA病毒,其基因组长约12.3 kb,只含1个大的开放阅读框(open reading frame,ORF),编码12个蛋白,其中衣壳蛋白C、囊膜糖蛋白Erns、E1、E2为病毒的结构蛋白,Npro、p7、NS2、NS3、NS4A、NS4B、NS5A和NS5B蛋白为病毒的非结构蛋白[3-4]。
CSFV的囊膜糖蛋白E2的相对分子质量为55 kDa,为Ⅰ型跨膜蛋白,其氨基端具有信号肽,羧基端具有跨膜区,将E2蛋白锚定在病毒囊膜上。E2可以通过二硫键形成同源二聚体,也可与E1蛋白形成E1-E2异源二聚体。研究表明,E1-E2异源二聚体对CSFV的侵入至关重要[5-7];此外,CSFV的E2蛋白在诱导保护性免疫反应中发挥重要作用,是产生中和抗体的主要糖蛋白[8-9]。
本团队前期制备了1株针对CSFV E2蛋白的单克隆抗体杂交瘤细胞株,命名为6E10。该杂交瘤细胞所分泌的单克隆抗体重链为IgG2a型,轻链为κ链[10]。由于杂交瘤细胞存在不稳定性,且不易保存,所以在本研究中,通过将6E10抗体的轻链(LC)、重链(HC)的基因扩增后与猪源抗体恒定区基因相融合,克隆至慢病毒表达载体,构建了稳定表达重组猪源化单克隆抗体6E10(p6E10)的HEK293S悬浮细胞系。应用AKTA蛋白纯化系统成功制备了纯化的p6E10抗体。通过Western blot、ELISA等试验证实,p6E10抗体组装成功,并与CSFV E2蛋白具有良好的反应性。中和试验结果表明,p6E10可以抑制CSFV的感染。因此,本研究制备针对CSFV E2蛋白的猪源化单克隆抗体p6E10可稳定、高效表达,并具有良好的反应性,为解析CSFV E2蛋白的结构和功能以及开发CSFV新型诊断制剂奠定基础。
1 材料与方法
1.1 细胞与病毒人胚肾细胞(HEK293T)、猪肾细胞(PK-15)和悬浮293细胞(HEK293S)由本实验室保存;CSFV石门株(CSFV-SM)由本实验室在PK-15细胞中增殖,于-80℃保存。
1.2 主要试剂及质粒DMEM培养基、胎牛血清、聚凝胺(polybrene)、HRP标记的山羊抗小鼠IgG、TMB底物、4′,6-DAPI购自Sigma-Aldrich公司;FITC标记的兔抗猪IgG、DH5α菌株购自Invitrogen公司;X-treme GENE HP转染试剂购自Roche公司;HiTrap Protein A HP 5 mL预装柱购自GE Healthcare公司;鼠源抗Strep单克隆抗体、HRP标记的兔抗猪IgG H&L购自Abcam公司;IRDye 800CW山羊抗鼠IgG购自LI-COR Bioscience公司;鼠抗猪IgG购自BD Pharmingen公司;293Pro®CD 293M无血清培养基购自上海源培生物科技股份有限公司;载体pFUGW、pSPAX2、pMD2.G、pCAGGS购自Addgene公司;PrimeSTAR®DNA聚合酶、T4DNA连接酶、QuickCut限制性内切酶购自TaKaRa公司;SDS-PAGE蛋白上样缓冲液购自Beyotime公司;猪源E2抗血清与6E10抗体轻、重链基因的重组质粒pMD18T-LC和pMD18T-HC由本团队保存。
1.3 慢病毒重组表达质粒的构建及鉴定以pMD18T-LC和pMD18T-HC质粒为模板,用表1中的引物扩增出p6E10-LC-Strep和p6E10-HC-Strep基因。将PCR产物以及pFUGW载体应用BamHⅠ和EcoRⅠ于37℃酶切2 h,经凝胶回收试剂盒回收目的基因与载体后,应用T4DNA连接酶于16℃连接12 h;连接产物转化至大肠杆菌感受态DH5α,涂布于氨苄青霉素抗性LB固体培养基,挑取单克隆菌落至氨苄青霉素抗性LB液体培养基中,振荡培养12 h后提取重组质粒,重组质粒应用BamHⅠ和EcoRⅠ进行双酶切鉴定,鉴定正确的重组质粒送吉林库美生物工程公司进行测序;将测序正确的质粒分别命名为pFU-p6E10-LC-Strep和pFU-p6E10-HC-Strep。
1.4 重组猪源化p6E10抗体的表达与纯化利用X-treme GENE HP转染试剂将pFU-p6E10-HC-Strep和pFU-p6E10-LC-Strep质粒分别与包装质粒pSPAX2、包膜质粒pMD2.G以21,14,7 μg共转染至生长于10 cm培养皿HEK293T细胞,48 h后收集上清,将上清经0.22 μm滤膜过滤后用超滤管浓缩10倍,获得慢病毒Lenti-p6E10-HC-Strep和Lenti-p6E10-LC-Strep,于-80℃保存备用。分别将2种浓缩后的慢病毒各1 mL与40 μL聚凝胺混合,加入HEK293S悬浮细胞,37℃、110 r/min悬浮培养3~5 d后,将该悬浮细胞扩大培养,收集细胞培养物在4℃、8 000×g离心15 min,收集上清经HiTrap Protein A HP 5 mL亲和层析柱纯化:先用Binding Buffer(20 mmol/L Na2HPO4,pH=7.0)洗涤层析柱,再将样品以5 mL/min的速度流过层析柱,用Binding Buffer洗去非特异性结合的蛋白,最后用Elution Buffer(0.1 mol/L Glycine-HCl,pH=2.7)洗脱抗体,收集洗脱液,加入1/10洗脱液体积的Neutralizing Buffer(1 mol/L Tris-HCl,pH=9.0),调节pH至7.4,于4℃透析12 h后,应用超滤管浓缩,获得纯化的p6E10抗体。应用SDS-PAGE与非还原性SDS-PAGE电泳后,进行考马斯亮蓝染色,以检测抗体组装情况,并通过Western blot试验以及双抗体夹心ELISA检测p6E10是否成功猪源化。

表1 p6E10-LC-Strep和p6E10-HC-Strep特异性引物序列
1.5 考马斯亮兰染色与Western blot鉴定纯化的p6E10抗体经SDS-PAGE和非还原性SDS-PAGE凝胶电泳后用考马斯亮兰染色15~30 min,进行脱色处理。Western blot用于确认纯化抗体是否成功猪源化,将p6E10抗体转移至NC膜后,5%脱脂乳常温封闭2 h,应用PBST洗涤3次,用HRP标记的兔抗猪IgG常温孵育1 h,PBST洗涤后,应用DAB显色。
1.6 双抗体夹心ELISA在ELISA板中加入100 μL/孔的1∶250稀释后的鼠抗猪IgG,终质量浓度为2 mg/L,4℃包被12 h。用PBST洗涤3次ELISA板,再加入100 μL/孔的5%脱脂乳,37℃封闭2 h。用PBST洗涤2次,加入100 μL/孔的p6E10抗体,37℃孵育1 h,弃掉抗体,用PBST洗涤3次。加入1∶4 000稀释的HRP标记的兔抗猪IgG 100 μL/孔,于37℃避光继续孵育1 h。用PBST洗涤3次后,加入TMB底物避光作用15 min显色,最后加入2 mol/L H2SO4终止反应,酶标仪检测D450 nm值。
1.7 间接ELISA及Western blot检测p6E10抗体与E2蛋白的反应性为检测p6E10抗体与E2蛋白的反应性,将CHO细胞表达的CSFV石门株E2蛋白作为包被抗原,包被质量浓度为0.04 mg/L,封闭后将纯化的p6E10抗体加入孔板中,37℃孵育1 h,PBST洗涤后加入1∶4 000稀释的HRP标记的兔抗猪IgG,37℃避光孵育1 h,TMB底物显色后用酶标仪检测D450 nm值。同时,将悬浮HEK293S细胞表达的CSFV-SM E2蛋白和C株E2蛋白应用SDS-PAGE电泳后转移至NC膜,5%脱脂乳常温封闭2 h,分别用p6E10和鼠源抗Strep单克隆抗体4℃孵育过夜,PBST洗涤3次,用HRP标记的兔抗猪IgG和IRDye 800CW山羊抗鼠IgG常温孵育1 h,用PBST洗涤后,应用DAB显色。
1.8 病毒中和试验检测p6E10抗体效价将纯化的p6E10抗体2倍梯度稀释,每个稀释度做4个重复,共做6个梯度。稀释好的抗体与每孔中100 TCID50的CSFV-SM等体积混合,37℃孵育2 h。孵育完成后加入预冷无水乙醇于-20℃固定30 min,然后加入1∶100稀释的CSFV阳性血清100 μL/孔,37℃孵育2 h。用PBST洗涤3次,加入1∶100稀释的FITC标记兔抗猪IgG,37℃避光孵育1.5 h。用PBST洗涤后,于倒置荧光显微镜下观察。
2 结果
2.1 重组表达质粒的构建及鉴定结果以质粒pMD18T-LC和pMD18T-HC为模板,应用特异性引物进行PCR扩增,扩增产物经1%琼脂糖凝胶电泳验证,可见大小约为750,1 500 bp的电泳条带,分别是p6E10-LC-Strep和p6E10-HC-Strep(图1A),与预期大小相符。将目的基因与pFUGW载体连接后转化大肠杆菌感受态DH5α,提取重组质粒后进行BamHⅠ和EcoRⅠ双酶切鉴定,在9 900,1 500,750 bp处有特异性条带(图1B),重组质粒经测序正确,表明pFU-p6E10-LC-Strep和pFU-p6E10-HC-Strep质粒构建成功。
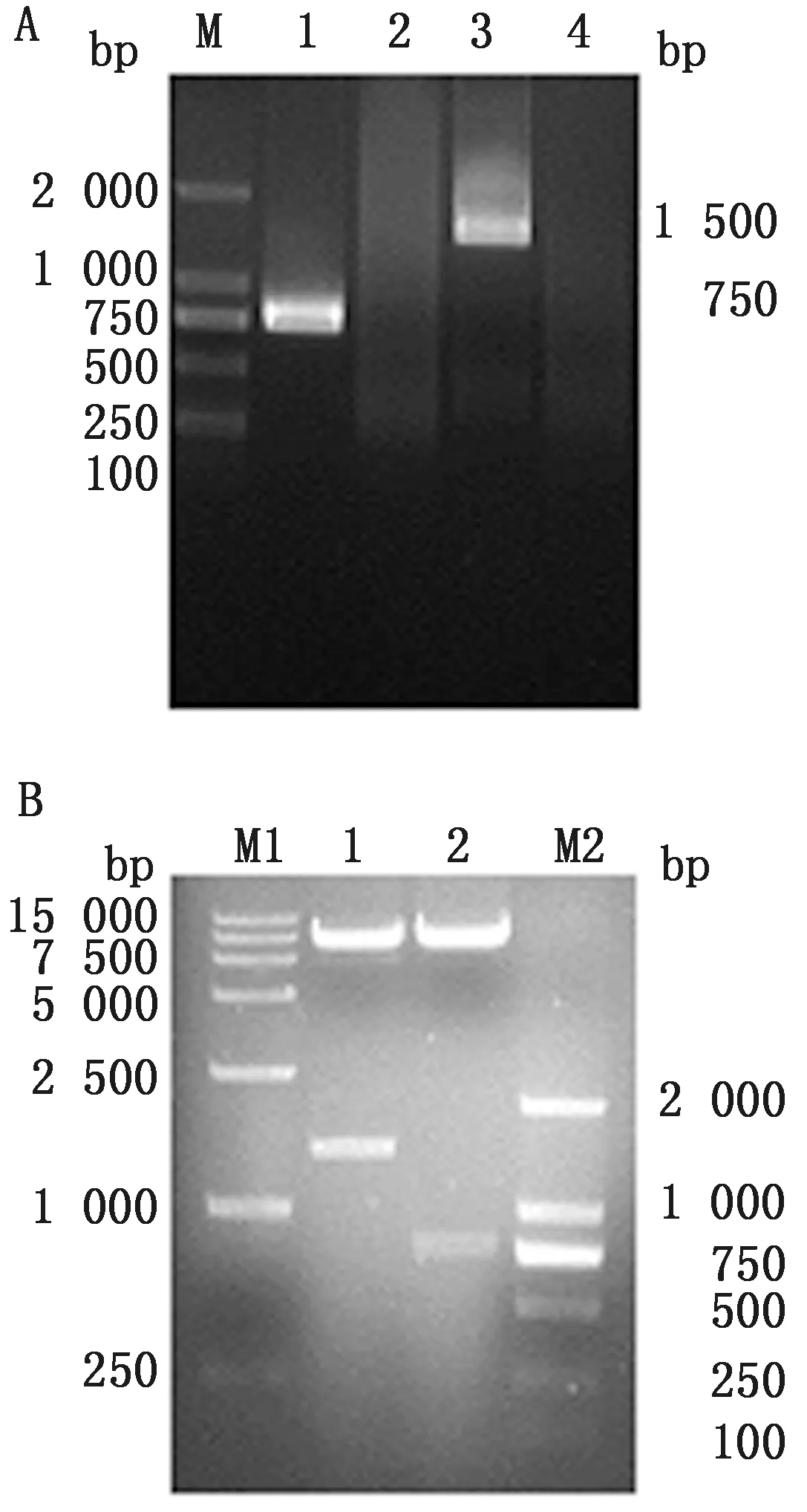
A.p6E10轻、重链PCR产物电泳结果(M.DL2000 DNA Marker;1.p6E10-LC-Strep PCR产物;2.p6E10-LC-Strep阴性对照;3.p6E10-HC-Strep PCR产物;4.p6E10-HC-Strep 阴性对照);B.重组质粒pFU-p6E10-HC-Strep 和 pFU-p6E10-LC-Strep 的双酶切鉴定(M1.DL15000 DNA Marker;M2.DL2000 DNA Marker;1.pFU-p6E10-HC-Strep 酶切产物;2.pFU-p6E10-LC-Strep酶切产物)
2.2 重组猪源化p6E10抗体的表达、纯化与鉴定结果HEK293S-p6E10细胞系培养上清应用AKTA蛋白纯化系统获得p6E10抗体(图2A),将纯化后的p6E10抗体进行SDS-PAGE电泳和考马斯亮蓝染色,以及Western blot鉴定。在SDS-PAGE中,纯化的p6E10抗体在30,60 kDa均有条带,表明该抗体纯化成功(图2B);而在非还原性SDS-PAGE中,纯化p6E10抗体在180 kDa处有条带(图2C),与预期重组猪源化抗体大小相符,表明含有轻链和重链的抗体成功组装。Western blot结果显示,重组猪源化p6E10抗体可被兔抗猪IgG所识别(图2D),同时双抗体夹心ELISA结果表明,p6E10可以与兔抗猪IgG反应(图2E),以上结果表明,该嵌合抗体已成功猪源化。
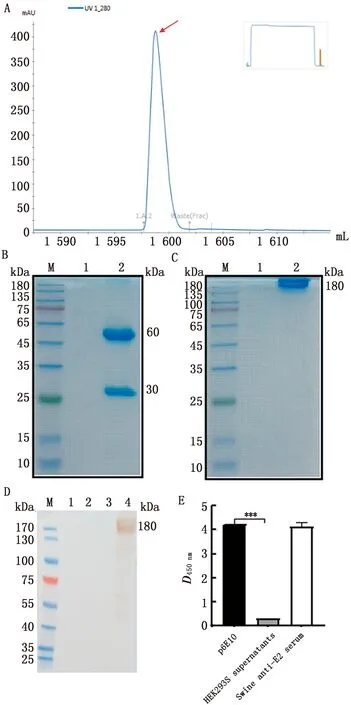
A.AKTA纯化p6E10图谱(红色箭头.p6E10收集峰);B.p6E10还原性SDS-PAGE(M.蛋白Marker;1.纯化的HEK293S细胞上清;2.制备的p6E10抗体);C.p6E10非还原性SDS-PAGE(M.蛋白Marker;1.纯化的 HEK293S细胞上清;2.制备的p6E10抗体);D.Western blot检测p6E10与 兔抗猪 IgG的反应性(M.蛋白Marker;1.未纯化的HEK293S细胞上清;2.未纯化的HEK293S-p6E10细胞上清;3.纯化的HEK293S细胞上清;4.制备的p6E10抗体);E.双抗体夹心ELISA检测 p6E10 与 兔抗猪 IgG 的反应性(p6E10.制备的p6E10抗体;HEK293S supernatants.纯化的HEK293S细胞上清;Swine anti-E2 serum.猪源E2抗血清)
2.3 重组猪源化p6E10抗体与E2的反应性鉴定应用间接ELISA检测p6E10抗体与CSFV E2蛋白的反应性,结果显示,与对照(纯化HEK293S细胞上清)相比,p6E10与CHO细胞表达的CSFV E2(CHO-E2)蛋白具有良好的反应性(图3A),说明该抗体可用于检测CSFV E2蛋白。同时,Western blot检测结果进一步证实,p6E10抗体与CSFV强毒石门株和弱毒疫苗C株的E2蛋白均有良好的反应性(图3B)。
2.4 重组猪源化p6E10抗体在CSFV感染中的抗病毒作用为了确定重组猪源化p6E10抗体对CSFV的抗病毒作用及其抗体效价,将CSFV-SM与2倍系列稀释的p6E10抗体预先孵育后感染PK-15细胞,48 h后进行间接免疫荧光试验(IFA)。结果显示,与对照相比,p6E10以1∶2,1∶4和1∶8的稀释度均可有效地中和100 TCID50的CSFV-SM,且在1∶16的p6E10稀释孔中,只检测到少量荧光(图4),结果表明,p6E10抗体对CSFV感染有一定的中和作用,且其抗体效价为1∶12。
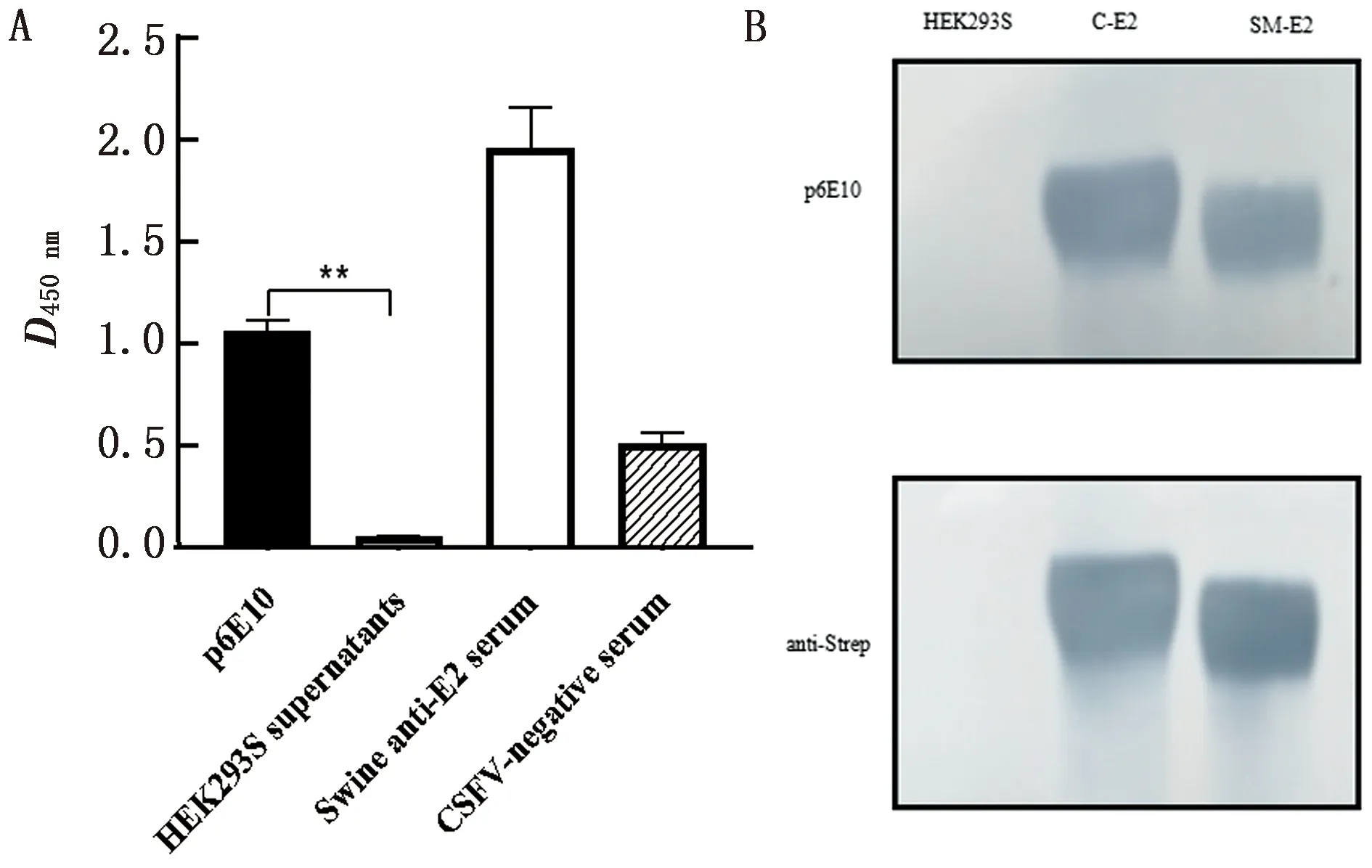
A.间接ELISA检测 p6E10 与 CSFV E2蛋白的反应性(p6E10.制备的p6E10抗体;HEK293S supernatants.纯化的HEK293S细胞上清;Swine anti-E2 serum.猪源E2抗血清;CSFV-negative serum.CSFV阴性血清);B.Western blot检测p6E10与 CSFV强毒和弱毒E2蛋白的反应性(HEK293S.HEK293S细胞上清;C-E2.C株E2蛋白;SM-E2.石门株E2蛋白)
3 讨论
CSFV的糖蛋白E2和Erns都是可诱导产生中和性抗体的病毒蛋白,而E2是最重要的免疫原性蛋白,因此应用E2蛋白作为免疫原制备识别CSFV的抗体。本实验室前期研究中制备了1株识别CSFV石门株E2蛋白的鼠源单克隆抗体6E10,其可以识别CSFV以及E2蛋白,但是传统制备单抗的方法是从小鼠腹腔抽取腹水,既费时又费力。此外,杂交瘤细胞的长期冷冻保存可能导致分泌抗体的杂交瘤细胞活力下降甚至杂交瘤细胞死亡。近年来,人源重组基因工程抗体已被临床应用于预防和治疗病毒感染[11-14]。然而,关于重组猪源抗体的报道较少,单克隆抗体多应用于病毒感染的宿主本身,因此将鼠源6E10抗体的轻、重链可变区基因与猪源IgG恒定区基因融合,表达出仍可特异性识别CSFV的重组猪源单克隆抗体p6E10。编码鼠源IgG恒定区的基因被猪源IgG替换,这可能导致重组抗体的糖基化、折叠和亲和力发生变化,更接近于猪体产生抗体的蛋白修饰情况。有趣的是,在Western blot结果中,重组猪源抗体p6E10反而可以更好地识别变性的E2蛋白,表明单克隆抗体猪源化后提高了其对病毒蛋白的敏感性,有助于CSF的诊断和预防。
目前,大多数基因工程抗体是通过质粒转染真核细胞而进行制备[15-16]。然而,在哺乳动物细胞中瞬时表达重组抗体费时、费力,而构建表达蛋白的悬浮细胞系是解决这一问题的有效途径。为了获得稳定表达高水平分泌型抗体的细胞系,本研究利用慢病毒系统构建了稳定表达p6E10抗体的HEK293S细胞系,并使用自动化高通量AKTA系统对抗体进行了纯化[17-18],且其产量(可达2 mg/L)和纯度均较高(98%以上)。本研究制备的猪源化单克隆抗体6E10可以用于CSFV E2蛋白的结构解析以及检测方法的建立。
本研究通过真核表达系统表达稳定的猪源重组单克隆抗体p6E10,证明该抗体成功组装及猪源化,与CSFV E2蛋白具有良好的反应性,并对CSFV感染具有抑制作用,为后续探究CSFV感染机制以及开发新型诊断试剂奠定了基础。
猜你喜欢
今日农业(2022年14期)2022-09-15 01:43:28
猪业科学(2021年3期)2021-05-21 02:05:54
中国现代医药杂志(2020年10期)2020-12-14 07:20:14
中成药(2017年9期)2017-12-19 13:34:27
CHINESE JOURNAL OF AERONAUTICS(2017年1期)2017-11-21 12:54:14
中成药(2017年5期)2017-06-13 13:01:12
现代检验医学杂志(2016年3期)2016-11-15 01:59:28
湖南畜牧兽医(2016年3期)2016-06-05 08:37:56
医学研究杂志(2015年3期)2015-06-10 06:41:52
特产研究(2015年1期)2015-04-12 06:36:20
