
Metformin promotes angiogenesis and functional recovery in aged mice after spinal cord injury by adenosine monophosphate-activated protein kinase/endothelial nitric oxide synthase pathway
2023-02-24 05:25JinYunZhaoXiaoLongShengChengJunLiTianQinRunDongHeGuoYuDaiYongCaoHongBinLuChunYueDuanJianZhongHu
中国神经再生研究(英文版) 2023年7期
Jin-Yun Zhao ,Xiao-Long Sheng,Cheng-Jun Li,Tian Qin,Run-Dong He,Guo-Yu Dai,Yong Cao,Hong-Bin Lu,,Chun-Yue Duan,,Jian-Zhong Hu,
Abstract Treatment with metformin can lead to the recovery of pleiotropic biological activities after spinal cord injury.However,its effect on spinal cord injury in aged mice remains unclear.Considering the essential role of angiogenesis during the regeneration process,we hypothesized that metformin activates the adenosine monophosphate-activated protein kinase/endothelial nitric oxide synthase pathway in endothelial cells,thereby promoting microvascular regeneration in aged mice after spinal cord injury.In this study,we established young and aged mouse models of contusive spinal cord injury using a modified Allen method.We found that aging hindered the recovery of neurological function and the formation of blood vessels in the spinal cord.Treatment with metformin promoted spinal cord microvascular endothelial cell migration and blood vessel formation in vitro.Furthermore,intraperitoneal injection of metformin in an in vivo model promoted endothelial cell proliferation and increased the density of new blood vessels in the spinal cord,thereby improving neurological function.The role of metformin was reversed by compound C,an adenosine monophosphate-activated protein kinase inhibitor,both in vivo and in vitro,suggesting that the adenosine monophosphate-activated protein kinase/endothelial nitric oxide synthase pathway likely regulates metformin-mediated angiogenesis after spinal cord injury.These findings suggest that metformin promotes vascular regeneration in the injured spinal cord by activating the adenosine monophosphateactivated protein kinase/endothelial nitric oxide synthase pathway,thereby improving the neurological function of aged mice after spinal cord injury.
Key Words:adenosine monophosphate-activated protein kinase/endothelial nitric oxide synthase pathway;angiogenesis;aged mice;compound C;metformin;spinal cord injury
Introduction
Traumatic spinal cord injury (SCI),when not fatal,frequently leads to permanent deficits in sensory and motor function.Despite major progress in neuro-therapeutic approaches for SCI,no clinically curative treatments are currently available.As the population ages,the social burden of older patients with SCI is rapidly growing (Devivo,2012;Singh et al.,2014).
Recently there has been increasing interest in understanding the effects of aging on the pathological changes that occur after SCI.SCI is characterized by acute mechanical injury,causing the rupture of blood vessels,which triggers a cascade of secondary injuries (Graumann et al.,2011;Figley et al.,2014).After trauma,blood vessels undergo significant remodeling,which is vital for the survival of neuronal tissue (Sapieha,2012;Ni et al.,2019).In addition,the structure of the microvasculature changes dramatically due to the induction of angiogenesis (Ng et al.,2011;Figley et al.,2014).New blood vessels grow into the lesion site to deliver nutrients that promote SCI repair (Figley et al.,2014).However,the effect of aging on vessel regeneration after SCI remains unclear.
The biguanide metformin has been used to treat type 2 diabetes and metabolic syndrome since the 1960s.More recently,increasing evidence has shown that metformin has important effects on cancer (Xu et al.,2015a)and polycystic ovary syndrome (Tao et al.,2018;Ali et al.,2019).Metformin is also cardioprotective (Calvert et al.,2008),suggesting that metforminbased therapy could be used to treat age-related diseases.In fact,studies have shown that metformin can delay the aging process (Barzilai et al.,2016;Piskovatska et al.,2020;Alhowail and Chigurupati,2022),reduce aging-related inflammation (Bharath et al.,2020),and positively modulate the gut (Pollak,2017).In addition,it has extensive effects in the central nervous system,showing benefit in the treatment of conditions such as Parkinson’s disease(Paudel et al.,2020),Huntington’s disease (Vázquez-Manrique et al.,2016),epilepsy (Sanz et al.,2021),and cerebral ischemia/reperfusion injury (Leech et al.,2019).Metformin is thought to modulate autophagy,inflammation,and neuronal apoptosis in the context of SCI in young rodent animals,which could help improve functional recovery (Wang et al.,2016;Zhang et al.,2017b;Song et al.,2021).Furthermore,metformin treatment not only ameliorates oxidative stress and stabilizes microtubules (Wang et al.,2020),but also prevents blood-spinal cord barrier breakdown by reducing the production of matrix metalloproteinase-9 and the amount of infiltrating neutrophils (Zhang et al.,2017a),reinforcing its potential therapeutic benefits in adult female SCI rats.However,it remains unclear whether metformin administration promotes robust vascular regeneration,thereby conferring neuroprotective effects and promoting functional recovery,especially in aged mice post-SCI,and what mechanisms may underlie these effects.
The present work focused on investigating whether angiogenesis is attenuated in aged mice after SCI.The results show that metformin affects blood vessel formation bothin vitroandin vivothrough the adenosine monophosphateactivated protein kinase/endothelial nitric oxide synthase (AMPK/eNOS)pathway.
Methods
Animals
Previous work has demonstrated that estrogen has an impact on angiogenesis after SCI (Samantaray et al.,2016;Ni et al.,2018),and thus we used male mice to exclude these effects.Two-month-old (young group,22–24 g,n=27)and 18-month-old C57BL/6J (aged group,32–38 g,n=132) male,specific pathogen-free mice were used for thein vivoexperiments,and another 10–12 C57BL/6 male mice (aged 8–12 weeks,22–26 g) were used forin vitroexperiments.All mice were acquired from the Experimental Animal Center of Xiangya Hospital,Central South University (license No.SYXK (Xiang) 2020-0019).All animal procedures were approved by the Animal Welfare and Ethics Committee of Xiangya Hospital,Central South University (approval No.2020sydw0374) on July 4,2020.Allin vivoexperiments were designed and reported according to the Animal Research: Reporting ofIn VivoExperiments(ARRIVE) guidelines (Percie du Sert et al.,2020) housed in separate cages under a 12/12-hour light/dark cycle in a temperature-controlled environment(temperature 22–24°C;humidity 60–80%),with free access to water and food.The animals were subjected to a surgical procedure mimicking SCI,and the outcomes were assessed in a blinded manner.To explore the differences between young and aged mice,young and aged SCI mice were randomly divided into young sham,young SCI,aged sham,and aged SCI groups.For the metformin treatment experiment,aged SCI mice were randomly divided into phosphate buffered saline (PBS) and metformin treatment groups.For the compound C administration experiment,aged SCI mice were randomly divided into sham,metformin only,and metformin plus compound C treatment groups.The study timeline is shown inAdditional Figures 1–3.
Contusive SCI model and experimental design
A modified Allen approach was used to construct the traumatic SCI mouse model (Allen,1911;Ni et al.,2019).Briefly,the mice were anestheti zed by intraperitoneal injection of xylazine (10 mg/kg;Topscience,Shanghai,China,Cat# T7046) and ketamine (80 mg/kg;Ketathesia,Henry Schein Animal Health,Dublin,OH,USA).An incision was made on the back midline,followed by laminectomy at the T10 level.Then,a 10-g weight was applied to the dorsal surface of the exposed spinal cord dorsal surface using a modified Allen weight-drop apparatus set to a vertical height of 15 mm.The mouse bladders were emptied manually twice a day until normal voiding activity was recovered.Metformin (MedChemExpress,Monmouth Junction,NJ,USA,Cat# HY-B0627) was diluted to 100 mg/mL.The injured mice were injected intraperitoneally with metformin (100 mg/kg per day) and compound C (20 mg/kg per day;MedChemExpress,Cat# HY-13418A),an AMPK inhibitor,from 3 days post-SCI until the mice were sacrificed.PBS (20 µL;Solarbio,Beijing,China,Cat# P1010) was used as the vehicle-only control.Behavioral parameters were evaluated by Basso Mouse Scale (BMS) and tacti le allodynia 24 hours after each daily dose of metformin and compound C.
Neurological assessment
The neurological assessment was based on motor and sensory tests post-SCI.The BMS hindlimb locomotor scale was used to assess recovery of hindlimb motor function before SCI and 1,3,7,14,21,28,35,42,and 56 days post-SCI,as previously described (Basso et al.,2006;Ni et al.,2019).The BMS scores ranged from 0 (complete paralysis) to 9 (normal hindlimb motor function).Two blinded,independent reviewers assessed hindlimb locomotor function,and the two scores were averaged to yield the final BMS score.
Von Frey filament test
The Von Frey filament test was carried out using Semmes-Weinstein monofilaments (Stoetling Company,Dale,IL,USA),as previously described(Chaplan et al.,1994).Briefly,the filament was used to stimulate the plantar surface of the hindpaw.Rapid withdrawal or lift ing of the stimulated hindpaw was considered a positive response.Paw withdrawal thresholds were measured as described previously (Ni et al.,2019).Tests were performed before surgery and 7,14,21,28,42,and 56 days post-SCI.
Electromyography
Motor-evoked potentials (MEPs) in the hindlimbs were assessed as described previously (Schlag et al.,2001).Briefly,56 days post-SCI mice were anestheti zed with xylazine (10 mg/kg) and ketamine (80 mg/kg).Then,stimulating electrodes (Chengdu Techman Instrument Co.,Ltd.,Chengdu,China) were placed on the skull over the cortical motor center,and recording electrodes were inserted in the contralateral hindlimb muscle.Electrical stimulation was performed at 3 V and 333 Hz and repeated every 2 seconds.The average amplitude of the MEP values and the latency period were recorded prior to SCI surgery and 56 days post-SCI.
Immunohistology
After anesthetization with xylazine (10 mg/kg) and ketamine (80 mg/kg),mice were perfused with normal saline and 4% paraformaldehyde (PFA) at different time points.The spinal cords were then collected and fixed with 4% PFA overnight,followed by immersion in a 30% sucrose solution and slicing into 16-µm transverse cryo-slices.For immunofluorescence staining,the spinal sections were incubated with anti-Ki67 (rabbit,1:1000,Abcam,Cambridge,UK,Cat# ab15580,RRID: AB_443209) and anti -CD31 (goat,1:100,R&D System,Inc.,Minneapolis,MN,USA,Cat# AF3628,RRID: AB_2161028)primary anti bodies overnight at 4°C.Then,the sections were incubated with secondary antibodies (donkey anti-goat IgG H&L (Alexa Fluor® 488),1:400,Abcam,Cat# ab150129,RRID: AB_2 687506;donkey anti-rabbit IgG H&L(Alexa Fluor® 594),1:400,Abcam,Cat# ab150076,RRID: AB_2782993) for 1 hour at 4°C.Next,the sections were washed with PBS and mounted with Vectashield DAPI Hardmount (Vector Laboratories,Burlingame,CA,USA) for nuclear visualization.A fluorescence microscope (Carl Zeiss Axio Observer Z1,Oberkochen,Germany) was used to observe five random fields of view from every section,with 10 sections analyzed for every group.The percentage of the newly formed blood vessels (CD31-positive) and double-stained proliferated vascular endothelial cells (CD31 and Ki67 double-positive) was calculated for each field of view and averaged to generate a final value for each section.Representative 8-µm transverse spinal cord slices at the injury epicenter (n=6 per group) were selected for hematoxylin-eosin staining to analyze the lesion size area,as previously described (Hu et al.,2016).
Three-dimensional microvascular visualization
The sample preparation for three-dimensional (3D) microvasculature imaging using an X-ray tomography microscope was performed as follows (n=6 per group).In brief,56 days post-SCI,young and aged mice from the SCI and sham groups were anestheti zed with xylazine (10 mg/kg) and ketamine (80 mg/kg),followed by perfusion with normal saline and 4% PFA and injection of a contrast medium mixture (Microfil,MV-122,Flow Tech,Carver,MA,USA) into the ascending aorta.Then,the spinal cords were harvested,fixed in 4% PFA,and prepared for 3D microvasculature scanning.A ZEISS Xradia 410 Versa (Carl Zeiss XRM,Pleasanton,CA,USA) was employed to perform high-resolution and contrast XRM non-destructive 3D submicron imaging using a unique dualstage process based on synchrotron-caliber optics followed by geometric magnification.The prepared spinal cord samples were scanned for 3 ms with the X-ray tube set to 40 kV and 10 kW.A pre-warm up procedure of 801 projections was performed,followed by 3001 projections of the target tissue.The image resolution was set at 3.25 µm/pixel.After scanning,all images were transformed into raw ti ff format.VG studio Max 3.0 soft ware (Volume Graphics,Heidelberg,Germany) was used to generate 3D images and analyze 3D vessel morphological parameters.
Footprint analysis
Footprint analysis was used to evaluate the hindlimb motor functional recovery before SCI surgery and 56 days post-SCI.Each animal’s front and hind paws were dyed blue and red,respectively,and the animal was then placed on the runway paper (7.5 cm × 100 cm),where they walked from one end to the other as trained.The footprints were then scanned and analyzed using Image-Pro Plus 7.0 soft ware (Media Cybernetics,Rockville,MD,USA).
Isolation and culturing of spinal cord microvascular endothelial cells
Primary mouse spinal cord microvascular endothelial cells (SCMECs) were isolated as described previously (Ruck et al.,2014).Briefly,the mice were anesthetized with xylazine (10 mg/kg) and ketamine (80 mg/kg) and then sacrificed by cervical dislocation (Ruck et al.,2014).The thoracic spinal cord was removed,minced,and digested with 0.1% (v/v) collagenase type II (Sigma-Aldrich,Burlington,MA,USA) in 10 mL Dulbecco’s modified Eagle medium (DMEM;Gibco,Invitrogen,Paisley,UK) for an hour at 37°C on an orbital shaker.To remove myelin,the digested spinal cord tissue was centrifuged,and the pellet was resuspended into 20% bovine serum albumin-DMEM (20 mL,w/v),followed by 20 minutes of centrifugation at 3000 ×gand 4°C.Next,the pellet was resuspended in 0.1% collagenase/dispase (Sigma,Burlington,MA,USA) in DMEM (10 mL),followed by 1 hour of digestion on the orbital shaker at 37°C.The solution was then centrifuged and,after rinsing,the pellet was resuspended twice with DMEM.The isolated SCMECs were inoculated onto endothelial cell medium plates supplemented with low-glucose DMEM,heparin (MedChemExpress,Cat# HY-17567A),20% fetal bovine serum (Gibco),basic fibroblast growth factor (Peprotech,Cranbury,NJ,USA),and epidermal growth factor (Sigma) and used to conduct the following experiments.
Transwell migration assay
A 24-well Transwell chamber (pore size,8 mm;Corning Incorporated,Corning,NY,USA) was used to evaluate the effect of metformin on SCMEC migration in the exponenti al growth phase.Trypsin was used to digest the SCMEC to create a cell suspension.Next,2 × 105cells/mL was added to the upper chamber,which contained DMEM (Gibco) supplemented with 1% fetal bovine serum,while medium containing 10% fetal bovine serum (600 µL) was introduced into the lower chamber.Thereafter,metformin at diverse doses (0.5,1,2,or 4 mM) and compound C (1 µM) were added into the bottom chamber.After incubation for 24 hours,the cells that had migrated to the bottom surface of the lower chamber were fixed in 4% PFA for 20–30 minutes,followed by 10 minutes of staining with 0.1% crystal violet.The vertically migrated SCMECs in five randomly chosen visual fields were quantified using Image-Pro Plus 7.0 soft ware.
Scratch migration assay and tube formation assay
SCMECs were cultivated in six-well plates.When the cells reached 90%confluency,a scratch was made in the monolayer using a 200-µL pipette ti p,and 200 µL metformin at different concentrations (0.5,1,2,and 4 mM) and compound C (1 µM) were added.At 0,6,and 12 hours following treatment,a light microscope (CKX41;Olympus Corporation,Tokyo,Japan) was used to obtain images.The wound width was measured as described previously (Ni et al.,2019).
Anin vitroangiogenesis assay tube formation kit (Cultrex,Trevigen,Inc.,Gaithersburg,MD,USA) was used to evaluate tube formation after 24 hours.Briefly,SCMECs (2 × 104/well) were inoculated into wells containing 100 µL Matrigel,followed by treatment with different concentrations of metformin.The wound closure length,total branching length,and number of mesh extensions were calculated using Image-Pro Plus 7.0 soft ware.
Western blot assay
Activation of the AMPK/eNOS pathway has been shown to have a beneficial effect on endothelial cells after metformin treatment.Spinal cord tissues were obtained at 7 days post-SCI and lysed using a radioimmunoprecipitation assay lysis buffer (Beyotime,Shanghai,China),and protein concentrations were measured using a bicinchoninic acid protein assay kit according to the manufacturer’s instructions (Bio-Rad,Hercules,CA,USA).Proteins were separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis,followed by transfer to a polyvinylidene fluoride membrane.The membrane was blocked with 5% milk in Tris-buffered saline with Tween.The membrane was then incubated overnight at room temperature (22–25°C) with anti bodies against AMPK (rabbit,1:500,Wanleibio,Shenyang,China,Cat# WL02254),phosphor-AMPK (p-AMPK;rabbit,1:500,Wanleibio,Cat# WL05103),eNOS(rabbit,1:500;Wanleibio,Cat# WL01789),and phosphor-eNOS (p-eNOS;rabbit,1:1000,CST,Beverly,MA,USA,Cat# 9571,RRID: AB_329837).Then,a secondary antibody (goat anti-rabbit IgG H&L (horseradish peroxidase),1:2000,Abcam,Cat# ab6721,RRID: AB_955447) was added,and the membrane was incubated for 1 hour at room temperature.Finally,enhanced chemiluminescence (Amersham Biosciences,Pittsburgh,PA,USA) was used for band visualization,and images were obtained using a gel documentation system (Bio-Rad Gel and Multi-Analyst version 1.1,Bio-Rad,Hercules,CA,USA).Image Lab 3.0 software (BioRad) was employed to quantify band intensities.
Quanti tative polymerase chain reaction
We choose four angiogenesis-related genes––angiopoietin-1 (ANG-1),brainderived neurotrophic factor (BDNF),connective tissue growth factor (CTGF),and vascular endothelial growth factor (VEGF)– to explore differences between the young and aged groups before and 7 days after SCI.Spinal cord samples (1 cm long) near the injury center were collected,and total RNA was extracted using TRIzol (Invitrogen,Carlsbad,CA,USA) according to the manufacturer’s instructions.Complementary DNA was then prepared by reverse transcription of the total RNA using a PrimeScript RT reagent kit (Takara,Kusatsu,Japan).A QuantStudioTM5 Real-time PCR system(Applied Biosystems,Carlsbad,CA,USA) was used to perform quantitative polymerase chain reaction (qPCR) using SYBR Premix Ex Taq (Takara) with the following conditions: 40 qPCR cycles and an annealing temperature of 60°C.Gene expression levels were determined by the 2–∆∆CTapproach (Livak and Schmittgen,2001),using glyceraldehyde phosphate dehydrogenase (GAPDH)as a housekeeping gene.The qPCR primer sequences are as follows: VEGF:forward,5′-CTG TCT AAT GCC CTG GAG CC-3′,and reverse,5′-ACG CGA GTC TGT GTT TTT GC-3′;ANG-1: forward,5′-CAG CAC GAA GGA TGC TGA TA-3′,and reverse,5′-TTA GAT TGG AAG GGC CAC AG-3′;BDNF: forward,5′-ATG ACC ATC CTT TTC CTT ACT-3′,and reverse,5′-CTA TCT TCC CCT TTT AAT GGT-3′;CTGF: forward,5′-GTC TTC ACA CTG GTG CAG CC-3′,and reverse,5′-ACT GGA AGA CAC ATT TGG CC-3′;GAPDH: forward,5′-AGA AAA ACC TGC CAA ATA TGA TGA C-3′,and reverse,5′-TGG GTG TCG CTG TTG AAG TC-3′.
Stati stical analysis
No statistical methods were applied to predetermine the sample size.However,more animals were used in this study than in a previous publication(Arifin and Zahiruddin,2017).No animals or data points were excluded from the analysis.The evaluators were blinded to the group assignments.GraphPad Prism 8.0.2 (GraphPad Software,San Diego,CA,USA,www.graphpad.com)was used to perform the statistical analyses.All the data were first tested for normality based on the Shapiro-Wilk test.All data are represented using means ± standard error of the mean (SEM).Two-tailed Student’st-test was adopted for comparison between two groups,whereas one-way analysis of variance followed by Tukey’s multiple comparisons test was performed to compare multiple groups.P<0.05 was considered to indicate statistical significance.
Results
Aging delays functional recovery and pathological outcomes in SCI
To assess how aging affects neurological recovery after SCI,three behavioral tests were implemented to analyze the differences between young and aged mice.As presented inFigure 1AandB,the BMS score and subscore were not significantly different between young and aged mice before SCI but trended toward recovery in both young and aged mice after SCI (Figure 1AandB).However,the aged SCI mice had markedly lower scores compared with young SCI mice starting at 7 days post-SCI.The aged group had a higher withdrawal threshold in response to mechanical stimuli post-SCI compared with the young group (Figure 1C).In addition,the aged group exhibited markedly impaired MEP amplitudes and latency periods 56 days post-SCI (Figure 1D–F).
Next,we examined the pathology of the aged spinal cord after SCI.First,we confirmed that the pathological changes 14 days post-SCI were consistent in the young and aged groups (Additional Figure 4) and found that the area of the lesion tissue around the injury site was larger in aged mice compared with young SCI mice at the same time point.Next,we explored the pathology at 56 days post-SCI.AsFigure 1G–Hshows,the area of the lesion around the injury site was larger in aged mice compared with young SCI mice at this time point and,conversely,there was less healthy spinal cord tissue in aged mice compared with young mice post-SCI (Figure 1I).Moreover,footprint analysis revealed that stride length and paw rotation were severely impaired in aged mice compared with young mice post-SCI (Figure 1J–L).Taken together,these findings imply that pathological changes in the spinal cord are worse,and neurological functional recovery is more impaired,in aged mice compared with young mice post-SCI.
Vascular regeneration is impaired in aged mice after SCI
To determine whether the poor recovery in aged mice post-SCI was due to impaired angiogenesis,the blood vessels were visualized by immunostaining.As presented inFigure 2AandB,the aged mice exhibited a lower percentage of newly formed blood vessels (CD31-positive cells) at 56 days post-SCI compared with young mice.Because the greatest degree of vascular proliferation occurs at 7 days post-injury (Cao et al.,2017),this time point was selected to evaluate the proliferation of SCMECs isolated from aged and young mice.AsFigure 2CandDshows,immuno-stained spinal cord slices taken 7 days post-SCI exhibited substantially fewer proliferated endothelial cells (Ki-67 and CD31 double-positive cells) in the aged group compared with the young group,suggesting that micro-vessel growth is attenuated in aged mice after SCI.Next,qPCR was conducted to analyze alterations in the mRNA expression levels of four angiogenesis-related genes (ANG-1,BDNF,CTGF,and VEGF) within injured spinal cord tissue from both groups.We found that ANG-1,BDNF,CTGF,and VEGF mRNA levels were markedly higher in the young group compared with the aged group at 7 days post-SCI (Figure 2E–H).Then,comparative X-ray microscope tomography imaging of the spinal cord microvasculature was performed to analyze specific 3D morphological alterations post-SCI in both young and aged mice.As shown inFigure 3A,visualization of the 3D architecture of the spinal cord micro-vessel networks enabled accurate quantification of vascular segment,vascular volume fractions,and vascular bifurcation.Figure 3Adepicts 3D images of the spinal cord microvasculature at the T10 levels in control and injured young and aged mice.In the control animals,the quanti tative parameters of the spinal cord vasculature were similar in aged mice and young mice.However,at 56 days post-SCI,the vascular parameters of the spinal cord were lower in aged mice than in young mice (Figure 3B–D),which was consistent with our immunostaining results.Collectively,these data suggest that the impaired neurological functional recovery observed in the aged mice may be associated with impaired vascular regeneration.
Metformin promotes SCMEC migration and tube formation in vitro
Metformin has anti-aging and pro-angiogenic effects in the context of several diseases (Takahashi et al.,2015;Barzilai et al.,2016;Yu et al.,2016;Piskovatska et al.,2020).To determine whether metformin promotes angiogenesis after SCI,we first explored its effects on SCMECsin vitro.First we isolated SCMECs and confirmed their identity by light microscopy and immunofluorescence (Additional Figure 5).Seven days after seeding,SCMEC colonies appeared (Additional Figure 5A),and CD31 immunofluorescence was detected (Additional Figure 5B).Next,we performed scratch and Transwell assays and found that metformin enhanced SCMEC migration horizontally and vertically in a concentration-dependent manner,as shown inFigure 4A–D.Additionally,SCMEC canaliculization increased markedly as the metformin concentration increased,as indicated by the tube formation assay (Figure 4E–G).Of note,4 mM metformin induced less SCMEC migration and tube formation than 2 mM metformin,suggesting that 2 mM may be the optimal concentration of metformin for promoting SCMEC-mediated angiogenesis.
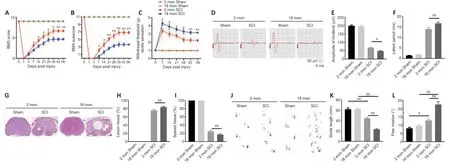
Figure 1|Aging attenuates neurological functional recovery and tissue repair after SCI.
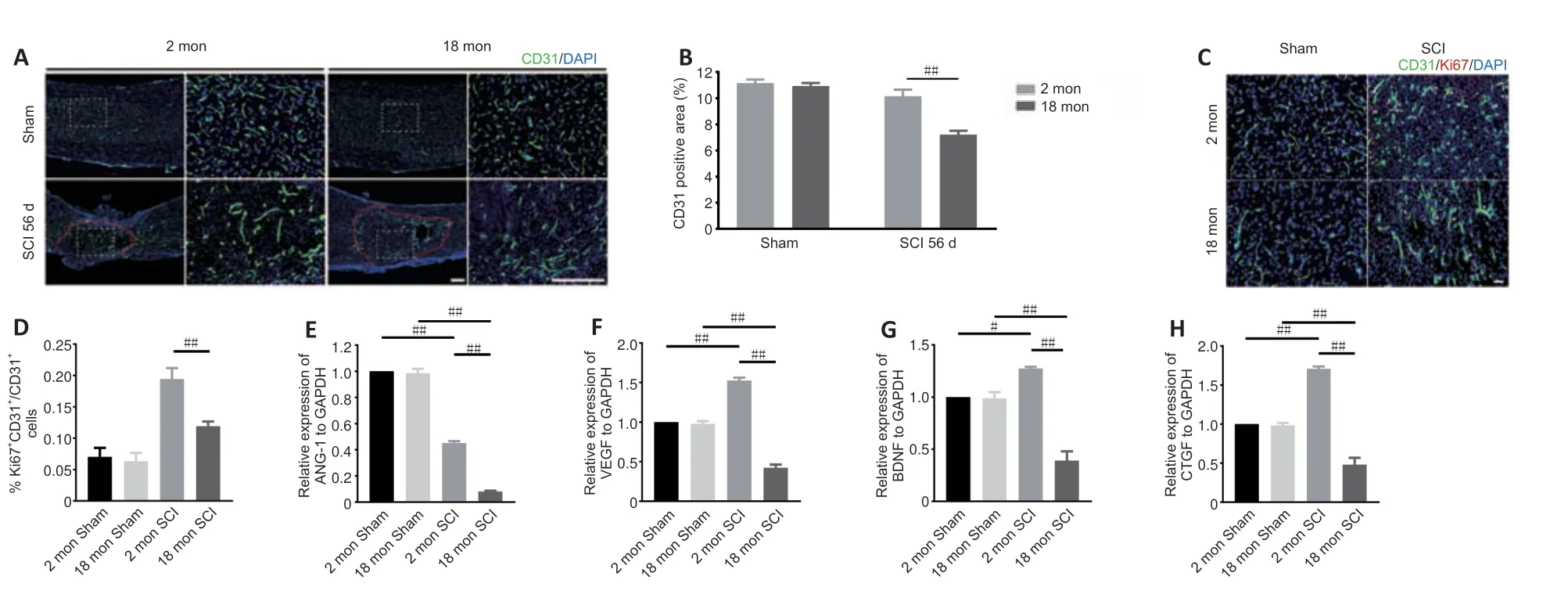
Figure 2|Aging results in reduced vessel numbers and vascular proliferation after SCI.
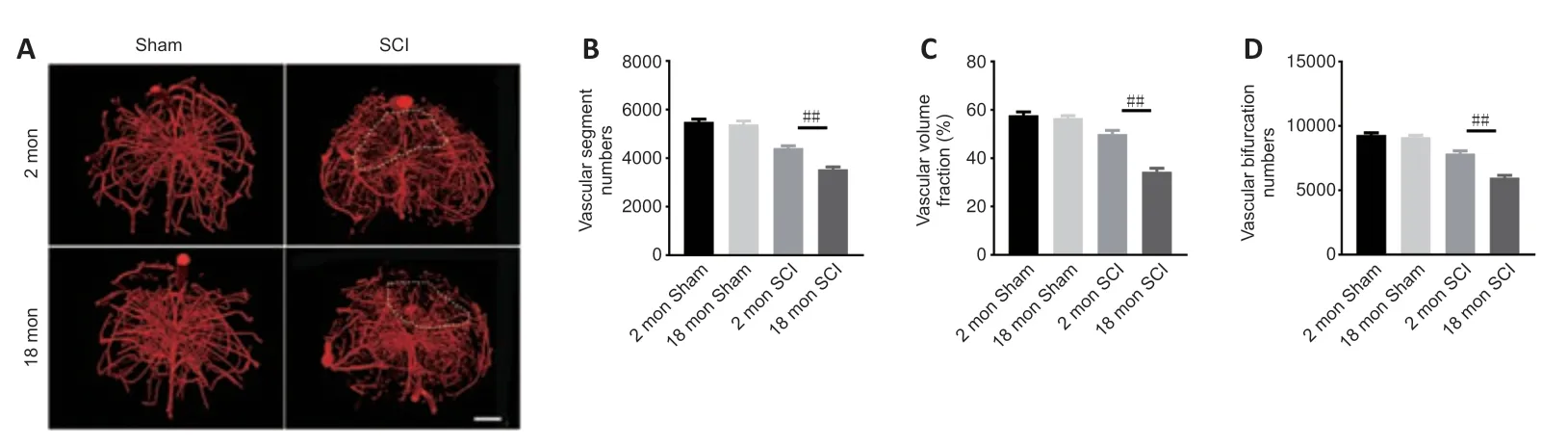
Figure 3|Three-dimensional visualization of microvasculature changes in the spinal cords of young and aged mice post-SCI.
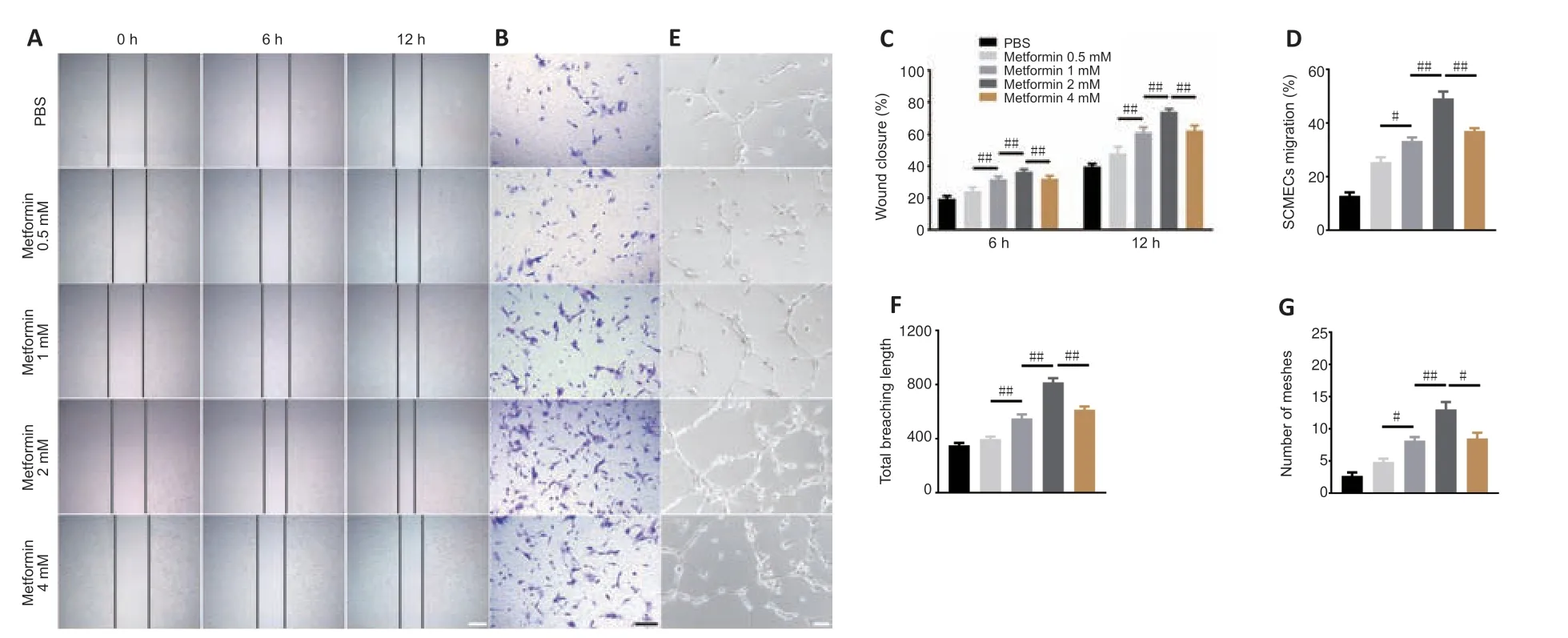
Figure 4|Metformin promotes SCMEC-mediated angiogenesis in vitro.
Metformin promotes neurological functional recovery and tissue repair in aged mice after SCI
To better illustrate metformin’s role in tissue repair and functional recovery in aged mice after SCI,we systematically evaluated the recovery of hindlimb motor function.First,the BMS score and subscore were determined to assess the recovery of neurological function in the metformin and PBS groups.As shown inFigure 5AandB,the aged mice treated with metformin had markedly higher BMS scores and subscores (starting at 7 days post-SCI) and enhanced motor function than the aged mice treated with PBS post-SCI.Additionally,the aged mice treated with metformin had lower withdrawal thresholds in response to mechanical stimulus than the aged mice treated with PBS post-SCI,suggesting that metformin may promote sensory recovery in aged mice post-SCI (Figure 5C).To better evaluate the role of metformin function in the recovery of motor function,electrophysiological testing was carried out.As shown inFigure 5D,treatment with metformin lead to clearly elevated MEP amplitudes and markedly shorter latency periods relative to treatment with PBS at 56 days post-injury (Figure 5EandF).H&E staining showed that the area of spinal cord tissue with normal integrity at the injury site was markedly increased in the metformin group compared with the PBS group (Figure 5GandH),and the area of injured tissue was dramatically decreased in the metformin group compared with PBS group (Figure 5I).Furthermore,footprint analysis showed that treatment with metformin resulted in a longer stride length and less paw rotation in aged mice at 56-day post-SCI relative to the PBS group (Figure 5J–L).These results suggest that metformin is neuroprotective and significantly improves tissue repair and neurological functional recovery in aged mice post-SCI.
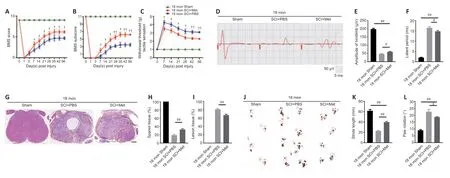
Figure 5|Metformin promotes neurological functional recovery and tissue repair after SCI in aged mice.
Metformin promotes angiogenesis in aged mice after SCI
Next,we asked whether the neuroprotective effect of metformin on SCI is due to its promotion of angiogenesis.Immunofluorescence analysis showed that metformin administration markedly increased the percentage of CD31-positive cells at the injury site in aged mice 7,14,and 56 days after SCI compared with PBS administration (Figure 6AandB).Furthermore,treatment with metformin increased the percentage of CD31 and Ki67 double-positive cells in aged mice at 7 days post-SCI (Figure 6CandD).
To explore the molecular mechanism by which metformin promotes angiogenesis in aged mice post-SCI,we investigated the AMPK/eNOS pathway,which has been shown to have a beneficial effect on endothelial cells after treatment with metformin (Takahashi et al.,2015;Yu et al.,2016).The AMPK/eNOS pathway was markedly activated in the spinal cord of aged animals 7 days post-SCI after treatment with metformin,as confirmed by western blotting analysis (Figure 6E–G).Collectively,these results suggest that activation of the AMPK/eNOS pathway may be involved in the metforminmediated angiogenesis observed in aged mice post-SCI.
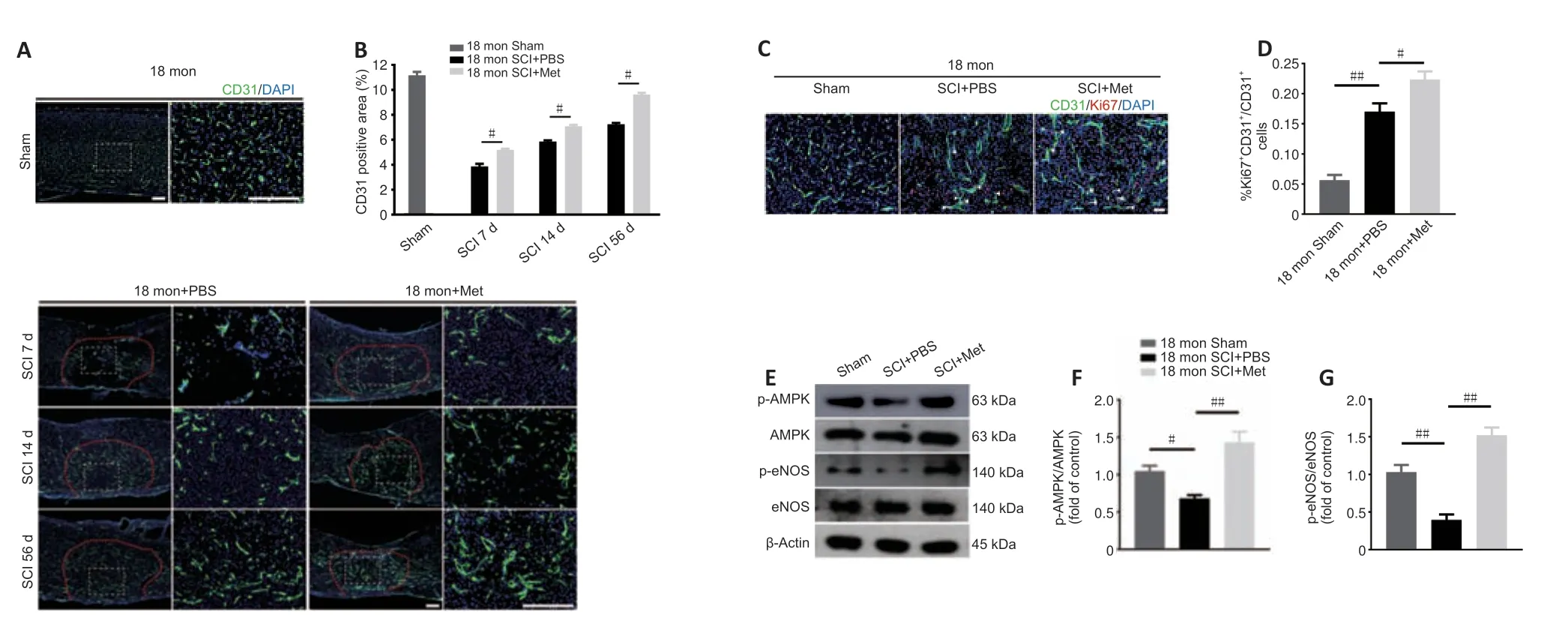
Figure 6|Metformin promotes vascular regeneration in aged mice post-SCI.
Inhibition of the AMPK/eNOS pathway reverses the pro-angiogenic effects of metformin on SCMECs
To determine whether the AMPK/eNOS pathway affects SCMEC function after treatment with metformin,AMPK expression was inhibitedin vitrousing compound C.First,we confirmed by western blotting that activation of the AMPK/eNOS pathway was blocked by treatment with compound C(Figure 7A–C).Then,we performed scratch and Transwell assays and found that compound C abolished the horizontal and vertical SCMEC migration caused by treatment with 2 mM metformin (Figure 7D–G).Compound C also robustly repressed SCMEC canaliculization,as determined by quantification of tube formation (Figure 7H–J).These findings indicate that inhibition of the AMPK/eNOS pathway attenuated the pro-angiogenic effects of metformin on SCMECsin vitro.
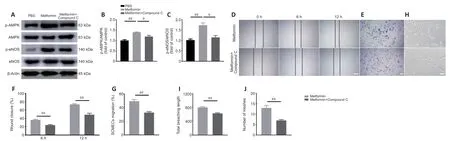
Figure 7|Inhibition of the AMPK/eNOS pathway attenuates the beneficial effects of metformin on angiogenesis in SCMECs in vitro.
Inhibition of the AMPK/eNOS pathway abolishes the beneficial effect of metformin on functional recovery and angiogenesis in aged SCI mice
As described above,the AMPK inhibitor compound C attenuates the ability of metformin to promote SCMEC migration and tube formationin vitro.To explore the effect of compound C on neurological functional recovery and angiogenesis,next we intraperitoneally injected compound C and metformin into aged mice after SCI.We first asked whether compound C affects the tissue architecture of the spinal cord before SCI and 56 days post-SCI.As shown inAdditional Figure 6,treatment with compound C did not affect the tissue architecture of spinal cord before or after SCI in aged mice.Next,we conducted behavioral assessments (the BMS scoring system and Von-Frey test) over an 8-week observation period to evaluate motor and sensory functional recovery.As shown inFigure 8AandB,BMS scores and subscores were clearly lower in the metformin plus compound C treatment groups than in the metformin only groups beginning at 7 days post-SCI (Figure 8AandB).The withdrawal thresholds in response to mechanical stimuli were also increased in aged mice treated with metformin plus compound C relative to those treated with metformin only,suggesting that compound C abolished the beneficial effect of metformin on motor and sensory functional recovery in aged mice post-SCI (Figure 8C).Electrophysiological analysis showed that,in aged mice post-SCI,the combined treatment group had markedly worsened MEP amplitudes and latency periods compared with metformin only group (Figure 8D–F).In addition,histological analysis revealed less preservation of spinal cord integrity following combined treatment compared with treatment with metformin only (Figure 8GandH).The opposite was true for the lesioned area: the injured area of the spinal cord following treatment with metformin plus compound C was markedly increased compared with metformin only (Figure 8I).Furthermore,footprint analysis demonstrated that the aged mice treated with metformin had a much longer stride length and less paw rotation at 56 days post-SCI,while treatment with metformin plus compound C inhibited hindlimb functional recovery (Figure 8J–L).Thus,compound C inhibits the ability of metformin to promote neurological functional recovery post-SCI in aged mice.
To further explore the role of the AMPK/eNOS pathway in metforminpromoted angiogenesisin vivo,we used immunofluorescence to detect vascular formation and endothelial cell proliferation following treatment with metformin plus compound C.Both the vascular ratio and endothelial cell proliferation at the injury site were reduced after treatment with compound C compared with treatment with metformin alone in aged mice after SCI(Figure 9A–D).These results suggest that compound C inhibits the ability of metformin to promote angiogenesis and neurological functional recovery in aged mice post-SCI.
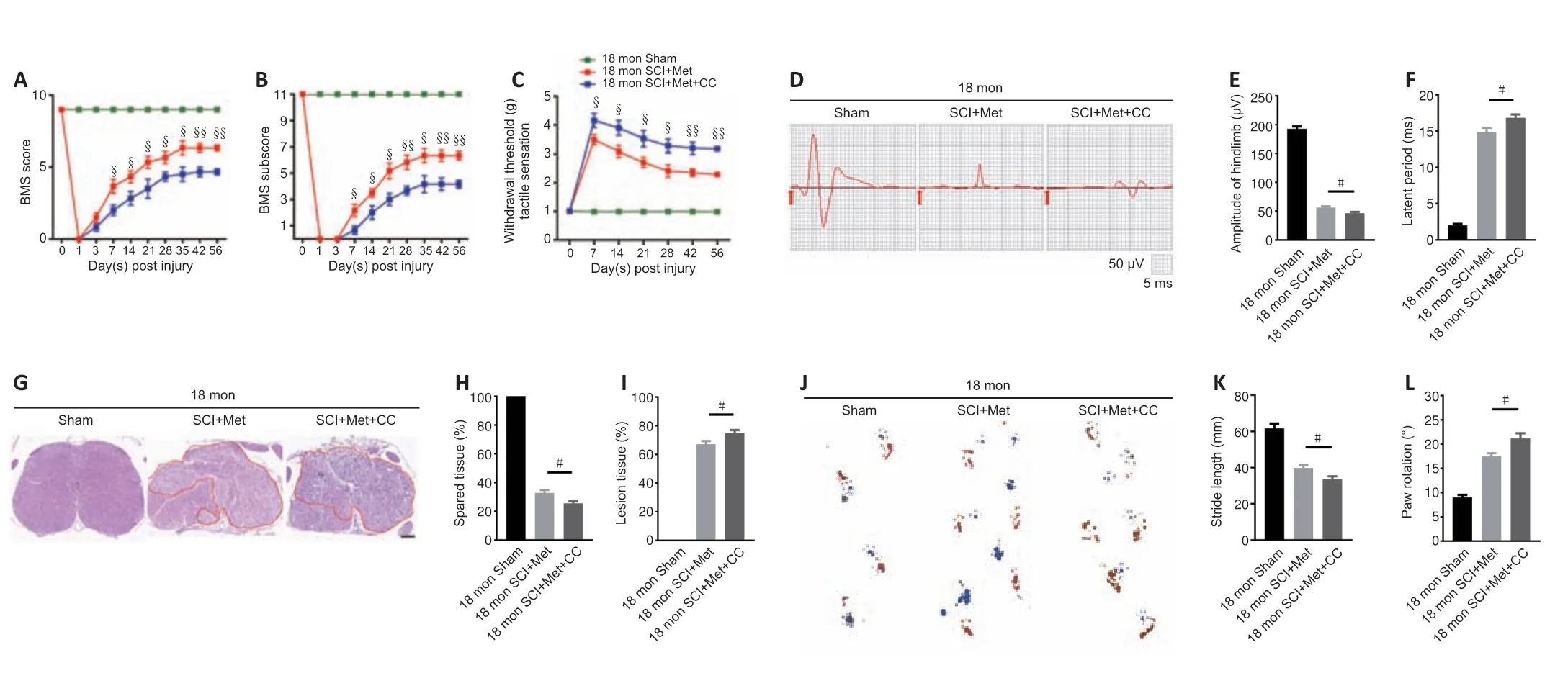
Figure 8|Inhibition of the AMPK/eNOS pathway abolishes the beneficial effects of metformin on functional recovery in the aged mice following injury.
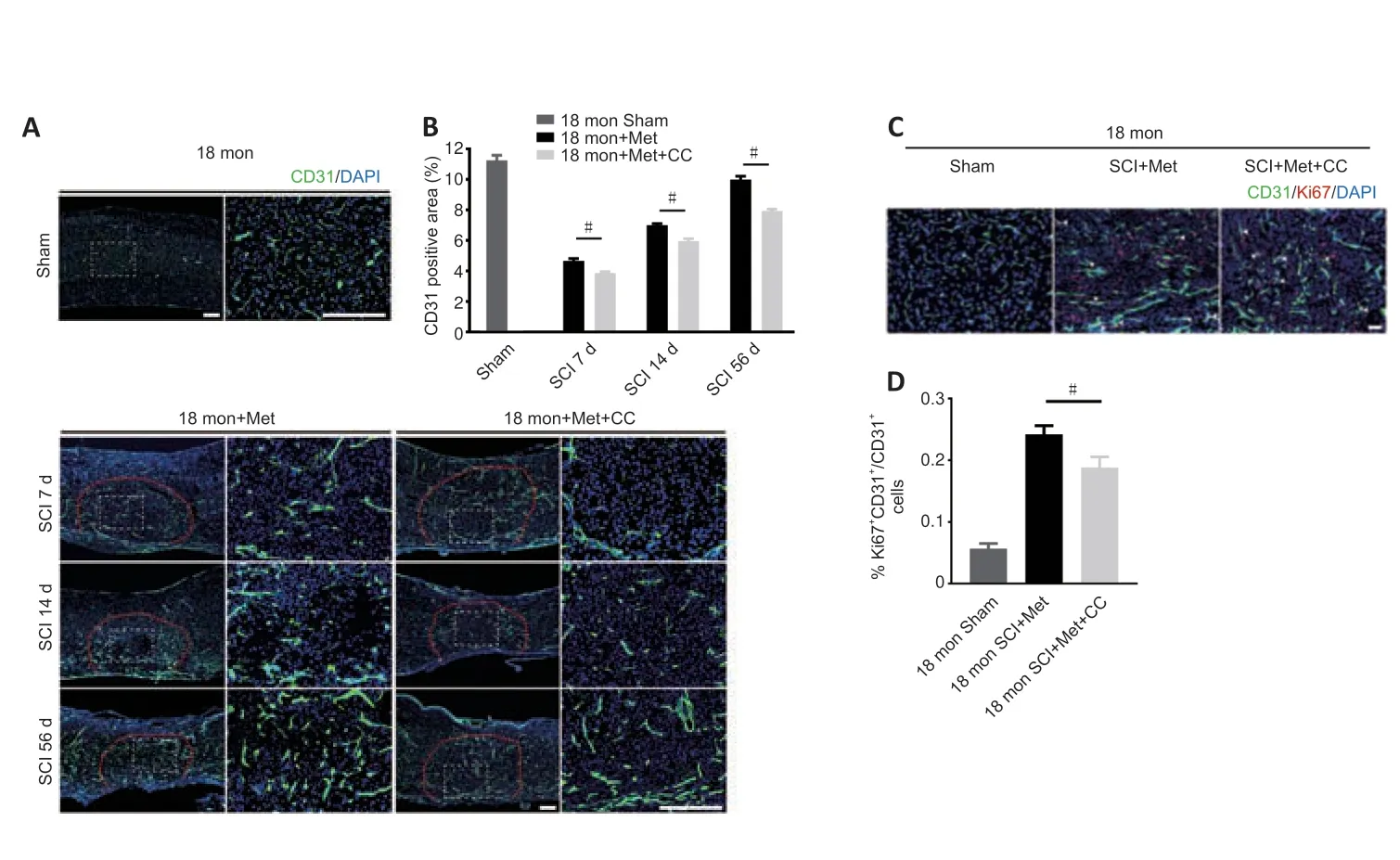
Figure 9|Inhibition of the AMPK/eNOS pathway reverses the beneficial effects of metformin on angiogenesis in aged mice following injury.
Discussion
The mammalian central nervous system has very low capacity for regeneration due to its limited regenerative plasticity (Hilton and Bradke,2017;Tran et al.,2018).Neovascularization is therefore of paramount importance for the neuroparenchyma tissue survival (Figley et al.,2014).In the healthy spinal cord,vasculature provides nutrients and removes metabolic waste,and these processes play an important role in maintaining normal function.Therefore,promoting angiogenesis post-SCI is a promising therapeutic goal.
The mechanical forces involved in SCI,however,result in destruction of the vasculature,causing local hypoxia at the lesion epicenter.Angiogenesis initi ates a few days post-SCI in response to hypoxia but regresses significantly around 14 days post-injury;thus,it cannot provide sufficient nutrition for SCI repair (Benton et al.,2008;Cao et al.,2017).All organs experience dynamic changes with aging (DiLoreto and Murphy,2015;Ungvari et al.,2018),and recovery after SCI requires blood vessel growth.Nevertheless,it is unknown whether aging impacts angiogenesis after SCI.
Previous research demonstrated that ischemia-induced angiogenesis was attenuated in aged rats subjected to middle cerebral artery occlusion(Tang et al.,2016).In the current study,we found that the morphological parameters of the spinal cord vascular network differed in aged versus young mice subjected to SCI.Furthermore,we found that,while traumainduced angiogenesis occurs in both young and aged mice after SCI,vessel regeneration is less robust in aged mice,which is consistent with findings from a previous study (Tang et al.,2016).
The functionality of the nervous system is dependent upon a normal blood supply (Figley et al.,2014),and therefore increased blood-vessel density is closely associated with improved functional outcomes after SCI.According to our results,in addition to reducing the angiogenic response in mice,aging limited the recovery of locomotive and sensory functions post-SCI.Therefore,our findings suggest that angiogenesis may influence nerve function recovery post-SCI.
Trauma-induced angiogenesis is regulated via complicated interactions among diverse molecules (Sinescu et al.,2010;Shi et al.,2017,2018).Previous studies have characterized the many cellular events involved in angiogenesis after SCI.Importantly,remodeling of the functional vascular network is dependent on factors secreted by the local microenvironment after injury (Graumann et al.,2011;Kundi et al.,2013).Our observations revealed that angiogenesis is impaired in the aged spinal cord in response to trauma,although the molecular pathways that are reactivated under these pathological conditions and the effect of aging on angiogenesis at the cellular and subcellular levels remain largely unclear.
Ang-1,which is a main member of the angiopoietin family,promotes endothelial cell survival,sprouting,and tube formation,which are involved in both physiological and pathological angiogenesis processes (Isidori et al.,2016;Duran et al.,2021;Joussen et al.,2021).Unlike numerous other vascular growth factors,the expression of BDNF,which is a newly described meditator of endothelial cell survival and angiogenesis,is relatively restricted to central arteries (Donovan et al.,2000;Kermani and Hempstead,2007;Wang et al.,2019).In addition to VEGF,CTGF plays a critical role in vessel homeostasis and angiogenesis (Brigstock,2002;Ungvari et al.,2017).In our study,we found that these four angiogenesis-related genes (ANG-1,BDNF,CTGF,and VEGF) were upregulated after injury to the spinal cord tissue.However,it appears likely that this response may be impaired by aging,which could help explain the impaired vascular function and angiogenesis observed following injury in the elderly (Dumbrava et al.,2022;Riggin et al.,2022).It will be of great interest to elucidate exactly how these angiogenic factors are affected by aging process.Novel therapies that promote secretion of these factors into the local microenvironment may be conducive to regenerating blood vessels in elderly patients.
In addition to changes in local molecular factors that occur with aging,we speculate that other aging-related physiological changes in the vasculature could also affect vascular regeneration and neuronal integrity.Loss of stem cell acti vity with aging is likely to disrupt the regeneration process.Previous research has identified a resident vascular progenitor cell that localizes around blood vessels and could be capable of regenerating capillary networks(Pang et al.,2017).However,aging could impair the function of endothelial progenitor cells and inhibit their differentiation (Ungvari et al.,2018).In addition,endothelial cells undergo cellular senescence with aging,ceasing to proliferate and thus limiting the capacity to form new vasculature (Lähteenvuo and Rosenzweig,2012;Ungvari et al.,2018).Our findings are consistent with those from a previous study (Tang et al.,2016).Future research may help define the underlying mechanisms of aging that impact angiogenesis.In addition,gaining a better understanding of the pathological changes that occur in the vasculature of aged mice after injury will help develop targeted therapeutic interventions for promoting angiogenesis in elderly patients.Pharmacological approaches that eliminate senescent endothelial cells could aid vessel regeneration and enhance the recovery of neurological function post-SCI in aged rodents.
Over the past decade,substantial progress has been made in treating SCI,although only a small number of treatment strategies have been successfully translated to the clinic,and these treatments only slightly improve functional recovery,especially among aged SCI patients.Given the aging of the global population,anti-SCI interventions for elderly patients are urgently needed.Over the past several years,increasing evidence has shown that metformin,a common hypoglycemic drug,has pleiotropic effects on several agingrelated diseases and biological functions associated with aging (Chen et al.,2016;Piskovatska et al.,2020) and reduced risk of cancer (Kamarudin et al.,2019).Treatment with metformin may be a promising approach to achieving functional recovery post-SCI because it promotes improvement of a variety of biological functions.In several preclinical studies of SCI,metformin was shown to be capable of regulating autophagy and repressing inflammation and neuronal apoptosis,thus improving functional recovery after SCI,potenti ally through activation of the Wnt/β-catenin pathway (Wang et al.,2016;Zhang et al.,2017b,2020).In addition,the administration of metformin enhances axon regeneration by reducing oxidative stress and stabilizing microtubules (Wang et al.,2020),indicating that it has substantial regenerative potential that could be helpful for the treatment of SCI.Moreover,metformin significantly prolongs longevity in mice by enhancing their overall health (Anisimov et al.,2010;Cabreiro et al.,2013).These effects not only emphasize the potenti al importance of metformin treatment in aging and in SCI treatment but also could open up new avenues for therapeutic developments in treating aged mice after SCI.In the current study,we found that metformin treatment significantly promotes vascular regeneration in aged mice after SCI,thereby enhancing tissue repair and recovery of hindlimb motor and sensory function.This finding highlights the role of metformin in regulating blood vessel formation,which enhance sensorimotor recovery and neuroplasticity.
AMPK,which is an important enzyme responsible for energy metabolism,is activated by metformin (Zhou et al.,2001;Musi et al.,2002).AMPK phosphorylates a number of enzymes,including acetyl-coenzyme A carboxylase,3-hydroxy-3-methylglutaryl-coenzyme A reductase,glycogen synthase,and eNOS,which are associated with multiple biosynthetic reactions(Chen et al.,1999;Kahn et al.,2005;Long and Zierath,2006).A previous report demonstrated that AMPK induces NO production by phosphorylating eNOS,thereby maintaining a functional balance in endothelial cells (Yuyun et al.,2018).In addition,activation of the AMPK/eNOS pathway is closely associated with vascular endothelial cell regeneration,particularly under hypoxic and ischemic conditions (Ikeda et al.,2012;Huang et al.,2015;Xu et al.,2015b;Ren and Luo,2021).Metformin has also been reported to possibly activate the AMPK/eNOS pathway,thereby stimulating angiogenesis by endothelial progenitor cells in mice with diabetes (Yu et al.,2016) and promoting revascularization in an ischemic hindlimb mice model (Takahashi et al.,2015).These findings demonstrate the importance of metformin in angiogenesis,which is consistent with the current study,in which we found that activation of the AMPK/eNOS pathway may mediate metformininduced angiogenesis in aged mice post-SCI.Administration of compound C abrogated the beneficial effects of metformin on angiogenesisin vivoandin vitro,indicating that metformin treatment promotes neurological functional recovery and angiogenesis in aged mice after SCI through the AMPK/eNOS pathway.
This study had several limitations.First,18-month-old mice were used for the aged group,while,considering the demographic shift in patients with SCI,older mice (such as 22–24 months of age) may be more appropriate to explore angiogenesis and hindlimb motor recovery post-SCI.Second,we only explored the effect of metformin on the AMPK/eNOS pathway;future work should investigate other pathways activated by metformin administration.Third,noninvasive imaging technologies for rendering 3D images of the spinal cord vasculature,especially those that can be translated to the clinic,should be used in future studies.
Taken together,our findings show that functional recovery and blood vessel formation are decreased in aged mice compared with young mice post-SCI.Metformin played a neuroprotective role and enhanced neurological functional recovery after SCI in aged mice,possibly by promoting angiogenesis.Furthermore,the ability of metformin to promote angiogenesis and neurological functional recovery after SCI may be mediated by activation of the AMPK/eNOS pathway.Moreover,our study provides a novel therapeutic approach for promoting angiogenesis and functional recovery after SCI in aged mice.
Acknowledgments:We would like to thank Prof.Hui Xie and other staff from the Movement System Injury and Repair Research Center,Xiangya Hospital,Central South University,Changsha,China;and Prof.Xiang-Hang Luo and other staff from the Department of Endocrinology,Endocrinology Research Center,Xiangya Hospital of Central South University,Changsha,China,for their kind assistance during the experiments.
Author contributions:Study design and supervision: CYD,JZH;experimentimplementation: JYZ;data analysis: XLS,CJL,TQ,RDH,GYD;manuscript draft :JYZ,YC;manuscript revision: HBL,CYD,JZH.All authors contributed to and approved the final version of manuscript.
Conflicts of interest:The authors have no conflicts of interest relevant to this article.
Availability of data and materials:All data generated or analyzed during this study are included in this published article and its supplementary information files.
Open access statement:This is an open access journal,andarticles are distributed under the terms of the Creative Commons AttributionNonCommercial-ShareAlike 4.0 License,which allows others to remix,tweak,and build upon the work non-commercially,as long as appropriate credit is given and the new creations are licensed under the identical terms.
Open peer reviewer:Úrzula Franco-Enzástiga,University of Texas at Dallas,USA.
Additional files:
Additional Figure 1:Schematic of the experiment to explore the difference between young and aged mice.
Additional Figure 2:Schematic of the experiment to explore the effect of metformin treatment on aged SCI mice.
Additional Figure 3:Schematic of the experiment to explore the effect of Compound C on the aged SCI mice with metformin treatment.
Additional Figure 4:Aging exacerbates histological tissue repair at 14 days post-SCI.
Additional Figure 5:Identification of SCMECs.
Additional Figure 6:Compound C did not change the histology tissue architecture of spinal cord at 56 days post-SCI among different groups.
Additional file 1:Open peer review report 1.

- 中国神经再生研究(英文版)的其它文章
- Bystanders or not? Microglia and lymphocytes in aging and stroke
- Alzheimer’s disease risk after COVID-19: a view from the perspective of the infecti ous hypothesis of neurodegeneration
- Serine and arginine rich splicing factor 1: a potenti al target for neuroprotection and other diseases
- Can glial cells save neurons in epilepsy?
- Lights for epilepsy: can photobiomodulation reduce seizures and offer neuroprotection?
- The landscape of cognitive impairment in superoxide dismutase 1-amyotrophic lateral sclerosis