
过硫酸钾强化蔬菜废弃物好氧堆肥的通风速率优化及其微生物特征
2023-02-26 12:39梅娟段恩帅苏良湖李贲周筱洁
江苏农业科学 2023年24期
关键词:微生物
梅娟 段恩帅 苏良湖 李贲 周筱洁


摘要:过硫酸钾能加快大分子降解,促进堆肥腐熟。在前期确定的过硫酸钾添加量(1.2%)基础上,进一步优化蔬菜废弃物堆肥的通风速率,并分析最优条件下微生物种群结构的变化。试验设置3 个通风速率VR1、VR2、VR4,分别为100、200、400 mL/min,VR1中加入堆体质量为15%的陶粒。结果表明,各组堆肥过程的理化性质差别不大,堆体温度>55 ℃的时间超过8 d,产品C/N为11.3~13.0,GI为91.9%~111.2%,均达到腐熟要求。各组第1天温度都超过55 ℃,升温快速,VR1的高温期温度高于其他组。堆肥过程中,各组可溶性有机物的紫外-可见吸收光谱特征值SUVA254、SUVA280值不断增加,最终VR1的特征值最高,堆肥的腐熟化程度较高。蔬菜废弃物堆肥的优势种群有厚壁菌门(Firmicutes)、放线菌门(Actinobacteriota)、变形菌门(Proteobacteria)、拟杆菌门(Bacteroidota),相对丰度总和达96%以上。对比分析VR1、不加过硫酸钾的常规处理(通风速率400 mL/min)发现,VR1 中Firmicutes的丰度显著低于常规处理,而放线菌门的丰度较高。在高温期,VR1由多样性的高温纤维素降解菌种群替代了常规处理中的优势属芽孢杆菌属(Bacillus)。整体上,常规处理细菌的种群演替滞后于VR1,VR1的堆肥进程更快。本研究证明,添加过硫酸钾的蔬菜废弃物在降低通风速率至100 mL/min并且加陶粒时,堆肥仍能够顺利进行,且堆体升温快速,有助于蔬菜废弃物堆肥的工艺优化。
关键词:好氧堆肥;蔬菜废弃物;通风速率;过硫酸钾;微生物
中图分类号:X705 文献标志码:A
文章编号:1002-1302(2023)24-0203-09
2020年我国的蔬菜产量超过7.2亿t,位居世界第一[1]。与此同时也产生了大量的蔬菜废弃物,其中大多被随意丢弃或进行填埋处置,造成严重的资源浪费和环境污染[2]。蔬菜废弃物营养成分高、有机质含量丰富,通过好氧堆肥,高温杀灭其中的虫卵、病原菌、草籽等,可使废弃物转化为有机肥,实现资源化利用[3]。
在实际的蔬菜好氧堆肥处理中,往往存在高温期短,物料降解不充分,进而影响堆肥腐熟度的问题[4],同时有废气排放需要处理。最近的研究发现,过硫酸钾用于堆肥可以促进木质纤维素等降解,使堆体快速升温,缩短堆肥周期,并能提高堆肥产品的腐熟度。Wang等发现,过硫酸钾用于牛粪好氧堆肥,可以大大提高纤维素、半纤维素、木质素的降解率,并促进溶解性有机质( DOM)的动态变化,加速腐殖酸的形成[5]。Mei等研究发现,过硫酸钾作为强氧化剂,可增加嗜热木质纤维素降解菌的丰度,提高高温期的最高温度和持续时间,减少堆肥过程中的氮素损失[6]。通过前期研究发现,过硫酸钾能加速蔬菜废弃物堆肥的升温,促进高温降解过程,添加量为1.2%时效果最好。
通风是影响好氧堆肥降解和腐熟过程的关键因素之一。通风速率过高,会带走堆体内的大量热量,不利于物料的高温降解;通风速率过低,不能满足微生物供氧,影响微生物活性;此外,通风速率还会对堆肥过程的废气排放量和堆体的碳氮损失造成影响[7-8]。根据堆肥原料的不同,不同堆肥研究選用的通风速率差别很大;[JP3]同样,在关于蔬菜堆肥的研究中,通风速率的设定值变化也较大。张陆等在甜瓜秧和鸡粪堆肥中选择的通风速率为0.01 m3/(m3·min)[9]。杨岩等在蔬菜废弃物堆肥研究中选择的通风速率为 0.27 m3/(m3·min)[10]。刘文杰等使用60 L堆肥装置,设定通风量为25 L/min[11]。本试验在前期研究确定的过硫酸钾添加量(1.2%)条件下,按一般通风速率适当递减通风量,进一步研究蔬菜废弃物添加过硫酸钾堆肥体系中,是否可以减少通风速率而不影响堆肥腐熟,旨在减少堆肥过程的动力消耗和废气处理量;同时分析最优条件下微生物的种群特征,通过与常规堆肥条件对比分析,探讨低通风速率处理蔬菜废弃物在过硫酸钾协同陶粒作用下堆肥体系中微生物种群的变化,以期为蔬菜废弃物快速好氧堆肥工艺优化和蔬菜废弃物就地堆肥处理技术的推广提供参考。
1 材料与方法
1.1 试验材料和装置
选取包菜叶、莴笋皮2种产生量较大的蔬菜废弃物作堆肥材料,这些材料取自苏州科技大学附近的学府花苑菜市场。用玉米秸秆、玉米粉、风干牛粪、陶粒作堆肥材料的调理剂,调节材料的结构和碳氮比(C/N)。牛粪、秸秆取自于连云港市东海农场,玉米粉购自学府花苑菜市场,陶粒购自淘宝网某园艺用品旗舰店,粒径0.8~1.0 cm,过硫酸钾(分析纯)购于国药集团化学试剂有限公司。堆肥材料的基本理化性质见表1,堆肥反应器选用50 L塑料桶[12]。在桶底部放置不锈钢架,其上放铁丝网用于承托堆肥材料并保证通风布气均匀。桶的底部、顶部分别有进气孔、排气孔。堆肥桶周围包裹保温材料,防止热量散失。
1.2 堆肥试验方法
将各种堆肥材料破碎至2~3 cm备用。按照初始含水率60%、C/N 20混合各种堆肥原料,每个处理混合材料湿质量为5.50 kg(含包菜叶2.00 kg、莴笋皮2.00 kg、牛粪0.24 kg、玉米粉 0.58 kg、玉米秸秆0.68 kg),过硫酸钾的添加量为1.2%(湿基添加量)。参考已有研究的适宜通风速率[12-13],设置3个通风速率VR1、VR2、VR4,分别为100、200、400 mL/min,连续通风。对VR1低通风速率条件的处理,添加湿质量15%的陶粒,以保证空气传质和扩散[14]。研究微生物变化的常规处理堆肥条件为不加过硫酸钾,通风速率为400 mL/min。
堆肥周期为50 d,在堆肥后1、3、6、10、15、21、35、50 d翻堆并采集样品。取样时,在不同深度多点采集混合,以保证样品具有代表性。每次取100 g鲜样于-20 ℃保存;另取100 g鲜样,风干后破碎,过80目筛,保存用于后续分析。每个样品测3个平行样。
1.3 测定指标与分析方法
用风干样品测定固体总有机碳(total organic carbon,TOC)含量、总氮(total nitrogen,TN)含量。湿样用于pH值、电导率(electrical conductance,EC)、种子发芽指数(seed germination index,GI)、可溶性有机碳(dissolved organic carbon,DOC)含量、紫外-可见吸收光谱特征值(SUVA254、SUVA280)的测定及微生物多样性分析。
使用电子测温计实时监测堆体温度,测温计的防水探头置于堆肥材料的中部。每天09:00和 18:00 记录堆体温度和环境温度。样品含水率采用105 ℃ 烘干法测定[12]。
取湿样10 g与去离子水按1 g ∶10 mL的比例混合,室温振荡30 min后过滤,获得浸提液Ⅰ。 浸提液Ⅰ用于测定pH值、EC值、GI。pH值、EC值分别用雷磁pHS-3C便携式pH计、雷磁DDB-303A便携电导率仪测定[12]。选用颗粒饱满、无破损的白菜种子测定GI:取10 mL浸提液浸润种子,在 25 ℃ 恒温下避光培养48 h,测量种子发芽数量和根长,按照式(1)计算GI[15]。以蒸馏水培养作为空白对照。
将浸提液Ⅰ在7 000 r/min条件下离心 7 min,再用0.45 μm滤膜过滤,获得浸提液Ⅱ。将 浸提液Ⅱ稀释50倍,用流动分析仪(Vario TOC select,Elementar,德国)测定DOC含量。同时使用紫外分光光度仪测定浸提液Ⅱ稀释10倍后在波长254、280 nm处的吸光度D254 nm、D280 nm,D254 nm、D280 nm与DOC含量的比值记为SUVA254、SUVA280[16]。
固体样品的TOC、TN含量分别按照NY/T 525—2021《有机肥料》标准中的重铬酸钾容量法、凯式定氮法测定。样品的碳氮比(C/N)是TOC(总有机碳)与TN(总氮)的比值。
1.4 微生物多样性分析
对高温期温度最高的VR1和常规堆肥组,在堆体温度变化明显的堆肥后3、6、10、15、21 d取样,分析细菌种群多样性。样品DNA的提取使用土壤DNA提取试剂盒(Omega Bio-Tek,Inc.,美国)。选用引物515F(5′-GTGCCAGCMGCCGCGG-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′),扩增16S RNA的V4区域。PCR反应体系为Trans Start Fast Pfu DNA Polymerase 20 μL。PCR反应条件为:95 ℃预变性 3 min;95 ℃变性30 s,53 ℃退火30 s,72 ℃延伸 45 s,共29个循环;72 ℃延伸10 min。样品测序由上海美吉生物医药科技有限公司完成,高通量测序使用Illumina MiSeq平台进行(Majorbio Bio Pharm Technology Co.,Ltd.)。
1.5 数据处理与分析
试验数据采用SPSS 25.0软件进行单因素方差分析和相关性分析,显著性差异设定为P<0.05。采用Origin 2021Pro软件绘制变化趋势图。微生物群落的多样性和差异性分析在Majorbio在线云平台(www.majorbio.com)上完成。
2 结果与分析
2.1 堆肥过程理化参数的变化
2.1.1 温度变化 4组处理的温度变化规律类似(图1)。堆肥初期,4组均快速升温,堆肥后 1 d温度均超过55 ℃,高溫期(>45 ℃)持续12 d左右。堆肥后10~15 d,堆体温度快速降低,之后逐渐接近室温。 VR1、VR2、VR4的最高温出现在堆肥后4 d,分别为67.3、64.3、65.8 ℃;VR1的温度最高,VR2的略低。常规处理于堆肥后 7 d达到最高温度 64.1 ℃。4组处理55 ℃以上时间在8 d左右,达到了堆肥化标准(GB/T 36195—2018《畜禽粪便无害化处理技术规范》)的腐熟要求。
2.1.2 pH值和EC值的变化 由图2-a可知,不同通风速率组的pH值变化规律一致,数值接近。堆肥的pH值呈先上升后缓慢下降的趋势,VR1、VR2、VR4这3个处理组的pH值由起始的5.0~5.3上升至35 d时的8.77~9.03,再缓慢降至 7.8~8.2;常规处理的pH值由起始的4.77上升至35 d时的9.23,再缓慢降至8.14。堆肥结束后,各组pH值均达到了成熟堆肥产品的要求[17]。由图2-b可知,堆肥期间的EC值呈先上升再波动下降的趋势,VR1、VR2、VR4这3组的EC值由初始的1.95~2.35 mS/cm,升至堆肥结束时的3.67~3.88 mS/cm,常规处理的EC值由起始的1.6 mS/cm上升至21 d的3.88 mS/cm,再缓慢降至3.17 mS/cm。通常堆肥的EC值小于4.00 mS/cm时,对植物生长无毒害作用[18]。因此,所有处理的产品EC值均符合堆肥腐熟的要求。
2.2 堆肥过程中碳、氮元素的变化
由图3-a可知,各组的总有机碳(TOC)含量整体呈下降趋势,VR1、VR2、VR4这3个处理组的TOC含量由堆肥前期最高的435.00~465.60 g/kg降至最终的378.85~402.66 g/kg,常规处理的TOC由前期最高的471.65 g/kg降至最终的405.22 g/kg。各组间TOC含量差异不显著(P>0.05)。由图3-b可知,各组的水溶性有机碳(DOC)含量整体呈下降趋势,VR1、VR2、VR4这3组由初始的1.65~2.11 g/kg下降至0.76~1.21 g/kg。常规处理的DOC含量由初始的1.90 g/kg下降至最终的 1.51 g/kg。由图3-c可知,各组的总氮(TN)含量整体呈先降低再升高的趋势。在堆肥后0~5 d,VR1、VR2、VR4这3[JP3]个处理组的TN含量由初始的25.96~28.97 g/kg下降至21.98~23.15 g/kg后,最终上升至结束时的31.08~34.25 g/kg。在堆肥后0~10 d,常规处理的TN含量由初始的22.58 g/kg上升至33.23 g/kg后,下降至结束时的30.08 g/kg。由图3-d可知,各组的碳氮比(C/N)都有明显降低的趋势,VR1、VR2、VR4这3组的C/N由初始的13.9~15.5降到堆肥结束时的11.3~13.0,常规处理的C/N由初始的17.4下降至结束时的13.5。一般研究认为,当堆肥C/N小于17时,堆肥达到腐熟[19],可见本研究中各组堆肥均达到腐熟要求。
2.3 堆肥腐熟度指数的变化
2.3.1 种子发芽指数变化 由图4可知,除了在堆肥过程中的异常高值外,各组的种子发芽指数(GI)随堆肥的进行逐渐升高。VR1、VR2、VR4这3组的GI从初始的10.0%~30.8%升至堆肥结束时的91.9%~111.2%,常规处理的GI由初始的10.0%上升至结束时的82.1%。
2.3.2 水溶性有机物的紫外-可见光谱特性 由图5可知,在堆肥过程中,各处理DOM的紫外-可见吸收光谱特征值SUVA254、SUVA280的变化规律一致,均呈现不断上升的趋势,组间差异不显著(P>0.05)。其中SUVA254从最初的0.144~0.229上升至堆肥结束时的0.458~0.626(图5-a);SUVA280从初始的0.126~0.196上升至堆肥结束时的0.400~0.528(图5-b)。
2.4 堆肥过程中微生物种群的变化
2.4.1 门水平的种群变化和演替 由表2可知,样品的测序文库覆盖率均超过0.99,测序结果代表样本的真实情况。CK的Ace指数和Chao指数呈波动变化,而VR1整体呈增加的变化规律。与CK相比,过硫酸钾的添加减少了VR1堆肥初期的细菌物种数量,但堆肥后6 d VR1的物种数量均比CK高。根据Shannon指数可知,2组群落多样性在降温期都开始显著增加。CK的群落多样性先增加后减小,VR1的群落多样性整体呈增加的趋势并在堆肥后期明显高于CK。而Simpson指数则有不同的变化规律,2组群落从堆肥后6 d开始均降低,显现出部分种群的优势度增大的趋势。
2组在门水平的优势种群主要有厚壁菌门(Firmicutes)、放线菌门(Actinobacteriota)、变形菌门(Proteobacteria)、拟杆菌门(Bacteroidota),相对丰度总和达96%以上(图6)。高温期,2组群落的种群结构差异显著。厚壁菌门是常规处理在高温期的最优势种群,堆肥后3、6 d的相对丰度分别为81.7%、98.9%。而在堆肥后3 d,放线菌门为VR1群落的最优势种群(58.5%);堆肥后6 d,厚壁菌门变为其绝对优势种群(92.5%)。2组群落的堆体温度从堆肥后10 d开始下降,组间的种群结构差异随之变小。整体上,降温后厚壁菌门的相对丰度降低,逐渐降至30%以下;放线菌门、变形菌门、拟杆菌门的相对丰度增加,放线菌门稳定在35%以上,变形菌门在堆肥后21 d的相对丰度增至33%。拟杆菌门的相对丰度较低,除了VR1群落在堆肥后 21 d,其余均低于11.4%。
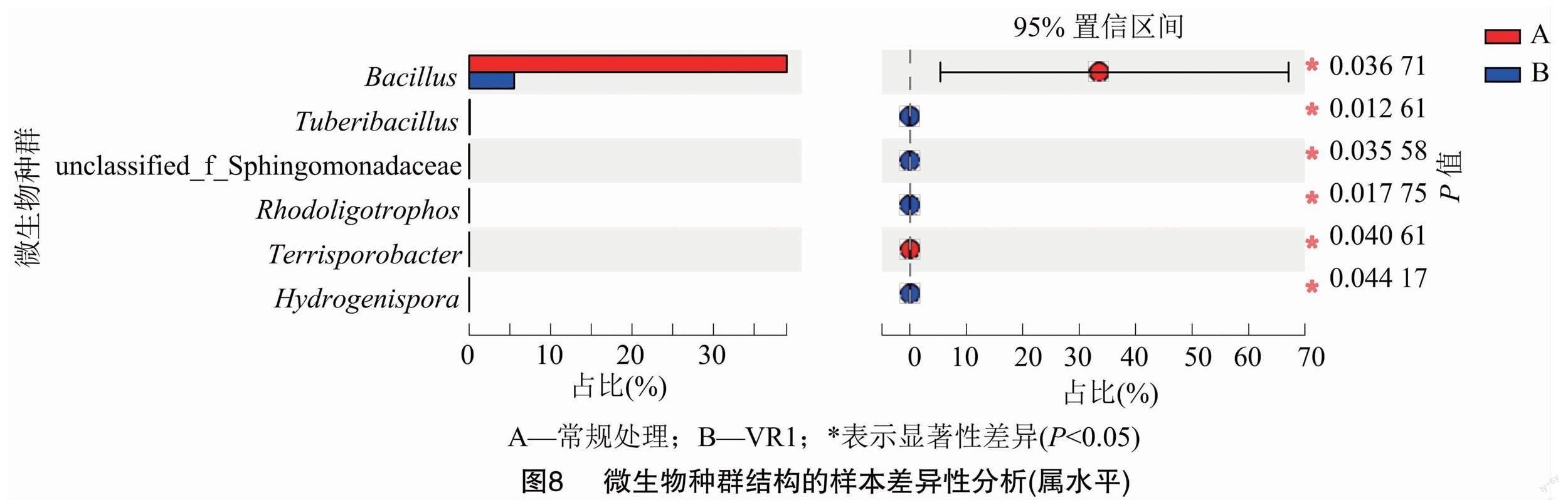
2.4.2 属水平的种群变化和演替 常规处理和VR1在高温期的优势属完全不同,降温期2组优势种群的多样性增加,且优势种群的组间差别变小(图7)。高温期,常规处理组Bacillus占绝对优势,堆肥后3、6 d的相对丰度分别为68.8%、92.6%,其他属的丰度均很低。Bacillus(Firmicutes)是重要的嗜热分解菌,具有广泛的生化活性,通常是堆肥全过程的优势种群[20-21]。高温期,VR1中细菌优势种群的多样性大大高于对照。堆肥后3 d的优势种群包括Saccharopolyspora、norank_f_Pseudonocardiaceae、Rhodococcus、Gordonia、Thermopolyspora、 Bacillus、Saccharomonospora。VR1堆肥后6 d的优势种群变为Firmicutes(92.5%),而且VR1中Firmicutes的多样性明显高于常规处理组。除了芽孢杆菌属,优势属还包括Symbiobacterium、Ureibacillus、Caldibacillus、Aeribacillus、Planifilum、Thermobacillus、unclassified_c_Clostridia,均属于Firmicutes。
堆体温度降低后,2组群落的种群结构差别变小。高温双岐菌属(Thermobifida)在2组群落中都是最优势种群,在VR1堆肥后15 d的相对丰度高达49.1%,对Thermobifida更有利。在常规处理中,Thermobifida相对丰度较高且有波动(6.9%~26.3%),其他的优势种群在降温期演替的顺序为:由堆肥后10 d的Bacillus、 Aeribacillus、 Caldibacillus变为堆肥后15 d的Bacillus、Saccharomonospora、Saccharopolyspora,再变为堆肥后21 d的Saccharomonospora、Pseudoxanthomonas、Bacillus。
3 讨论
3.1 通风速率对堆肥过程理化性质的影响
本研究参考已有研究,选用常规通风速率为400 mL/min[12-13],与其他蔬菜堆肥研究相比,通风速率偏低。刘文杰等在对蔬菜废弃物的堆肥研究中,对18~24 kg物料的通风量设为25 L/min[11]。本试验结果表明,在添加1.2%过硫酸钾条件下,200 mL/min通风条件也能顺利升温,只是高温期温度稍低,而通风速率100 mL/min加陶粒的温度是3组中最高的。这可能与低通风速率和陶粒的添加有关,低通风速率减少了热量散失,而陶粒增加物料的孔隙率,有利于空气在堆肥物料中的扩散[14,22],确保低通风速率条件下降解过程对氧气的需求。在提高材料孔隙率条件下,添加1.2%过硫酸钾,在降低通风速率75%时,物料降解仍能顺利进行,而且能实现堆体快速升温。常规处理的最高温度低于其他处理,且温度峰值的出现明顯滞后于其他处理,表明过硫酸钾能在一定程度上加速物料进入高温期。
堆肥物料的容重间接反映了堆体的孔隙度,因此容重对堆体的透气性影响极大。容重越大,孔隙度越小,堆体内的氧气输送受阻,越不利于提高堆肥效率。本研究堆肥物料的初始容重估算为 0.16~0.22 g/cm3。秦翠兰等在牛粪堆肥研究中测定的容重变化范围为0.4~1.3 g/cm3,初始容重为0.8~1.2 g/cm3[23]。相较于此研究,本研究物料的初始容重较小,孔隙较大,空气扩散条件好,对低通风速率条件较为适宜。
堆肥过程中,pH值会随着物质的转化和微生物的活动而变化,同时也是影响堆肥进行的重要因素。pH值过高或过低,都不利于堆体中微生物的生长,pH值在6.5~8.5比较合适。各处理组起始的pH值在5.0~5.3之间,为弱酸性,可能是由于初始微生物降解糖类等易降解物质,产生大量的有机酸,使得pH值下降[24]。堆肥的前6 d,VR1、VR2、VR4的pH值均快速上升,这是由于高温阶段,物料剧烈分解使大量有机氮矿化和有机酸降解而造成的;而常规处理在堆肥后3 d的pH值降低,与其他3组处理显著不同(P<0.05),表明添加过硫酸钾有利于避免易腐有机物高温降解初期的酸化问题,有利于堆肥顺利升温。堆肥后35 d,堆肥进入腐熟阶段,微生物矿化作用减弱,硝化细菌开始活跃,将铵态氮通过硝化作用转变为硝态氮,pH值又有所降低[25]。
堆肥产品的EC值能够反映产品中的盐含量。堆肥前期有机物被不断降解,铵盐、硝酸盐等不断释放,同时由于物料降解引起质量损失,导致盐分浓缩,最终使堆肥的EC值升高。整体上EC值表现为VR2>VR4>VR1,降低通风速率可降低堆肥过程中物料的EC值,最终产品的EC值差别不大(P>0.05)。常规处理的EC值整体低于其他3组,是因为没有添加过硫酸钾,盐含量低。整体上,降低通风量对堆肥过程的进行和理化性质的变化没有不利影响。
3.2 堆肥过程中碳、氮元素的变化
随着有机质被微生物降解和利用,堆肥物料中碳含量会逐渐降低[26]。各组初始的总有机碳含量(TOC)较高,这可能是由于初期物料还处于原始结构状态,不易风干,导致相同风干时间条件下,初始样品的含水率偏高,造成总有机碳(TOC)的计算结果偏低。
可溶性有机碳含量(DOC)是反映堆肥穩定性的重要参数之一。DOC是能够被微生物利用、合成自身生命体的最直接碳源,强烈影响着堆肥分解过程。堆肥中后期出现DOC升高和波动现象,可能是由物料中纤维素等难降解组分引起的。有研究认为,有些DOC是腐殖酸形成的前体物,腐殖酸的形成会使堆肥后期的DOC含量降低[25]。在堆肥过程中,VR1的DOC含量基本上低于其他组。堆肥结束时,各组DOC含量小于初始含量,分别为0.757、1.157、1.208 g/kg,均达到堆肥稳定标准化(<17 g/kg)[27]。
随着堆肥的进行,各组的总氮(TN)含量呈现较大的波动。高温期,各组TN含量先降低再升高。高温期TN含量的降低可能与有机氮的快速降解有关,有机氮降解产生的氨态氮在高温条件下以氨的形式挥发,使TN含量降低。堆肥后期,微生物降解利用木质纤维素,干物质降解损失,浓缩效应使物料的TN含量又增加[28-29]。堆肥结束时,产品的TN含量增加明显(P<0.05),比初始含量提高20%左右。VR1最终的TN含量低于其他组,这可能与其高温条件和高孔隙率导致的氨挥发有关。
3.3 通风速率对堆肥腐熟的影响
种子发芽指数(GI)是综合评价堆肥产品腐熟的指标。当GI达到腐熟要求时,堆肥产品对植物生长的毒性已消除。与牛粪秸秆等材料的堆肥过程相比,本研究GI达到腐熟要求的时间较晚,这可能是堆肥原料的性质差异造成的[12,30]。堆肥结束时,各组GI分别为97.5%、111.2%、91.9%,均达到NY/T 525—2021《有机肥料》GI大于70%的腐熟要求。堆肥中后期,VR1的GI高于VR2、VR4,且3组处理的GI均高于常规处理,此工艺条件对堆肥产品的植物毒性没有不利影响。
水溶性有机物DOM的紫外-可见吸收光谱特性被证明可以用于评价堆肥的腐熟度[31]。本研究选择紫外-可见吸收光谱特征值SUVA254、SUVA280来评判堆肥的腐熟度。SUVA254与有机物中C—C键化合物组成有关,而SUVA280则受有机物芳构化程度及分子量大小影响,二者的值越大,腐殖化程度越高[32]。随着堆肥的进行,各组SUVA254、SUVA280不断增加,表明各组在堆肥的中后期腐殖化程度不断提高。娄义晟等对鸡粪和稻壳混堆的研究表明,SUVA254、SUVA280值的增加与堆体的稳定性呈正相关,可作为堆肥腐熟的辅助性评价指标[33]。按照此腐熟标准判断,本研究堆肥结束时各组的SUVA254、SUVA280均呈上升趋势,表明堆肥的腐熟度提高。VR1最终的SUVA254、SUVA280高于VR2、VR4,表明在该条件下堆肥的腐熟化程度较高。
3.4 微生物种群差异性分析
VR1和常规处理在细菌种群的多样性和种群结构方面均有明显差异。VR1增加了细菌的丰富度和多样性,且高温期的优势种群与常规处理不同。
在门的水平,2组群落中Firmicutes、Actinobacteriota的差异显著。VR1中Firmicutes的相对丰度明显低于常规处理。尤其在高温期开始,VR1群落在堆肥后3 d的Firmicutes相对丰度只有26.7%,而常规处理为81.7%。而VR1对Actinobacteriota的生长和活动有利,Actinobacteriota是VR1的最优势种群,除堆肥后6 d外,相对丰度达35%~70%。Actinobacteriota可以通过分泌相关酶来降解木质纤维素,是堆肥过程的主要种群之一,其数量一般在高温期过后增加,被认为是堆肥腐熟的指示菌[34]。而VR1在高温期Actinobacteriota也有很高的相对丰度(堆肥后3 d为58.5%)。
在属的水平,常规处理组高温期Bacillus占绝对优势(相对丰度68.8%~92.6%)。Bacillus(Firmicutes)是重要的嗜热分解菌,具有广泛的生化活性,通常是堆肥全过程的优势种群[35]。而VR1在堆肥后 3 d的优势种群多为嗜热放线菌,其中最优势种群Saccharopolyspora的相对丰度为27.4%。VR1对高温放线菌有利,这可能由于堆肥初期过硫酸钾能够氧化易降解有机酸,使堆肥材料的pH值快速升高(见“2.1.2”节),有利于放线菌的繁殖。VR1在堆肥后6 d的优势种群变为Firmicutes(92.5%),但其中除了Bacillus,还包括Symbiobacterium、Ureibacillus等多种高温耐热菌,这些属具有纤维素、半纤维素降解功能,有利于有机氮、有机酸快速降解[36-37]。VR1在高温期Bacillus的相对丰度不超过10%,远低于常规处理组的70%~90%。与常规处理组相比,VR1在高温期的Bacillus种群明显受到抑制(图8),其他纤维素半纤维素降解高温厚壁菌及有机物快速降解菌取代了Bacillus。这种变化可能是堆肥初期过硫酸钾对有机物氧化降解造成的。过硫酸钾对有机物的分解取代Bacillus种群的功能,减少Bacillus的降解基质,抑制其生长和活动。
2组对比发现,VR1与常规处理组的优势种群更替顺序一致,但常规处理的种群演替滞后于VR1,表明VR1的堆肥进程更快,可以促进物质降解和腐熟。由于VR1的处理包括1.2%过硫酸钾、15%陶粒和100 mL/min通风速率,三者协同作用,单一因素对微生物的影响还需要进一步探究。
4 结论
蔬菜废弃物在添加1.2%过硫酸钾的条件下堆肥50 d,通风速率100、200、400 mL/min的组间理化性质差别不大。各组高温期(>45 ℃)持续13 d左右,产品pH值为7.8~8.2,EC值为3.67~3.88 mS/cm,C/N为11.3~13.0,GI为91.9%~111.2%,均达到腐熟,符合NY/T 525—2021《有机肥料》的要求。
各处理组均升温较快,堆肥后 1 d温度都超过了 55 ℃。VR1的高温期温度略高于另外2组,表明添加1.2%过硫酸钾并提高材料孔隙率条件下,通风速率为100 mL/min时堆肥仍能顺利进行,而且升温速度快。VR1最终的TN含量低于其他组,这可能与其高温条件和高孔隙率导致的氨挥发有关。随着堆肥的进行,各组DOM的SUVA254、SUVA280值整体均不断增加,表明堆肥的腐熟度提高。
蔬菜废弃物堆肥的优势种群在门水平有Firmicutes、Actinobacteriota、Proteobacteria、Bacteroidota,相对丰度总和达96%以上。添加过硫酸钾对Firmicutes、Actinobacteriota影响明显,尤其在堆肥初期降低了Firmicutes的豐度,而对Actinobacteriota的生长和活动有利。堆肥后3 d,常规处理中的优势种群是Bacillus,而VR1为多种高温纤维素降解菌种群。整体上,常规处理中微生物的种群演替滞后于VR1,VR1的堆肥进程更快,VR1条件有利于有机质的降解和腐熟。由于VR1处理添加1.2%过硫酸钾、15%陶粒并选用低通风速率100 mL/min,单一因素对微生物的影响还不够明晰。今后的研究需要在单因素试验的基础上,进一步探究过硫酸钾、陶粒及通风速率单独作用下对蔬菜废弃物堆肥过程微生物的影响。
参考文献:
[1]袁 远,张金辉,李银月,等. 蔬菜废弃物好氧堆肥处理研究进展[J]. 江苏农业科学,2022,50(19):29-35.
[2]陆晓林,杨玉欣,洪春来,等. 猪粪辅料促进茄果类蔬菜废弃物堆肥品质的微生物机理[J]. 农业环境科学学报,2022,41(5):1097-1107.
[3]刘 宇,席运官,高 丽,等. 有机农场蔬菜废弃物堆肥效果研究[J]. 江苏农业科学,2021,49(1):182-186.
[4]林向宇,孟寒玉,王晓君,等. 分散式堆肥设备结合二次堆肥对蔬菜废弃物的处理效能及能耗分析[J]. 环境卫生工程,2022,30(5):40-46.
[5]Wang S S,Meng Q R,Zhu Q H,et al. Efficient decomposition of lignocellulose and improved composting performances driven by thermally activated persulfate based on metagenomics analysis[J]. Science of the Total Environment,2021,794:148530.
[6]Mei J,Li B,Su L H,et al. Effects of potassium persulfate on nitrogen loss and microbial community during cow manure and corn straw composting[J]. Bioresource Technology,2022,363:127919.
[7]杨 佳,王国英,唐若兰,等. 生物炭和菌剂对羊粪微好氧堆肥腐熟度和温室气体排放的影响[J]. 农业工程学报,2022,38(10):224-231.
[8]张相锋,王洪涛,聂永丰. 通风量对蔬菜和花卉废弃物混合堆肥的影响[J]. 农业工程学报,2005,21(10):134-137.
[9]张 陆,曹玉博,王惟帅,等. 鸡粪添加对蔬菜废弃物堆肥腐殖化过程的影响[J]. 中国生态农业学报(中英文),2022,30(2):258-267.
[10]杨 岩,孙钦平,李 妮,等. 不同过磷酸钙添加量对蔬菜废弃物堆肥的影响[J]. 农业资源与环境学报,2017,34(1):66-72.
[11]刘文杰,王黎明,沈玉君,等. 碳氮比对蔬菜废弃物好氧发酵腐熟度及臭气排放的影响[J]. 环境工程,2020,38(6):233-239.
[12]吴梦婷,梅 娟,苏良湖,等. 硫酸亚铁和过磷酸钙对牛粪秸秆混合堆肥氮损失和腐殖化的影响[J]. 生态与农村环境学报,2020,36(10):1353-1361.
[13]逯延军. 城市生活垃圾与城市污泥好氧共堆肥的工艺特性及机理研究[D]. 上海:同济大学,2008:17-18.
[14]李 旭,路明艺,师晓爽,等. 多孔填充剂促进牛粪秸秆高温恒温堆肥有机质降解减少氮损失[J]. 农业工程学报,2018,34(增刊1):132-137.
[15]Yao Y L,Zhu F X,Hong C L,et al. Utilization of gibberellin fermentation residues with swine manure by two-step composting mediated by housefly maggot bioconversion[J]. Waste Management,2020,105:339-346.
[16]李鸣晓,何小松,刘 骏,等. 鸡粪堆肥水溶性有机物特征紫外吸收光谱研究[J]. 光谱学与光谱分析,2010,30(11):3081-3085.
[17]吕 潇,常瑞雪,毛瑞鑫,等. 添加炭基材料对蔬菜废物好氧堆肥进程和腐熟度的影响[J]. 环境工程学报,2015,9(3):1422-1426.
[18]Bai L,Deng Y,Li J,et al. Role of the proportion of cattle manure and biogas residue on the degradation of lignocellulose and humification during composting[J]. Bioresource Technology,2020,307:122941.
[19]鮑艳宇,周启星,颜 丽,等. 畜禽粪便堆肥过程中各种氮化合物的动态变化及腐熟度评价指标[J]. 应用生态学报,2008,19(2):374-380.
[20]Jurado M,López M J,Suárez-Estrella F,et al. Exploiting composting biodiversity:study of the persistent and biotechnologically relevant microorganisms from lignocellulose-based composting[J]. Bioresource Technology,2014,162:283-293.
[21]Nakasaki K,Araya S,Mimoto H.Inoculation of Pichia kudriavzevii RB1 degrades the organic acids present in raw compost material and accelerates composting[J]. Bioresource Technology,2013,144:521-528.
[22]陈 晨,谭 昊,金海红,等. 村镇易腐垃圾堆肥过程的氮持留行为研究[J]. 环境污染与防治,2022,44(5):573-578.
[23]秦翠兰,王磊元,刘 飞,等. 木醋液添加牛粪堆肥过程容重时空层次变化及水分时空变化的灰色关联分析[J]. 江苏农业科学,2016,44(4):482-485.
[24]常瑞雪. 蔬菜废弃物超高温堆肥工艺构建及其过程中的氮素损失研究[D]. 北京:中国农业大学,2017:47-49.
[25]张 惠,段亚军,王小敏,等. 腐熟菌剂和辅料组合对蔬菜废弃物堆肥进程和腐熟度的影响[J]. 中国蔬菜,2021(9):69-77.
[26]鲍艳宇,颜 丽,娄翼来,等. 鸡粪堆肥过程中各种碳有机化合物及腐熟度指标的变化[J]. 农业环境科学学报,2005,24(4):820-824.
[27]Bernal M P,Alburquerque J A,Moral R.Composting of animal manures and chemical criteria for compost maturity assessment. A review[J]. Bioresource Technology,2009,100(22):5444-5453.
[28]程 丰. 外源添加剂减少好氧堆肥过程氮素损失的效果研究[D]. 无锡:江南大学,2021:14-17.
[29]古君禹,王秋君,孙倩,等. 农林废弃物堆肥产物复配黄瓜育苗基质配方筛选[J]. 江苏农业学报,2022,38(5):1238-1247.
[30]Zhan Y B,Zhang Z Y,Ma T T,et al. Phosphorus excess changes rock phosphate solubilization level and bacterial community mediating phosphorus fractions mobilization during composting[J]. Bioresource Technology,2021,337:125433.
[31]唐朱睿,席北斗,何小松,等. 猪粪堆肥过程中水溶性有机物结构演变特征[J]. 光谱学与光谱分析,2018,38(5):1526-1532.
[32]赵 越,魏雨泉,李 洋,等. 不同物料堆肥腐熟程度的紫外-可见光谱特性表征[J]. 光谱学与光谱分析,2015,35(4):961-965.
[33]娄义晟,王宇蕴,吴 昊,等. 堆肥过程中水溶性有机物的SUVA254、SUVA280值的变化及其与腐熟度进程的关系[J]. 土壤通报,2020,51(3):641-646.
[34]Meng Q X,Yang W,Men M Q,et al. Microbial community succession and response to environmental variables during cow manure and corn straw composting[J]. Frontiers in Microbiology,2019,10:529.
[35]Zhao Y,Lu Q,Wei Y Q,et al. Effect of Actinobacteria agent inoculation methods on cellulose degradation during composting based on redundancy analysis[J]. Bioresource Technology,2016,219:196-203.
[36]程旭艳,霍培书,尚晓瑛,等. 堆肥中高温降解菌的筛选、鉴定及堆肥效果[J]. 中国农业大学学报,2012,17(5):105-111.
[37]朱 屹,李俊良,焦 博,等. 整合宏组学方法研究番茄与玉米秸秆共堆肥生境中的关键微生物及其功能[J]. 福建农业学报,2020,35(7):764-772.
猜你喜欢
科技创新与应用(2017年3期)2017-02-18
湖北农业科学(2016年20期)2017-02-15
中小企业管理与科技·下旬刊(2016年11期)2016-11-18
求知导刊(2016年26期)2016-10-31
中国实用医药(2016年15期)2016-05-24
科技视界(2016年9期)2016-04-26
科技视界(2016年7期)2016-04-01
天津农业科学(2015年5期)2015-05-30
纺织导报(2015年2期)2015-04-07
