
疥螨Sar s 14.3过敏原蛋白原核表达及间接ELISA方法的建立
2023-02-20 08:17唐志强张浩吉唐文强党志胜孙悦翔叶碧锦黄福强
中国人兽共患病学报 2023年1期
李 鑫,唐志强,张浩吉,唐文强,党志胜,孙悦翔,李 静,叶碧锦,黄福强
疥螨病是由疥螨科、疥螨属的疥螨(Sarcoptesscabiei)寄生于人和其它动物表皮内所引起的一种接触性、传染性皮肤病[1-2]。严重感染的疥螨病由于痂皮及皮下存在大量的疥螨,病原学诊断能较易检测到。但在普通感染的疥螨病例中,由于皮内仅有少量的疥螨,且缺乏明显的症状,因此病原学检测较为困难,往往出现假阴性结果[1]。疥螨病免疫诊断多釆用疥螨粗提取物作为诊断抗原,但由于疥螨虫体小且缺少体外培养系统,很难短时间内获得足够疥螨粗提取物用于疥螨病的免疫诊断[1]。因此,探寻特异性高、免疫性强的诊断抗原,开发简单、高效的诊断方法显得非常必要。
随着高通量测序技术和相关组学研究的发展,越来越多物种的基因组和转录组学信息被揭示。Rider SD等(2015)[3]和Korhonen PK等(2020)[4]分别发表了关于疥螨犬变种和疥螨猪变种基因组草图及相关组学的研究。疥螨基因草图的公布为新的诊断标志物和疫苗研发奠定了重要基础。应用基因组、蛋白质组和转录组等生物信息学方法筛选靶标分子,然后结合后续试验验证靶标分子的有效性已经成为挖掘诊断靶标分子及寄生虫与宿主互作的一种有效策略[5]。在疥螨病免疫诊断的研究中发现,一些过敏原分子具有较好的诊断价值[6-9]。为了找出疥螨猪变种过敏原相关分子,本试验对所采集疥螨用转录组测序并从头组装转录本,然后借助COMPARE过敏原数据库(https://comparedatabase.org/)再结合BLASTx程序,预测组装的疥螨转录组中过敏原数量和种类。最后,为了今后开展疥螨病诊断和流行病学调查,试验还用系列生物信息分析软件对国外已报道的有较好早期诊断价值的疥螨过敏原Sar s 14.3蛋白[1,9]进一步进行了生物信息学分析、原核表达及纯化,并利用该过敏原蛋白成功构建了间接ELISA诊断方法。
1 材料与方法
1.1 疥螨猪变种转录组测序 在广东某规模化猪场里通过巡栏找到感染疥螨的猪(图 1)。用勺子刮取病患交界处的皮肤组织,将采到的组织样品挑取一小块放入到干净的大平皿中,再加入适量的超纯水,混匀后在解剖显微镜下用挑虫针将疥螨一个一个挑出装入PCR管中,挑取50只,然后用TransZol UP Plus RNA Kit提取总RNA。将所提取的总RNA送往上海派森诺生物工程有限公司进行RNA-Seq测序及数据的处理。同时对采集的感染疥螨的猪皮屑按Omega Bio-tek Stool DNA Kit试剂盒所推荐的方法对皮屑样品进行DNA进行提取,以鉴定物种,物种鉴定结果参见文献[10]。

A:解剖镜下雌性疥螨(400×);B:严重感染疥螨的猪耳图1 疥螨猪变种及其引起的病变Fig.1 Sarcoptes scabiei and its lesions
1.2 疥螨猪变种过敏原目的基因的检索 从COMPARE过敏原数据库(https://comparedatabase.org/)中下载过敏原蛋白质序列及注释文件,然后以转录组测序组装的转录本数据为比对数据库,以COMPARE过敏原数据库中的过敏原蛋白序列为目标数据库,对本试验组装的转录本进行过敏原蛋白注释(Cutoff:E<1E-20,Identity>30%)。
1.3 疥螨Sar s 14.3蛋白生物学特性预测
1.3.1 Sar s 14.3过敏原理化性质预测 利用Expasy提供的在线工具ProtParam (http://web.expasy.org/protparam/)分析疥螨Sar s 14.3蛋白分子质量、理论等电点、氨基酸组成、原子组成、不稳定系数和总平均疏水性等理化性质。同时应用Phobius(https://phobius.sbc.su.se/)给定的默认参数对疥螨Sar s 14.3蛋白序列进行信号肽和跨膜结构域预测。
1.3.2 疥螨Sar s 14.3蛋白二、三级结构分析 应用SOMPA在线分析工具(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html )对疥螨Sar s 14.3 二级结构进行预测、分析,使用I-TASSER(https://zhanglab.dcmb.med.umich.edu/I-TASSER/)工具对疥螨Sar s 14.3 三级结构进行预测、分析。
1.3.3 Sar s 14.3蛋白多序列的对比 将本次测序并从头组装的疥螨猪变种Sar s 14.3核酸序列与NCBI nr数据库中所有蛋白进行BLASTx比对,找出其同源序列。
1.4 疥螨Sar s 14.3重组蛋白表达、纯化和鉴定 根据疥螨Sar s 14.3基因分析结果及原核表达宿主菌的密码子优化原则,由卡梅德生物科技(天津)有限公司合成并添加NdeI、XhoⅠ酶切位点,完成目的片段的合成。然后将其克隆到pET28a(+)表达载体上,构建成功的重组表达载体命名为pET28a-Sar,并将构建的pET28a-Sar转化至Rosetta(DE3)感受态细胞,涂板后37 ℃倒置培养过夜,再选取单克隆于5管LB培养基37 ℃培养至菌体OD600为0.6~0.8,加入IPTG至终浓度为0.5 mmol/L,培养4 h后离心,收菌制样。向接种保种菌培养至菌体OD600为0.6~0.8后的4管培养物中加入IPTG至终浓度分别为0.2 mmol/L,分别在37 ℃和15 ℃下220 r/min培养4 h和16 h,诱导融合蛋白表达,各条件制样SDS-PAGE分析最优表达条件,最后取上一步各条件菌液离心收集菌体,破碎(PBS buffer,pH7.4),上清、沉淀(用变性物将沉淀溶解制成“上清”样品)分别制样,SDS-PAGE进行可溶性分析。采用Ni柱亲和层析法对重组蛋白进行纯化,纯化后分别将流传液、裂解液、洗涤液和洗脱液进行SDS-PAGE电泳检测,纯化步骤按照蛋白纯化试剂盒(碧云天)说明书。对表达纯化后的Sar s 14.3 重组蛋白进行Western blot验证,将疥螨兔阳性血清(1∶200)作为一抗孵育,山羊抗兔 HRP-IgG(1∶1 000)作为二抗孵育,对其反应原性进行鉴定。
1.5 疥螨Sar s 14.3重组蛋白间接ELISA方法的建立
1.5.1 血清 建立方法所用标准血清:疥螨猪阳性血清,为采自经病原学和PCR检测呈阳性的猪;疥螨猪阴性血清,采自无疥螨感染史的猪场且无任何临床症状的猪。
1.5.2 棋盘法确定包被抗原浓度与血清的最佳倍工作浓度 用包被液将重组蛋白进行稀释,终浓度分别为0.5 μg/mL、1 μg/mL、2 μg/mL、3 μg/mL,每个孔加入100 μL稀释后的蛋白,4 ℃过夜包被,重复3组;PBST洗涤5次,每次5 min;使用5%脱脂奶粉对包被板进行封闭处理,37 ℃ 2 h;甩去封闭液,PBST洗涤。
标准阳性血清与阴性血清按照1∶100、1∶200、1∶400和1∶800稀释后,加入包被板中,每孔100 μL,每个稀释度重复3次,37 ℃孵育1 h;PBST洗涤,方法同上;将HRP标记兔抗猪IgG按1∶2 000稀释,每孔100 μL,37 ℃孵育30 min。避光环境下每孔加入100 μL的TMB显色液,室温孵育15 min后,每孔加入50 μL的终止液,用酶标液测定OD450值,并对结果进行分析,确定重组蛋白作为包被抗原时,最佳的抗原和血清的工作浓度。
1.5.3 包被时间、封闭液、封闭时间、孵育时间及二抗稀释倍数的确定 将抗原用最适包被浓度包被至酶标板中,分别在37 ℃ 1 h后4 ℃过夜、37 ℃ 2 h后4 ℃过夜、4 ℃过夜、37 ℃ 2 h的条件对抗原最佳包被时间进行优化,其余操作与1.5.2一致。最后通过OD450值确定最佳包被条件。为确定不同种类的封闭液和封闭时间是否会影响OD450值,选择5%脱脂奶粉、5% BSA、抗体稀释液在37 ℃条件下各孵育1 h、1.5 h、2 h,每个条件重复3次,封闭液加入的剂量都为200 μL/孔。以P/N比的大小决定最佳封闭时间和封闭液。用最佳包被浓度及包被时间将抗原包被至酶标板,在血清以最佳稀释浓度稀释好后,于37 ℃恒温培养箱内分别孵育0.5 h、1 h、1.5 h、2 h;使用PBST洗涤后;将HRP标记兔抗猪IgG以1∶2 000、1∶4 000、1∶6 000、1∶8 000的比例进行稀释,分别加到不同血清孵育时间后的孔内,37 ℃孵育0.5 h。


2 结 果
2.1 疥螨猪变种参考转录本组装 通过测序并拼接可获得转录本序列。提取每个基因下最长的转录本作为该基因的代表转录本即获得疥螨猪变种的参考转录组,参考转录组信息见表1。

表1 疥螨猪变种参考转录组信息Tab.1 Overview of reference transcriptome information for Sarcoptes scabiei var. suis
2.2 疥螨猪变种转录本过敏原预测 用本次测序所构建参考转录组中18 980个转录本检索COMPARE过敏原数据库中的蛋白序列共获得390个过敏原候选分子(Table S1),其中有28个基因序列的E值(随机匹配可能性)为0(Table S1),本实验组装的转录本(TRINITY_DN12024_c0_g1)按照编码蛋白翻译出的Sar s 14.3过敏原蛋白氨基酸序列与已发表的人源(Q8I9R5/AAO15613)、猪源(R9RIP2/AGM48615)疥螨第14组分过敏原(Group 14 allergen)蛋白部分序列、犬源(A0A132AJ56/KPM11048)和猪源(A0A834REG9/KAF7495137)疥螨第14组分过敏原蛋白序列比对部分相似性为100%(图2),提示疥螨过敏原Sar s 14.3氨基酸序列在不同宿主来源疥螨比较保守,能检测不同宿主来源疥螨血清。

图2 不同宿主来源疥螨第14组分过敏原蛋白(Sar s 14)C端氨基酸序列比对TRINITY_DN12024_c0_g1为本实验拼接并翻译的氨基酸序列Fig.2 C-terminal amino acid sequence alignment of group 14 allergen (Sar s 14) from different hosts of Sarcoptes scabiei TRINITY_DN12024_c0_g1 was the spliced and translated amino acid sequence in this experiment
2.3 Sar s 14.3蛋白理化性质 疥螨Sar s 14.3蛋白由330个氨基酸构成,理论分子质量约为38 kDa;理论pI值为8.59;其中含有57个强碱性(+)氨基酸(K,R)和54个强酸性(-)氨基酸(D,E);原子组成为C1694H2693N473O521S13;不稳定系数为29.76,归类为稳定蛋白(小于40时,预测蛋白为稳定);平均亲水系数(GRAVY)为-0.920,归类为亲水性蛋白(GRAVY值的范围在-2到2之间,负值表明为亲水性蛋白),且通过Phobius软件预测Sar s 14.3蛋白不含信号肽及跨膜域,但预测该蛋白位于非细胞质部分,提示能分泌到细胞外。
2.4 Sar s 14.3蛋白二、三级结构分析 SOPMA二级结构预测结果(图3)显示,Sar s 14.3蛋白α-螺旋占4.55%(15个氨基酸);β-折叠占41.82%(138个氨基酸);无规则卷曲占53.64%(177个氨基酸)。同样I-TASSER三级结构预测结果显示Sar s 14.3蛋白空间结构也主要以β-折叠和无规则卷曲结构为主(图4)。其中无规则卷曲较集中的部位形成抗原表位的可能性较大。

蓝色:α-螺旋;红色:β-折叠;紫色:无规则卷曲区域图3 疥螨Sar s 14.3蛋白SOPMA二级结构预测Fig.3 Prediction of the secondary structure of Sar s 14.3 protein of Sarcoptes scabiei by SOPMA

图4 疥螨Sar s 14.3蛋白I-TASSER三级结构预测Fig.4 Prediction of the tertiary structure of Sar s 14.3 protein of Sarcoptes scabiei by I-TASSER
2.5 Sar s 14.3蛋白同源性及系统进化分析 通过在线BLASTx对疥螨Sar s 14.3进行同源性比对发现本次通过高通量测序组装的猪疥螨Sar s 14.3蛋白序列与野猪疥螨(KAF7495137)、犬疥螨(KPM11048.1)和人疥螨(AAO15613.1)局部一致性均为100%;与粉尘螨(BAA04558.1、P39673)、埋内欧尘螨序列(OTF81155.1)和屋尘螨序列(AAM21322)局部一致性也达50%以上(表2)。

表2 疥螨猪变种Sar s 14.3过敏原蛋白序列NCBI BLASTx比对结果Tab.2 Results of NCBI BLASTx alignment of Sar s 14.3 protein sequences of Sarcoptes scabiei

表2(续)物种蛋白描述E值一致性/%序列登录号埋内欧尘螨High molecular weight allergen M-177 pre-cursor 4.00E-17058.8AAF14270埋内欧尘螨Allergen-like protein (Apolipophorin-like) 2.00E-4353OTF70165热带无爪螨Allergen Blo t Mag 1 2.00E-11349.55AAM10780热带无爪螨Group 14 allergen Blo t 14 7.00E-12846.39ABU97467椭圆食粉螨Allergen Ale o 14 Mag 1 5.00E-7045.87ABU97462.雪果绕实蝇Uncharacterized protein 2.00E-11442.17XP_017492868
2.6 pET28a-Sar鉴定结果 将提取的质粒经QuickCUtTMXhoⅠ、QuickCutTMXbaI酶切后,通过1%琼脂糖凝胶电泳检测,结果如图5所示:酶切后所得片段大小与预期结果基本一致(pET28a:5 369 bp,目的基因:1 005 bp)。
2.7 Sar s 14.3重组蛋白表达、纯化和鉴定 在选定的最佳条件下对重组表达菌诱导表达,SDS-PAGE检测细菌破碎后的上清和沉淀,蛋白主要以包涵体形式存在(图6)。将包涵体蛋白通过Ni柱层析亲和纯化后,对收集的穿流液、裂解液、洗涤液和洗脱液进行SDS-PAGE检测,检测结果显示纯化后的目的蛋白在洗脱液中有一条约40 kDa 的目的条带,第5道洗脱液中目的蛋白的量较多,纯化效果好(图7)。将纯化后的疥螨Sar s 14.3重组蛋白经聚丙烯凝胶电泳后,进行免疫印迹实验,如图8所示,纯化后的蛋白可以与疥螨兔阳性血清发生明显的反应,表明疥螨Sar s 14.3重组蛋白具有良好的反应原性。

M:Marker;1:重组质粒双酶切;:目的基因图5 重组质粒鉴定结果Fig.5 Recombinant plasmid identification results

M:Marker;1:诱导上清;2:诱导沉淀;:目的基因图6 Sar s 14.3重组蛋白可溶性分析Fig.6 Sar s 14.3 recombinant protein solubility assay

M:Marker;1:穿流液;2:变性裂解液;3:变性洗涤液;4-6:变性洗脱液图7 Sar s 14.3重组蛋白的纯化Fig.7 Purification of Sar s 14.3 recombinant protein

M:Marker;1:疥螨阳性血清(兔,多克隆抗体);2:疥螨阴性血清(兔)图8 重组蛋白 Western blot 鉴定Fig.8 Western blot identification of Sar s 14.3 recombinant protein
2.8 疥螨Sar s 14.3重组蛋白间接ELISA方法的建立
2.8.1 抗原最佳包被浓度和血清最佳稀释度的确定 使用方阵法对抗原包被浓度和血清最佳稀释倍数进行确定,结果显示,当抗原包被浓度为2 μL/mL,血清稀释倍数为1∶400,其疥螨猪阳性血清OD450与阴性血清OD450比值(P/N值)最大为6.277(表 3),表示此条件抗原包被浓度及血清稀释倍数最佳。
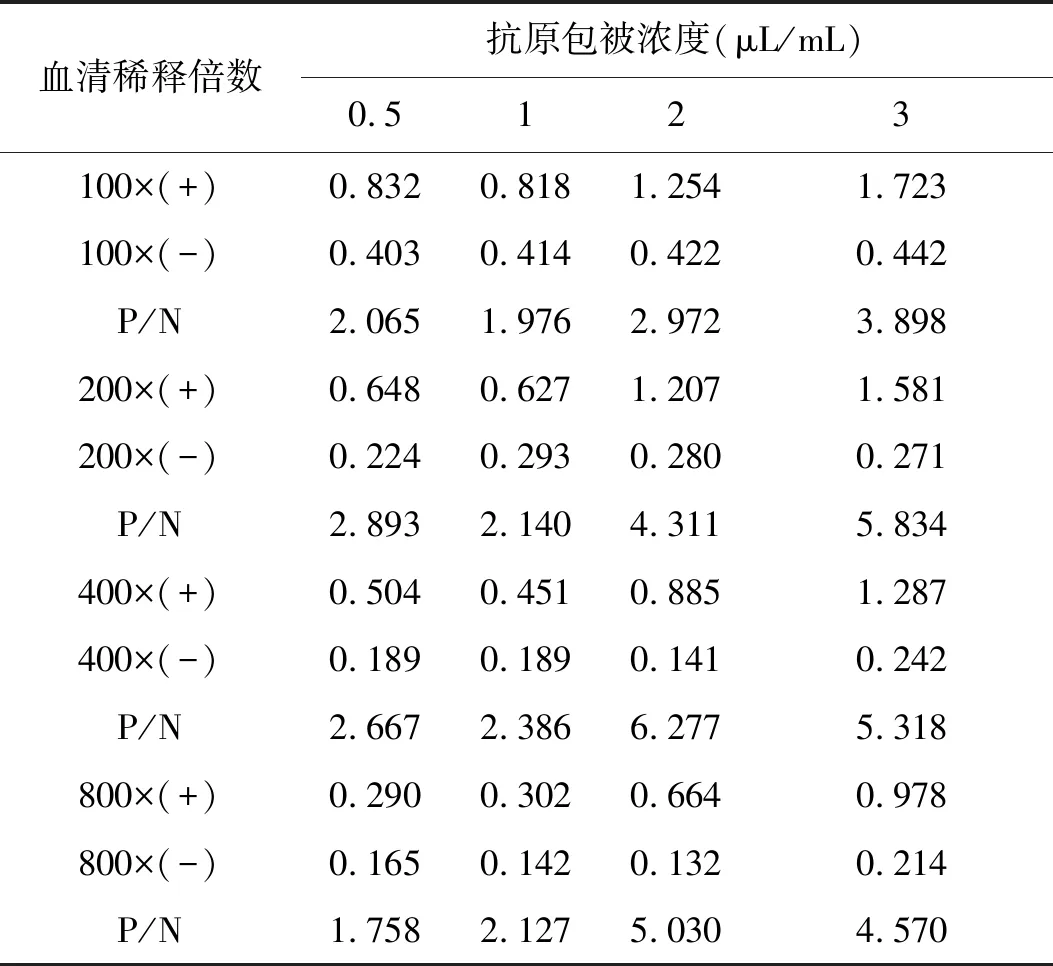
表3 最佳抗原包被浓度和血清稀释倍数Tab.3 Optimum antigen coating concentration and serum dilution ratio
2.8.2 抗原最佳包被条件的确定 抗原在4 ℃包被过夜的条件下,所测的阳性血清OD450与阴性血清比值最大(表4),可作为最佳包被条件。

表4 抗原包被最佳条件Tab.4 Optimum conditions for antigen encapsulation
2.8.3 最佳封闭液和封闭时间的确定 当使用5%脱脂奶粉,37 ℃条件下封闭1 h时,封闭效果最佳,P/N最高(表5)。

表5 37 ℃下最佳封闭液和封闭时间Tab.5 Optimum sealing fluid and sealing time at 37 ℃
2.8.4 最佳血清孵育时间和二抗稀释倍数的确定 当血清于37 ℃孵育1.5 h,二抗稀释倍数为1∶4 000时,P/N值达到最大(表6),故此可以确定该方法为最佳的孵育时间和稀释倍数。

表6 最佳血清孵育时间和二抗稀释倍数Tab.6 Optimum incubation time of serum and dilution ratio of secondary antibody
2.8.5 酶标二抗孵育时间与底物显色时间的确定 通过方阵滴定法对酶标二抗孵育时间和显色时间进行筛选,当二抗于37 ℃孵育1 h,室温显色20 min,所得P/N值最大,为最佳条件(表7)。

表7 37 ℃下最佳酶标二抗孵育时间和底物显色时间Tab.7 Optimum incubation time of HRP-conjugated secondary antibody and chromogenic time of substrate at 37 ℃
2.8.7 批内和批间重复性试验 选取3份疥螨猪阳性血清样本,使用同一批次纯化的蛋白,以最佳ELISA检测方法,每份样品重复3次,批内变异系数介于1.2%~5.3%,均小于10%,从而表示所建立ELISA检测方法批内重复性较好。另外选取3份疥螨猪阳性血清样本,使用不同一批次纯化的蛋白,以最佳ELISA检测方法,每份样品重复3次,批内变异系数介于2.5%~7.1%,均小于10%,从而表示所建立ELISA检测方法批间重复性较好。
3 讨 论
疥螨病作为一种危害生猪健康的寄生虫病,严重影响着生猪产业的健康发展。由于疥螨病易与过敏性皮炎、秃毛廯、湿疹及其他原因引起的脱毛性疾病相混淆,临床上需要对疥螨病进行鉴别诊断[2]。目前对疥螨病诊断的金标准仍然是病原学检查,但几乎有一半的疥螨病例皮屑检查看不到病原[1-2],因此血清学检查就成为疥螨病诊断尤其是早期诊断的一种有效辅助诊断方法。目前比较流行的疥螨血清学诊断方法主要是用疥螨的粗抗原或排泄抗原检查血清中的IgG或IgE抗体[1,9,11],但疥螨虫体抗原或排泄抗原较难获取,因此寻找新的诊断抗原靶标就显得极为迫切。
为了对疥螨病进行有效诊断,近年来重组过敏原用于疥螨诊断的案例不断增加[12],且表现出一定优势。本试验通过RNA-Seq.测序共获得18 980个转录本,通过与过敏原数据库中的序列进行相似性比对在疥螨猪变种转录组中共发现390条过敏原相关序列,且大部分过敏原都与尘螨属过敏原有一定的同源性。这些预测的过敏原序列将极大的减少对疥螨诊断抗原筛选的盲目性。EggNOG分类显示,预测的疥螨过敏原分子中的多数包含在下列分类:“碳水化合物转运及代谢(G:Carbohydrate transport and metabolism)”、“翻译后修饰、蛋白质周转及分子伴侣(O:Posttranslational modification,protein turnover,chaperones)”和“细胞骨架(Z:Cytoskeleton)”,提示过敏原分子能参与多种生物学进程。
Sar s 14.3过敏原序列首先由Fischer等在人疥螨cDNA文库中筛选出来,其克隆出的mRNA长度为991 bp,编码330个氨基酸[13],位于相当于粉尘螨过敏原第14组分(Der p 14)氨基酸序列的1263-1655区间[13]。随着近几年多个不同物种宿主来源疥螨基因组的解析,发现疥螨基因组中过敏原第14组分编码基因(Sars14)有5 004个核苷酸,可编码1 667个氨基酸,Sar s 14.3氨基酸序列位于Sar s 14编码蛋白氨基酸序列的1306-1585区间[3-4],且其对普通感染疥螨的人血清的IgE有高度的结合力[12]。本实验通过高通量测序亦发现在疥螨猪变种中同样有Sars14.3基因的转录(FPKM=55.1),且翻译的蛋白质序列与人疥螨Sar s 14.3蛋白序列相似性为100%。通过生物信息软件分析发现Sar s 14.3蛋白二级结构主要以无规则卷曲和β-折叠为主,phobius在线软件预测其不含信号肽和跨膜域,但位于非细胞质的部分,由于Sar s 14.3蛋白氨基酸序列为Sars14.3基因编码蛋白的部分序列,且全长Sar s 14蛋白中有一段信号肽序列,推断Sar s 14.3蛋白很可能分泌到细胞外参与寄生虫-宿主互作过程。
从疥螨各变种Sar s 14.3蛋白氨基酸序列的BLAST局部比对结果来看,疥螨各变种中Sar s 14.3蛋白氨基酸序列相似性很高(大于99.7%)。以往的研究认为虽然不同宿主来源的疥螨其生物特性有所不同,甚至还具有一定的宿主特异性[1-2],但通过分析疥螨各物种分离株的核糖体ITS2基因序列[14-15]和已测序的基因组[3-4]来看,不同宿主来源疥螨应为同一个种。本研究通过分析疥螨Sar s 14.3蛋白氨基酸序列也同样得到相似的结论。以上结果说明即使是用不同宿主来源的疥螨重组过敏原Sar s 14.3分子也可以诊断来自其他动物和人的疥螨病。
近年来,随着社会经济的发展,国民的生活水平也在不断地提高,促使宠物养殖和国家的畜牧业发展也越来庞大。目前畜禽的养殖模式都是集约化的养殖,动物生长的环境比较拥挤、潮湿,这些条件都很适合疥螨病的传播和流行。目前针对疥螨病的诊断主要是病原学诊断,在潜伏期和早期很难直接检测到虫体或虫卵[16]。可以诊断疥螨病的检测试剂盒多是采用成虫粗抗原检测血清中总免疫球蛋白,这种方法的缺点在于假阴性和假阳性率较高,诊断抗原制作困难,并且难以形成标准化。因此,应用成分明确的重组抗原替代粗抗原,建立高灵敏度的诊断方法,是免疫诊断试剂盒研制的必然趋势。
ELISA是一种用于检测特异性抗体和可溶性抗原的异质免疫分析技术,可用于定量或定性分析,无需复杂或昂贵的设备[17],是目前应用较为广泛的血清学诊断方法。该方法在特定酶的催化作用下,抗原抗体反应被放大,具有很高的敏感度[18]。虽然用PCR检测疥螨准确率较高[19],但相对于PCR检测方法,间接ELISA检测法操作时间更短、所需劳动量更少且单次可以检测大量的样品[20]。ELISA实验数据准确性的关键是要降低非特异性反应的结合,本试验建立的间接ELISA方法仅以纯化后的疥螨Sar s 14.3重组蛋白作为包被抗原,可以有效的降低假阳性的产生。本试验结果中阳性和阴性血清的OD值都偏高,这可能是由于蛋白的浓度偏高。经过多次筛选和不断对间接ELISA检测方法的条件进行优化,最终确定的最佳抗原包被浓度为2 μL/mL,在4 ℃包被过夜,用5%脱脂奶粉,37 ℃条件下封闭1 h。验证所建立的间接ELISA诊断方法时,批间和批内的变异系数均小于10%,说明其重复性良好,该重组蛋白适合作为检测抗原。由于本试验中未找到感染其他寄生虫的患病猪的阳性血清,但本实验室保存有人工实验感染棘球蚴、贾第虫和胞内劳森菌的小鼠阳性血清检测,用上述小鼠的血清与疥螨猪标准阳性血清和阴性血清进行了交叉实验,结果显示上述感染小鼠血清OD450值均小于0.341,结果为阴性,说明本次建立的间接ELISA方法有一定的特异性,但还需与一些同源性更近的患寄生虫病尤其是痒螨病的病猪的阳性血清来比较才能使建立的间接ELISA诊断方法的特异性检验更加确信。
本试验通过高通量测序获得了18 980条疥螨转录本序列,并预测出其中有390条转录本与过敏原有同源性,所预测的过敏原序列主要参与与碳水化合物转运、翻译后修饰、蛋白质周转及细胞骨架构成等生物学过程。通过对猪疥螨Sar s 14.3蛋白结构及功能分析预测显示,该蛋白很可能分泌到细胞外与宿主互作,同时同源性分析提示疥螨Sar s 14.3与尘螨遗传距离较远,疥螨属各变种遗传距离非常近,可以用一个物种中克隆表达出的Sar s 14.3重组抗原诊断不同动物和人的疥螨病。
利益冲突:无
猜你喜欢
吉林畜牧兽医(2022年1期)2022-11-16
现代畜牧科技(2021年3期)2021-07-21
中国畜禽种业(2020年5期)2020-01-12
中国乳品工业(2018年10期)2018-11-16
饲料博览(2018年8期)2018-03-19
农民致富之友(2017年2期)2017-03-23
现代农业(2016年5期)2016-02-28
上海故事(2015年13期)2016-01-22
现代畜牧科技(2015年1期)2015-10-21
伴侣(2015年5期)2015-09-10
