
GABA旁路在阪崎克罗诺杆菌耐干燥胁迫中的作用
2023-03-07 14:18黎丽曹怡芳张艳肖性龙
现代食品科技 2023年2期
黎丽,曹怡芳,张艳,肖性龙
(华南理工大学食品科学与工程学院,广东广州 510640)
阪崎克诺杆菌是一种杆状的革兰氏阴性能严重危 害特定人群生命安全的条件致病菌[1]。它可导致新生儿严重的脑膜炎、小肠结肠炎、败血症和菌血症,死亡率高达27%[2]。出生后两个月内的新生儿,尤其是体重不足的早产儿[3],胃酸较少,免疫能力不足,他们的细菌感染风险更高[4],病死率可达到40%~80%[5]。相对于其它常见食源性致病菌大肠杆菌O157:H7、肠炎沙门氏菌、鼠伤寒沙门氏菌和单核细胞增生李斯特菌,阪崎克罗诺杆菌具有更强耐干燥胁迫能力[6]。在高温干燥的生产环境中,阪崎克罗诺杆菌在PIF基质成分的保护作用下,依旧能表现出较强的存活能力[7],PIF在生产过程中通常采用巴氏杀菌,PIF生产设施中与克罗诺杆菌污染较高相关的区域包括喷雾干燥器、流化床和空气过滤设施[8],巴氏灭菌只能在PIF生产的早期阶段灭活阪崎克罗诺杆菌,但在控制来自下游成分、过滤器、喷雾干燥塔和其他加工设备的污染方面无效,阪崎克罗诺杆菌能在喷雾干燥中存活并长期存在[9]。由此可能形成巨大的食品安全隐患,成为食品微生物污染与控制的难点。综上所述,研究阪崎克罗诺杆菌耐干燥,最重要的一个原因是其污染的传播途径是水分活度低级的食品-PIF,因此研究阪崎克罗诺杆菌耐干燥特性有重要意义。
γ-氨基丁酸(γ-Aminobutyric Acid,GABA)是一种广泛存在于微生物和动物、植物中的非蛋白氨基酸。γ-氨基丁酸转氨酶基因gabT是阪崎克罗诺杆菌代谢GABA分支的关键基因之一。在植物方面,在高等植物中,很多研究报导表明GABA旁路是植物耐受干旱胁迫的重要机制,逆境胁迫会导致植物体内大量积累GABA[10]。Bao等[11]通过沉默番茄中gabT基因,导致番茄中GABA含量增加,琥珀酸含量降低,提高番茄耐盐性。Renault等[12]研究表明,gabT是耐盐拟南芥GABA代谢调控的关键。GABA代谢旁路在微生物对环境胁迫如耐酸[13]、应激反应[14]和致病性细菌毒力[15]的响应中起着重要作用。研究表明,突变的gabT基因降低了丁香假单胞菌[16]的致病性。Hu等[17]通过使用iTRAQ等蛋白组学方法系统性研究了阪崎克罗诺杆菌ATCC 29544的耐干燥胁迫机制,发现在干燥胁迫下腐胺(Putrescine)代谢相关的酶PuuR、PuuA、PuuD大幅上调表达,表明菌体内GABA的合成被大幅激活,因而显示了腐胺等多胺降解积累的GABA可能是阪崎克罗诺杆菌耐干燥胁迫的重要机制,合成路径如图1所示。因此,GABA对生物体耐受逆境胁迫有重要作用,然而相关研究多见于植物,但在细菌方面的研究很少。gabT基因是否能通过调节GABA代谢影响阪崎克罗诺杆菌的活性,干燥应激环境对GABA合成量有什么影响尚不清楚。
鉴于阪崎克罗诺杆菌较其他肠杆菌具有较强的干燥环境耐受性,能够长期生存于PIF中,且对新生儿有潜在的危害性,因此对于阪崎克罗诺在干燥环境耐受机制的研究有重大意义。为了探究gabT基因是否能通过调节GABA代谢影响阪崎克罗诺杆菌的活性,干燥应激环境对GABA合成量的影响,本研究通过基因敲除技术获得阪崎克罗诺杆菌ATCC 29544的gabT基因缺失株即ΔgabT菌株,以阪崎克罗诺杆菌野生株WT 和ΔgabT菌株为研究对象,开展干燥逆境胁迫下菌体存活率测定、细胞内GABA含量测定实验,同时借助扫描电子显微镜分析干燥胁迫条件下野生株WT和ΔgabT菌株的形态变化,从而验证GABA代谢旁路在阪崎克罗诺杆菌耐干燥胁迫响应中的重要作用。为阪崎克罗诺杆菌中基于GABA旁路的靶向调控奠定了基础,从而为阪崎克罗诺杆菌防控提供新思路。
1 材料与方法
1.1 原料
1.1.1 原料与试剂
惠氏婴幼儿配方奶粉,上海惠氏;无水乙醇(分析纯)、2.5%戊二醛固定液(电镜专用)均购于上海艾妍生物科技有限公司;GABA试剂盒(项目编号69-99842)购于Merck Biotech公司。
1.1.2 培养基及菌种
胰酪大豆胨琼脂培养基(TSA)、胰酪大豆胨液体培养基(TSB)购于广东环凯微生物科技有限公司;阪崎克罗诺杆菌ATCC 29544购于中国工业微生物菌种保藏管理中心;ΔgabT突变株在实验中获得;菌种均在37 ℃,180 r/min摇床TSB中培养活化24 h,调整菌悬液浓度为6 lg CFU/mL后备用。
1.1.3 主要仪器设备
XW-80A微型漩涡混合仪,上海泸西分析仪器厂有限公司;UV-7500紫外可见分光光度计,上海棱光技术有限公司;LRH-250A-2恒温培养箱,韶关市泰宏医疗器械有限公司;LEO1530VP扫描电子显微镜,德国Zeiss公司;HZQ-F100恒温振荡培养箱,哈尔滨市东联仪器有限公司;5424小型离心机,德国Eppendorf公司;YXQ-LS-100Ⅱ立式压力蒸汽灭菌器,郑州南北仪器设备有限公司。
1.2 实验方法
1.2.1 基因敲除突变体的构建
利用Luo等[18]的方法构建了阪崎克罗诺杆菌ATCC 29544的gabT敲除突变体,敲除位点如图1所示。使用表1中的引物。将gabT基因上下游DNA片段进行PCR融合,克隆到自杀载体质粒pLP12cm中。将重组自杀载体从β2163转移到阪崎克罗诺杆菌ATCC 29544进行同源重组。用0.4% L-阿拉伯糖筛选gabT基因突变体。用相同的引物对进行测序,通过PCR确认缺失突变体。在含有20 μg/mL氯霉素(Chloramphenicol,Cm)和0.3% D-葡萄糖的LB平板上,选择整合质粒到特定染色体位点的阪崎克罗诺杆菌单交叉细胞。在添加0.4% L-阿拉伯糖的反选择LB平板上选择双交叉重组突变体。缺失突变体检测采用两种外部引物分别锚定gabT基因的上下游区域。

图1 敲除基因位置示意图Fig.1 Schematic diagram of knockout gene location

表1 基因敲除的引物设计Table 1 Primer design
1.2.2 干燥胁迫下WT和ΔgabT突变菌株存活率测定
将106CFU/mL的WT和ΔgabT突变株菌悬液重悬于复配婴幼儿乳粉中,惠氏婴幼儿配方奶粉(上海惠氏)已按照GB 4789.40-2016所述检测,证明无阪崎克罗诺杆菌污染。在无菌培养皿中制备0.5 mL点重悬的细菌培养物。将无盖培养皿置于含脱水硅胶的40 ℃培养箱中培养4 h,干燥后盖上盖,在干燥皿中25 ℃保存。每隔一周用平板计数法测定存活细胞数量。本研究以 37 ℃ TSB培养12 h的阪崎克罗诺杆菌为对照样品。
式中:
A——存活率,%;
n1——实验组菌落数;
n0——对照组菌落数。
1.2.3 干燥胁迫下WT和ΔgabT菌株GABA含量的测定
GABA含量按照GABA试剂盒操作说明进行。试剂盒购于Merck Biotech公司(项目编号69-99842)。本试剂盒采用酶联免疫吸附法测定标本中GABA含量。GABA抗体、GABA与辣根过氧化物酶(HRP) 标记的GABA抗体形成抗体-抗原-酶标记的抗体复合物,经彻底洗涤后与底物四甲基联苯胺(TMB)着色。TMB经辣根过氧化物酶(HRP)催化,在酸的作用下转化为蓝色和最终的黄色。颜色的深度与GABA呈正相关。用酶标仪检测培养物的OD450根据标准曲线计算样品中GABA的浓度。
1.2.4 干燥胁迫对WT和ΔgabT菌株细胞形态的影响
参照Paula等[19]的方法,使用扫描电子显微镜观察干燥胁迫对WT和ΔgabT突变菌株细胞膜形态的影响。使用生理盐水溶出方法1.2.2中干燥后的盖玻片上的细胞,加入2.5%的戊二醛溶液中固定过夜(4 ℃)。后用不同浓度梯度(10%、30%、50%、70%、90%、100%)的乙醇溶液依次洗脱菌体,每次10 min。真空干燥后,将盖玻片粘贴在扫描电镜专用载物台上,待样品脱水镀金后,在Zeiss扫描电子显微镜下观察细胞微观形态结构。
1.3 数据处理
使用和Origin 2021软件对实验结果进行统计分析并绘图,使用SPSS 22.0软件进行差异显著性分析,P<0.05为显著差异。
2 结果与讨论
2.1 干燥胁迫对WT菌株和ΔgabT菌株存活率的影响
实验前已证明,ΔgabT菌株在适宜环境中能正常生长。通过平板计数法测定WT和ΔgabT菌株干燥后的存活率,如图2所示,WT与ΔgabT菌株在干燥条件处理一周后的存活率均显著下降。干燥一周后野生株存活率为1.57%,ΔgabT菌株存活率为28.64% (P<0.05),可以看出,在干燥条件胁迫后,ΔgabT菌株存活率更高。随着干燥时间的增加,在干燥第二周后,WT菌株存活率下降至0.18%,ΔgabT菌株存活率下降至20.66%(P<0.05),由此可见,随着干燥时间的增加,ΔgabT菌株存活率情况更显优势,且证明在干燥环境中,阪崎克罗诺杆菌具有一定的存活能力并且能够长期存活,这与易萌等[20]研究结果一致。在此之后,菌株的存活情况维持在较稳定的状态,干燥五周后WT和ΔgabT菌株存活率分别为0.15%和19.34%(P<0.05)。说明gabT基因的缺失使得阪崎克罗诺杆菌在干燥胁迫条件下更易存活,且存活率相比野生型菌株大幅提升,表明gabT基因的缺失可能在一定程度上有助于细胞抵御干燥环境。

图2 干燥处理后细菌存活率的变化Fig.2 Changes of bacterial survival rate after drying treatment
2.2 干燥胁迫对WT和ΔgabT菌株中GABA含量的影响
ΔgabT菌株和WT菌株中GABA含量结果如图3所示,干燥两周后,ΔgabT菌株GABA含量由 6.54 μmol/L增加到8.54 μmol/L(P<0.05),WT菌株GABA含量由4.41 μmol/L增加到5.56 μmol/L (P<0.05),在应对干燥条件胁迫时,ΔgabT菌株和WT菌株中GABA含量都有所增加。干燥被认为是一种高渗透的胁迫状态,对于阪崎克罗诺杆菌在干燥环境中的调控机制由两级相应完成,第一级主要的反应是积累电解质,如钾,谷氨酸等,由转运蛋白进入细胞内,通过提高细胞内离子浓度使渗透压升高来抵消升高的外部渗透压[21]。但长时间的维持较高的离子浓度可能会对细胞造成高浓度离子毒害作用,因此第二级反应是基于吸收或合成其他种类具对细胞损伤较小且能长时间存在于细胞内的相容性物质,随着相容性物质的累积,导致细胞内渗透压升高,以此来抵御外界环境对细胞的损伤[22]。GABA是一种四碳非蛋白氨基酸,在正常生理pH条件下为两性离子,易溶于水。它的生化特性类似于小的渗透分子,如脯氨酸和甜菜碱。因此,GABA可以作为一种小分子渗透调节物质,增加细胞质中的渗透水势,提高细胞的保水性能,减缓干燥对细胞的损害。Shabarinath等[23]的相关研究表明,干燥环境下海藻糖在阪崎克罗诺杆菌中积累。且强调了其他次生渗透物质如脯氨酸、胆碱等物质在干燥生存中的潜在辅助作用。在干燥环境下,ΔgabT菌株和WT菌株都可能通过累积GABA来抵御干燥胁迫。
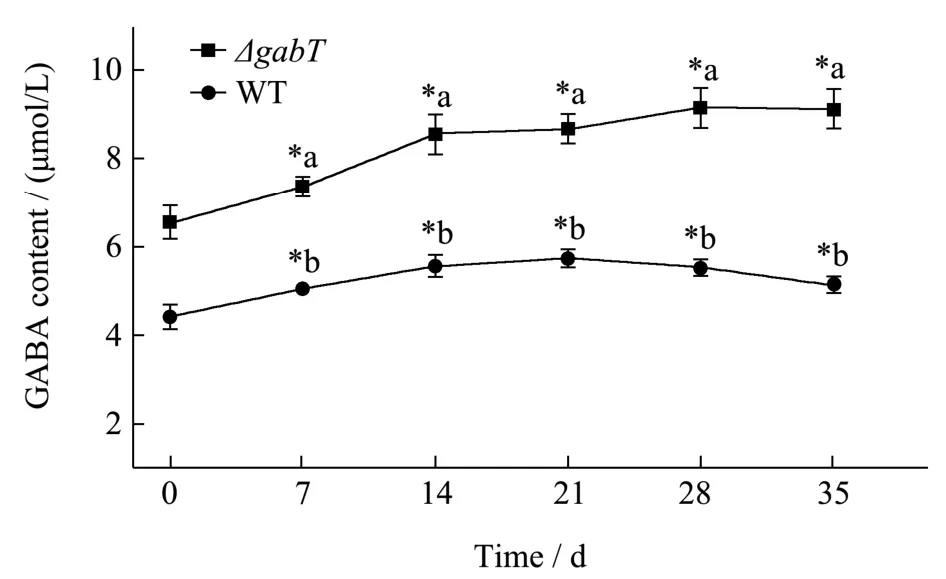
图3 干燥处理后细菌GABA含量的变化Fig.3 Changes of GABA content of bacteria after drying
干燥五周后,ΔgabT菌株GABA含量达到 9.12 μmol/L,WT菌株在干燥2~5周期间GABA含量保持相对稳定,在干燥5周后GABA含量为5.13 μmol/L(P<0.05),由此可见,在干燥胁迫下ΔgabT菌株GABA含量显著高于WT菌株,ΔgabT菌株由于gabT基因的缺失,其对应的GABA转氨酶功能缺失使得其具有更强的GABA累积能力。Du等[22]的研究也表明阪崎克罗诺杆菌的干燥阻力的调节机制与钾积累和流出以及相容溶质的吸收和合成有关,与相容性物质合成与转运相关的betAB、proVW等基因在耐干燥性强的菌株中的水平高于弱干燥耐受性菌株中的基因水平,这些基因的较高水平可能使菌株对干燥环境更具抵抗力。在本研究中,ΔgabT菌株在干燥胁迫下积累了更高的GABA(图3),这一结果与推测是吻合的。在ΔgabT中,由于gabT基因的缺失,GABA代谢受阻,导致GABA积累。GABA的积累有助于细胞抵抗干燥胁迫,ΔgabT菌株较于WT菌株更高水平的GABA累积导致ΔgabT的存活率高于WT。
2.3 干燥胁迫对WT和ΔgabT菌株细胞形态的影响
通过扫描电子显微镜观察了干燥一周后野生型菌株和ΔgabT突变株菌体形态的变化,结果如图4。从图4中可以看出,未经处理的WT菌体无褶皱,菌体表面光滑(图4a),菌体形态轮廓正常;未经处理的ΔgabT菌体呈现出轻微皱缩情况(图4c),经干燥条件处理后,野生型菌株与ΔgabT突变株菌体均出现皱缩现象(图4b和4d),表面轮廓不完整现象。相比与ΔgabT菌株而言,WT菌体部分出现皱缩、粘连、破裂现象,菌体变形更加严重(图4b)。上述结果表明,干燥环境对于阪崎克罗诺杆菌WT菌株和ΔgabT菌株细胞形态会造成一定的破坏和损伤,但菌体依然具有存活能力,依旧具有一定的危害性。

图4 干燥处理后细菌细胞形态的变化Fig.4 Changes of bacterial cell microstructure after drying
由细胞形态变化可以发现,干燥对于ΔgabT突变株菌体形态影响更小,由此推测出gabT基因的缺失使得细胞内GABA代谢受阻进一步累积,维持细胞内渗透压在一定程度上保护细胞膜的完整性。微生物细胞膜的完整性是保证其维持自身新陈代谢等正常生理活动的重要因素[24]。胡心怡等[25]研究发现百里香酚与肉桂醛通过破坏沙门氏菌细胞膜的完整,从而抑制了沙门氏菌的增殖。失水干燥引起细菌的细胞质的渗透性失衡,膨胀压力的损失导致水在细胞膜上的位移,并导致细胞收缩[26]。ΔgabT菌株中GABA的大量累积,在一定程度上保护了细胞失水收缩,相对于WT菌株而言使菌体的细胞膜不易造成破坏和损伤,菌体形态不易发生改变。细胞膜是细菌与环境间的半透膜屏障,当其结构和完整性被破坏,将会导致细胞失去正常的生理代谢活动,进而使菌体失活和死亡[27,28],该过程往往伴随着细胞膜的通透性的变化以及胞内成分的流失[29]。因此,在干燥条件下GABA的累积在有助于维持阪崎克罗诺杆菌的胞内渗透压,维持菌体形态的基础上一定程度保证细胞正常生理代谢活动,进而防止其因细胞膜通透性变化导致的内容物泄露以及细胞收缩导致菌体的死亡。
3 结论
本文通过基因敲除手段,构建ΔgabT菌株,同时对WT菌株和ΔgabT菌株耐干燥能力及GABA含量测定。干燥条件下ΔgabT菌株存活率明显高于WT菌株,且ΔgabT菌株干燥中GABA含量也远高于WT菌株,说明GABA的积累与gabT基因缺失有关,且GABA累积在耐干燥胁迫中具有关键作用,在干燥条件下通过调控GABA的代谢,保护阪崎克罗诺杆菌抵抗干燥胁迫,结合扫描电子显微镜检测进一步证实了GABA累积有助于阪崎克罗诺杆菌抵御干燥胁迫。故经基因敲除手段获得的ΔgabT菌株由于GABA代谢受阻,GABA累积使得干燥后阪崎克罗诺杆菌更耐干燥。总结得出gabT基因在干燥胁迫下的功能特性以及GABA含量与菌体耐干燥胁迫关系,在已有传统物理化学致病菌防控手段基础上为探究阪崎克罗诺杆菌逆境胁迫机制提供新的思路,为预防和控制食品(特别是婴幼儿配方奶粉)中阪崎克罗诺杆菌污染奠定基础。
猜你喜欢
当代水产(2022年1期)2022-04-26
中国乳品工业(2021年11期)2021-03-31
食品科学(2020年17期)2020-09-21
中国酿造(2020年4期)2020-05-15
食品工业科技(2020年5期)2020-02-15
食品与机械(2019年10期)2019-11-16
食品科学(2019年18期)2019-10-08
中国调味品(2017年2期)2017-03-20
现代检验医学杂志(2016年3期)2016-11-15
中国乳品工业(2015年7期)2015-12-16
