
生物法抑制硫酸盐还原菌在油田的应用研究进展*
2023-02-15 09:12陶建强
安全、健康和环境 2023年1期
陶建强
(中国石化新疆新春石油开发有限责任公司,山东东营 262500)
0 前言
目前,我国大部分油田已进入二次、三次采油阶段,大量的水和化学剂(高分子聚合物、表面活性剂等)注入地层,引起地层内部、地面系统等的微生物种群发生变化,导致一些有害细菌滋生,影响油田的正常生产。

本文从SRB对油田生产过程的危害、生物法抑制SRB的机理及影响因素、现场应用等方面对生物法抑制SRB技术进行综述,以期为该技术的推广应用提供借鉴。
1 SRB对油田生产过程的危害
SRB及其代谢产物(硫化氢)给油田生产带来诸多危害,如油藏变酸、设备及管线腐蚀、聚合物溶液黏度损失等。
1.1 导致油藏变酸

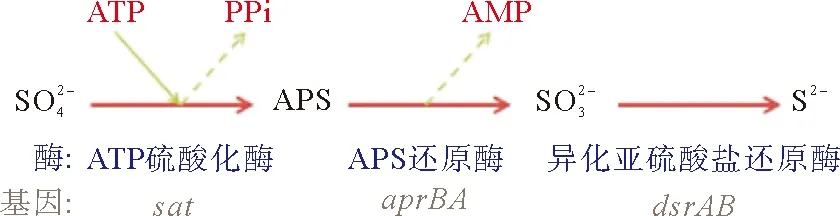
图1 油藏变酸机理[10]
1.2 加剧腐蚀
SRB是造成油田生产系统管线、设备腐蚀的主要微生物。SRB导致金属腐蚀由荷兰学者于1934年提出,此后逐渐得到重视。SRB引起腐蚀的机理主要有去极化、代谢产物腐蚀(MIC)、膜下腐蚀、浓差电池、细胞外电子转移MIC(EET-MIC)等[11,12]。张润杰,等[13]研究认为SRB吸附在碳钢表面,利用金属中的电子进行生命活动,相当于强化阴极过程,加剧腐蚀。
1.3 导致聚合物黏度损失
SRB引起聚丙烯酰胺(HPAM)溶液黏度降低的原因主要有2个:①SRB直接以HPAM为碳源或氮源进行生长繁殖,HPAM被降解为小分子物质,引起黏度降低[14,15];②SRB代谢产生的S2-以及与SRB腐蚀产生的Fe2+等引起HPAM分子链断裂,降解产物被SRB作为营养底物利用,促进SRB生长繁殖,进一步降解HPAM[16]。
2 NRB抑制SRB技术


表1 分离自油田的具有硝酸盐还原功能的细菌
2.1 NRB抑制SRB机理
2.1.1 营养底物竞争
NRB和SRB同属于厌氧型微生物,生活习性、生长环境相似。当基质和营养底物不足时,便产生了对营养物的竞争。NRB对营养物的亲和力更强,当营养物浓度较低时更占优势。Zhang,等[24]研究发现,当向室内搭建的反应器加入硝酸盐后,以碳源为电子受体的NRB丰度增加,SRB丰度降低,COD消耗显著,表明被激活的NRB加速了碳源的利用。但研究也发现,NRB的基质降解速率比SRB低[25],在营养物浓度足够高时,SRB能有效利用基质进行生长代谢,NRB对SRB的抑制作用不明显。马放,等[26]的研究表明,当COD浓度为120 mg/L时,系统中的硝酸盐氮消耗明显,亚硝酸盐氮有一定累积,但硝化过程进行不完全;硫酸盐还原反应基本停滞,硫离子在初期有少量生成,随后消失至反应结束。但当基质浓度高时,NRB对SRB的抑制作用减弱。油藏环境中有机质一般不足,大多数油田环境中可溶于水的挥发性脂肪酸(VFA)含量较低(低于500 mg/L),注入合适的硝酸盐或反硝化菌,异养反硝化菌能利用油层中的VFA快速生长,变为优势菌群,反硝化菌对营养的利用使SRB失去赖以生长的营养底物,代谢受到抑制,活性降低,数量减少[27]。
Hitzman,等[28]向油藏中注入硝酸盐激活NRB,使NRB与SRB竞争环境中的挥发性脂肪酸,异养反硝化菌成为优势菌群,从而抑制SRB产生硫离子。Casey Hubert,等[29]研究了NRB对油藏变酸的控制及与SRB竞争营养底物的作用,发现加入硝酸盐后,生物种群结构从以SRB为主变为以NRB为主,同时出现了硝酸盐还原硫氧化菌(NR-SOB)和异养NRB,前者可以直接氧化还原性硫,可见NRB通过底物竞争阻止了SRB代谢(图2)。2种NRB均可以通过反硝化作用抑制SRB。

图2 竞争营养底物抑制原理
2.1.2 硫氧化细菌直接氧化硫离子
激活硝酸盐还原-硫氧化细菌(NR-SOB)可以利用硝酸盐或亚硝酸盐将硫离子重新氧化为单质硫和硫酸盐[9],抑制机理如图3所示。NR-SOB通过同时氧化硫离子和还原硝酸盐,而非与SRB争夺营养,抑制硫酸盐的还原速率。胜利油田检出的Thauera(Rhodocyclales目)菌可在硝酸盐存在的厌氧条件下氧化硫离子[30]。Gevertz,等[31]通过基因组预测证明了Arcobacter能够以硝酸盐为电子受体完成硫化物的氧化过程。此外,油藏中还发现了大量硫氧化细菌(SOB),如Bosea, Methylobacterium, Paracoccus, Pseudomonas和Halomonas等,它们可以通过氧化硫离子获得能量,例如Paracoccus能够在有氧条件下利用硝酸盐氧化不同价态的硫,Pseudomonas和Halomonas氧化硫离子的过程也与硝酸盐还原相关[20,26,32]。

图3 NR-SOB消除硫化物的原理
2.1.3 中间产物抑制机理


Hao,等[39]发现一些SRB(如Desulforhopalus、Thermodesulfobium等),即使在硝酸盐浓度较低时仍具有较强的亲和力,这类SRB优先利用硝酸盐,当硝酸盐利用殆尽时才利用硫酸盐。这类细菌的存在,可能会对单纯利用化学分析手段监测生物抑制SRB的效果给出假性结果,因为在硝酸盐量足够时,该类细菌以相对低的丰度存在于群落中,一旦硝酸盐消耗殆尽,该类细菌大量滋生,导致生物抑制SRB失败。
2.1.5 提高氧化还原电位

2.2 NRB抑制SRB的现场应用
Sturman[43]带领美国生物膜工程中心研究团队在新墨西哥的San Juan盆地和荷兰阿莫科Rijn的油田开展了注入硝酸盐抑制SRB、控制硫化氢的现场应用研究。注入的亚硝酸盐在抑制SRB活性的同时,能去除已经生成的硫化氢,且有效期较长。在脱水器处投加硝酸盐后,水中硫化氢含量由40~60 mg/L降至低于0.5 mg/L,SRB含量由103~104个/mL降至10个/mL以下。利用LPR探头监测脱水器处添加硝酸盐后的腐蚀情况发现,未添加硝酸盐前的腐蚀速率为0.03~5 mm/a,添加硝酸盐后腐蚀速率稳定在0.2~0.3 mm/a之间。在生产井A2进行的实验显示,硫化氢由6 mg/L降至未检出,采出水中的铁离子含量增加近20倍(达到10 mg/L),日产原油量增加27.4~54.8 t,这是由于FeS沉淀的溶解以及阻止新的FeS沉淀生成,解除了地层堵塞。由于实验前的腐蚀速率缺失,但试验期间的腐蚀速率没有升高。可见该技术同时起到了降低腐蚀速率和提高采收率的效果。
Anchliya[44]针对SRB引起的油藏变酸、腐蚀等问题,提出了SRB生物控制措施。通过向未被污染的油藏注入硝酸盐体系,预防SRB滋生和硫化氢生成。对于已经变酸的油藏,硝酸盐基处理技术可以去除油藏、采出液、井筒、管线中的硫化物,控制硫化氢进一步生成。在北海油田的应用效果显示,超过90%的S2-被消除,腐蚀速率降至0.2 mm/a,下降了71.4%,该技术在Statfjord、Valhall等油田均得到推广应用。
G E Jenneman,等[45]在Coleville油田开展了利用本源微生物去除硫离子的现场试验。在两口注入井连续注入硝酸铵和磷酸二氢钠,浓度分别为400 mg/L和12 mg/L。两口注入井的注水量分别为224 m3/d和45 m3/d,处理前的硫化物含量分别为140 mg/L和112 mg/L,SRB和NRB的含量均在103~105之间。注入激活剂后,NRB的数量增加到108个/mL,SRB数量得到控制,注入50天后停注,NRB数量有所下降,SRB数量最低降至100个/mL,说明优势菌群发生了改变。
Hitzman[46]的研究团队对厌氧条件下NRB作用的研究发现,硝酸盐的注入能够激活油藏内的NRB,使油藏菌落结构发生改变,SRB不再占优势地位,在控制硫化物的同时,增加了原油产量。针对油田实际生产工况,随回注水伴注硝酸盐/亚硝酸盐混合体系,比只加硝酸盐更加有效。经过处理后,硫化物的产生量由处理前的400 kg/d持续降低至完全消除,50口生产井的产量由10 000 t增加到10 960 t。
国内利用NRB抑制SRB的技术在乌南油田、大庆油田、胜利油田等也开展了现场试验,取得了不错的效果[47]。其中,胜利油田开展了利用NRB抑制SRB,进而提高污水配聚黏度的规模化现场应用,实施效果见表2。该技术实施后,污水硫离子含量由2~4 mg/L降至0.2 mg/L以下,配聚黏度由33.2 mPa·s上升至53.2 mPa·s,且保持长期稳定(大于30 d)。对群落结构进行分析发现,生物抑制后,Thauera、Petrobacter、Paracoccus、Marinobacter、Hyphomonas、Hyphomicrobium等具有硝酸盐还原功能的细菌丰度增加,表明激活剂的加入激活了NRB,与SRB竞争营养底物并限制其活性。

表2 NRB抑制SRB在胜利油田的应用效果对比
3 NRB抑制SRB技术需进一步解决的问题
如前所述,NRB抑制SRB技术的机制复杂,现场应用效果差别较大。从基因和蛋白组学角度,借助现代生物分子生态学和分子示踪等技术揭示微生物代谢途径及菌群间的相互关系,是该技术在机理和应用研究中重要研究方向。
3.1 各菌群间的相互关系需进一步明确
油田污水系统是一个复杂的“生物反应器”,含有丰富的微生物种群,除了NRB、SRB、硫氧化菌外还有铁细菌、石油烃降解菌、产甲烷古菌等,这些细菌均会利用碳源进行生长繁殖,受各菌群代谢途径、环境因子、生物酶、底物传递等多种因素影响,生物竞争抑制过程中各菌群之间的相互作用机制复杂,反应动力学丰富(图4)。明确各菌群间的相互作用及对营养底物的竞争利用能力具有重要意义。近年来,基因定量技术、新一代测序技术、宏组学技术的发展,可以有效地揭示群落水平和个体水平的代谢能力和竞争合作关系,多维宏组学与环境依赖型实验可以有效地阐释影响降解表现的相互关系,使解析生物竞争抑制过程中复杂的代谢途径及各菌群的相互关系有了技术保障,可以更加充分地了解复杂的混合生物关系。

图4 硫酸盐和硝酸盐还原途径中部分关键酶的表达[24]
3.2 碳-氮-硫转化相关的生物化学过程需进一步揭示
油田环境中碳-氮-硫转化相关的生物化学过程复杂(图5),目前的研究主要基于IC、HPLC、GC-MS、NMR等仪器分析手段,通过检测抑制过程中代谢产物的含量,推断碳-氮-硫转化的动力学模型,但在抑制过程复杂的微生物环境中,常规的化学分析手段不能检测到完整的代谢产物。借助分子生物学手段确立抑制过程中的关键物种和核心物种,揭示不同微生物菌群(SRB、NRB、硫氧化菌和石油烃降解菌等)潜在参与的碳、氮、硫代谢具体途径,明确碳-氮-硫转化的生物化学过程,对指导NRB抑制SRB技术应用具有重要意义。

图5 碳氮硫转化的部分过程
4 结语与展望
采用生物法抑制SRB具有低成本、无二次污染与其他化学剂配伍性好等特点,是控制SRB的有效手段。
目前,关于生物法抑制SRB的主导机理为[20,24,48]:①通过注入功能性激活剂(硝酸盐、亚硝酸盐等)激活内源硝酸盐还原菌群(DNB)或者注入外源的DNB,与SRB竞争生活空间和营养底物,利用DNB对营养基质的优先利用,导致SRB所需营养不足而使活性受限甚至衰亡,抑制硫化物产生;②激活硝酸盐还原-硫氧化菌(NR-SOB),利用硝酸盐/亚硝酸盐的生长代谢,并直接将硫离子转化为单质S或硫酸盐;③亚硝酸盐、氧化亚氮和其他代谢中间产物可抑制SRB并去除硫离子,尤其是亚硝酸盐可抑制异化亚硫酸盐还原酶,阻断硫化物产生。
过去的20年里,尽管已开展了许多利用生物法抑制SRB的室内研究与现场试验,但成功应用的实例不多。主要原因是在抑制机理和工艺生态调控等方面的研究不足。投加抑制剂(如硝酸盐、钼酸盐、DNB等)对SRB的基因和蛋白表达的影响不明确,对微生物种群的组成、群落结构动态演替规律剖析不足,生物法抑制SRB提高配聚黏度过程中激活的功能菌与SRB的相互关系及聚合物黏度提高的内在机制不明确[49,50]。此外,添加硝酸盐抑制SRB的潜在负面影响尚不明确,如关于添加硝酸盐是否加剧腐蚀的文献报道意见不一致。在没有H2S存在时,NRB代谢的中间产物(亚硝酸盐)可能会加剧腐蚀。Rizk,等[10,21,51]证实了亚硝酸盐(浓度低于10 mg/L)与腐蚀增强之间的相关性,低浓度亚硝酸盐可能会诱发点蚀。存在H2S时情况更加复杂,因为亚硝酸盐与H2S直接反应产生的单质硫、硫代硫酸盐等物质也可能引发腐蚀。抑制过程中激活的NR-SOB可以将硫离子氧化为单质硫,加剧局部腐蚀。因此,生物法抑制SRB的效果应根据单个应用试验进行针对性的评估。
由于生物法抑制SRB的效果与油层本源微生物的群落结构、油层物理化学特性、激活剂加量、激活优势菌群结构、群落间相互关系等密切相关。因此,借助快速发展的分子生物学技术及化学分析手段,解析抑制过程的菌群代谢机制与相互关系,明确抑制有效性与激活剂添加条件、群落结构演替等之间的关系,进一步探明抑制机理对于该项技术的大规模应用具有重要意义。
猜你喜欢
中老年保健(2022年3期)2022-11-21
现代畜牧科技(2021年6期)2021-07-16
科学(2020年2期)2020-08-24
当代水产(2019年6期)2019-07-25
当代水产(2018年12期)2018-05-16
恋爱婚姻家庭·青春(2016年7期)2016-07-05
电源技术(2015年10期)2015-08-01
中国果业信息(2015年12期)2015-01-24
西安建筑科技大学学报(自然科学版)(2014年5期)2014-11-10
天津医科大学学报(2011年4期)2011-07-13
