
植物ABC转运蛋白功能研究进展
2023-02-15 07:26唐桃霞孔维萍任凯丽
西北农业学报 2023年1期
唐桃霞,孔维萍,任凯丽,程 鸿
(甘肃省农业科学院 蔬菜研究所,兰州 730070)
ABC转运蛋白,又名ATP结合盒式蛋白(ATP-binding cassette transporter,ABC),得名于含有一个腺苷三磷酸(ATP)结合盒,且依赖于 ATP 水解产生的能量而实现底物在细胞内外的跨膜转运,是目前已知数量和功能最多的一类蛋白[1]。据报道,ABC转运蛋白家族在所有生物中广泛存在,已在人类、酵母和大肠杆菌中发现了数量不等的ABC转运蛋白及其基因,而在陆生植物中鉴定到的 ABC 转运蛋白数量远高于其他生物[2-3],其中,模式作物拟南芥和水稻基因组中分别存在130个和128个 ABC 转运蛋白[4-5],此外,玉米(Zeamays)[6]、小麦(Triticumaestivum)[7]、甘蓝型油菜(Brassicanapus)[8]、百脉根(Lotuscorniculatus)[9]、菠萝(Ananascomosus)[10]以及番茄(Solanumlycopersicum)[11]中均有报道,而研究最多的拟南芥仅有一小部分ABC 转运蛋白被鉴定出来[12],还有大部分成员的转运底物及其功能尚不清楚,而有关其他作物ABC转运蛋白及其功能还未见报道。因此,本文通过对已鉴定的植物ABC转运蛋白成员及功能进行归类总结,旨在为研究其他作物ABC转运蛋白及功能提供思路和依据。
1 ABC转运蛋白的结构及分类
典型的ABC转运蛋白由亲水核苷酸结合结构域(Nucleotide-binding domain, NBD)和疏水跨膜结构域(Transmembrane domain, TMD)组成[13],其中亲水性的 NBDs 包含3个保守的特征基序:Walker A,Walker B 和ABC 标签序列(又称 Walker C)[4],因此,其结构和序列在 ABC 转运蛋白家族中高度保守,而疏水的TMD是由数个跨膜 α- 螺旋构成,形成跨膜运输通道[14],转运时NBDs负责结合并水解 ATP 使其构象发生改变,从而激活转运过程,而 TMDs则形成通道使得底物通过脂质双分子层,二者共同参与实现物质转运[15]。ABC 转运蛋白家族的转运分为向外和向内两种转运方式,大多数的ABC向内转运蛋白还包括一个可溶性的位于细胞外或是锚定在膜上的胞外区结合蛋白(Substrate-binding protein,SBP),SBP 可以结合不同种类的底物并把底物传递到相应的 TMDs 中。图1展示了ABC 转运蛋白转运过程中依靠构象改变的转运机制[16]。
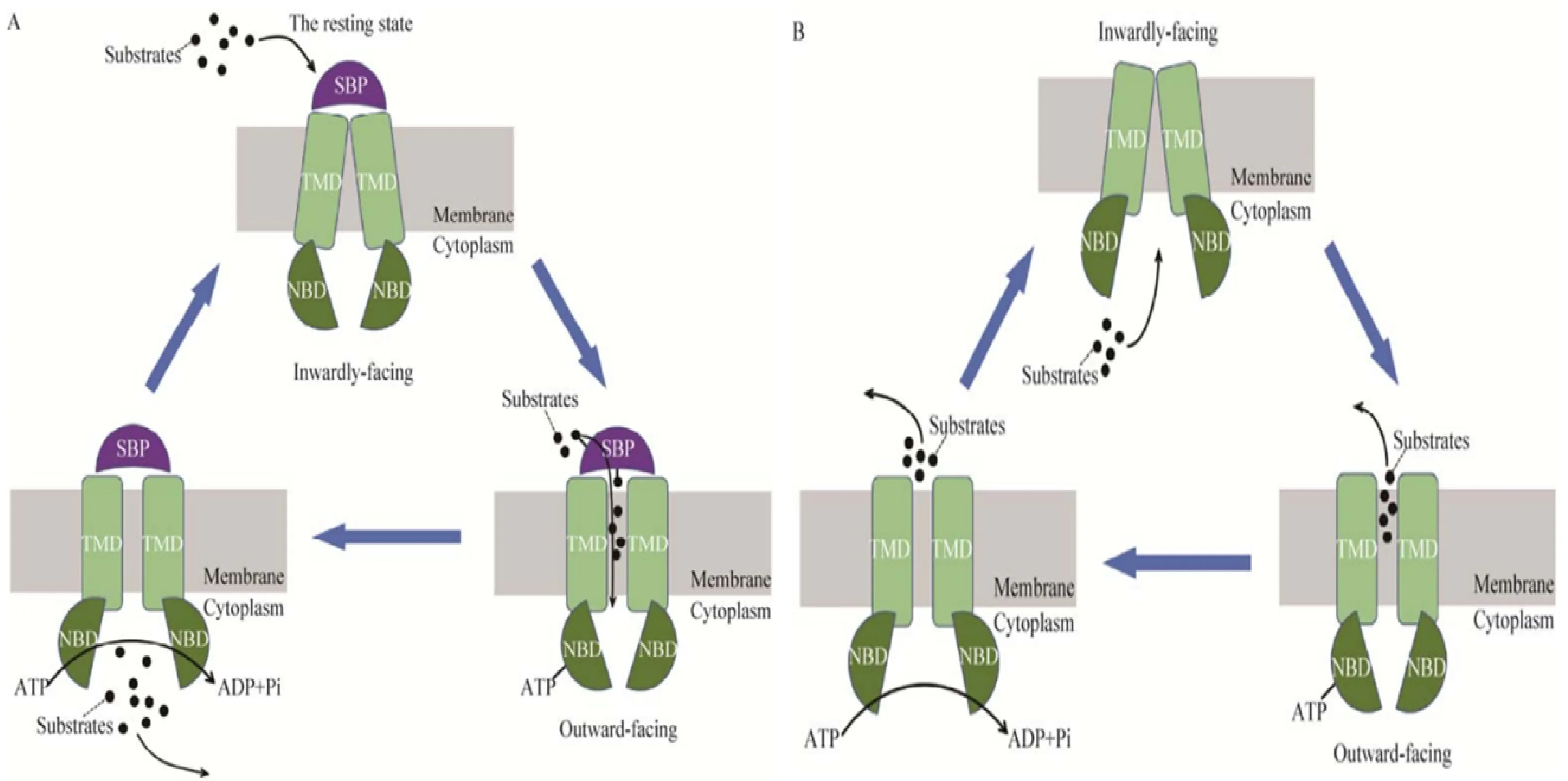
A.向内转运机制;B.向外转运机制A.Import mechanism; B.Export mechanism
由于 ABC 蛋白包含的NBD 和 TMD 数量以及排列方式存在差异,其又被分为全分子 ABC 蛋白、半分子ABC 蛋白和可溶性ABC蛋白,全分子 ABC 蛋白由 2 个 NBD 和 2 个 TMD构成,4 个结构域结合在一起才可执行转运功能,而半分子 ABC 蛋白仅由 1 个 NBD 和 1个 TMD 构成,从而排列形成 TMD-NBD 或NBD-TMD,任一的 TMD-NBD 或 NBD-TMD 是没有功能的,它们必须形成同源或者异源二聚体才能发挥功能[17]。通常,研究者们按照国际命名系统将植物ABC分为8个亚族(ABCA~ABCG和ABCI),植物中未发现ABCH 亚族成员,但存在一类特殊的细菌类型 ABC 转运蛋白,被命名为 ABCI 亚族[1]。目前,研究较多的为 ABCG、ABCB和ABCC[18]。ABC 转运蛋白参与植物体中多种重要的转运过程,其转运功能需要NBD和TMD的共同参与。由于NBD的高度保守性和每个亚族结构的相似性,导致其转运功能具有专一性,也具有相似性。
2 ABC转运蛋白功能
2.1 ABCA亚族
迄今为止,在人类基因组中已鉴定出12个ABCA亚族成员,主要与身体各部位的脂质转运有关[19],而拟南芥中的 ABCA1 与人类 ABCA1具有高度的序列同源性,推测植物ABCA1 也可能具有脂质运输功能[20]。目前,在拟南芥中也发现12个ABCA成员,且只有 At ABCA9的功能被鉴定出来,并发现其过量表达可促进种子三酰甘油 (Triacylglycerol,TAG)的生物合成,表明AtABCA9 可能是增加油籽脂质产量的有力工具[12,21],随后李健春[22]在大豆中发现GmABCA7转运蛋白,并将其定位于过氧化物酶体,表明其在拟南芥中的过量表达影响脂肪酸的 β氧化并促进种子萌发;此外,拟南芥根的转录组分析表明 AtATH14 和 AtATH15 表达对盐胁迫有反应[23],也有报道AtABCA1 与花粉萌发、种子萌发和种子成熟有关[9,24],但具体功能未做解析。
ABCA包括全分子的AOH和半分子的ATH两种类型。在蔬菜作物番茄中发现,SlABCA1 是唯一全分子ABCA 和最大的 ABC 蛋白,由1 910 个氨基酸残基组成,另一主要特征是 ABCA 亚族在双子叶植物中存在一个全分子的ABCA (AOH),包括番茄[11]、拟南芥[4]、百脉根[9]和葡萄[24],截至目前还未在水稻[25]、玉米[6]等单子叶植物中发现,这表明这种全分子 ABCA可能特定于双子叶植物;据番茄[11]的基因表达谱显示 SlABCA1 和 SlABCA2 优先在根中表达,它们可能参与根的分泌活动,SlABCA4-7 在花中特异性表达,表明在花器官中具有特定功能。此外,吴鹏等[26]在黄瓜中克隆到控制ABCA19转运蛋白的基因,通过功能分析发现黄瓜果实中的AB-CA19对霜霉威胁迫的响应比较迅速。以上研究只是对ABCA 成员做了初步表达,其转运底物可能比较多样,具体功能还需进一步解析验证。
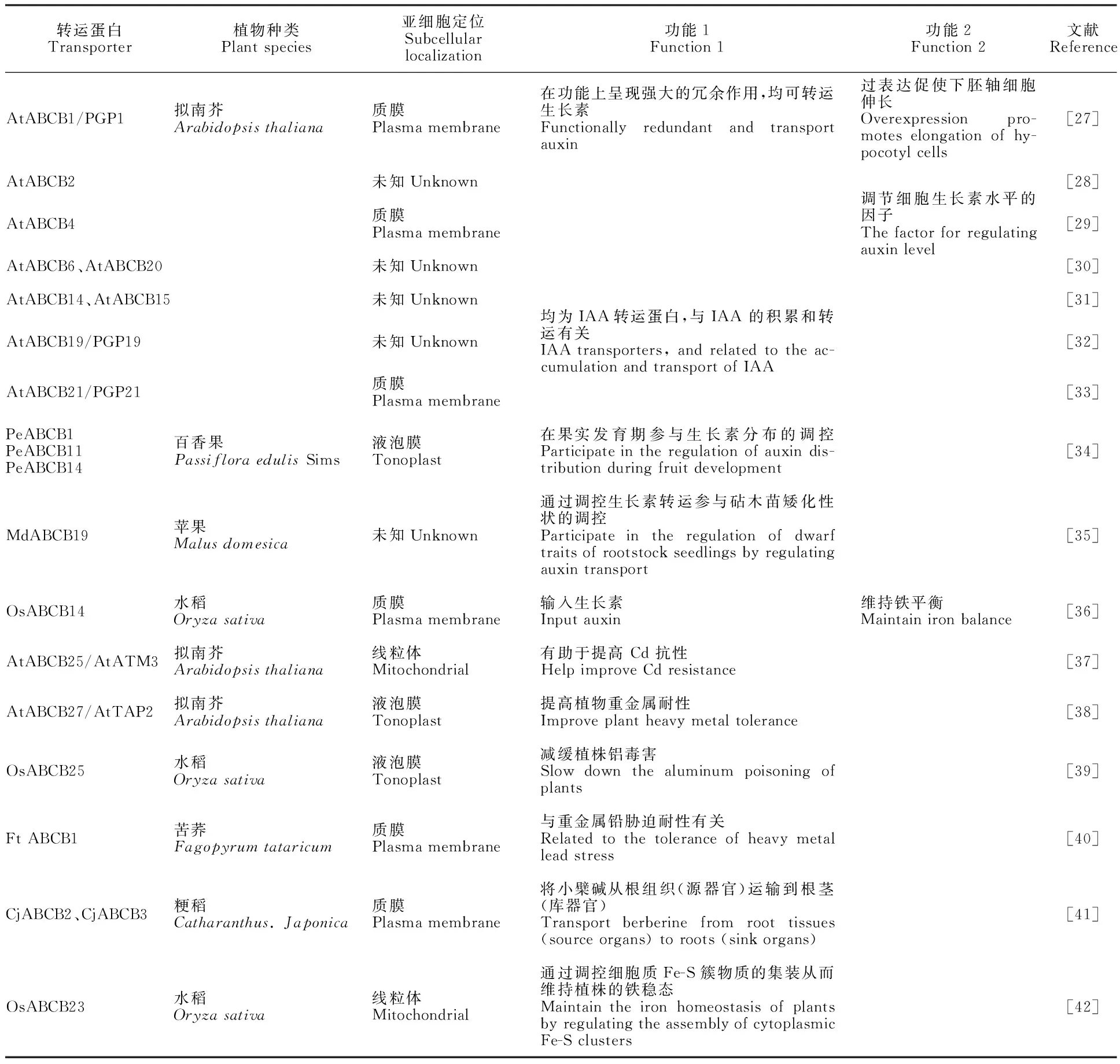
表1 植物ABCB转运蛋白功能Table 1 Function of plant ABCB transporters
2.2 ABCB亚族
ABCB是仅次于ABCG的第二大亚族,拟南芥中发现29个ABCB成员[4]。据表1可知,已鉴定的大多数ABCB成员主要与激素类物质(生长素)的运输及调控有关,且部分成员功能冗余,均能转运生长素;其次,ABCB还参与离子及重金属转运,如在番茄中研究发现大多数 SlABCBs 在根中高度表达,这可能表明这些 SlABCB 参与根中的离子和重金属运输[11]。除上述功能外,ABCB还具有转运生物碱和维持铁稳态的作用。综上可知,还需加大对ABCBs功能的研究,以期为阐释植物对激素和非生物胁迫响应的生理机制奠定基础。
2.3 ABCC亚族
根据国内外研究报道发现,C亚族目前已鉴定的功能之一是转运代谢产物(花青素、植酸、叶酸等物质),有研究表明,玉米ZmMRP3和葡萄VvABCC1均具有运输液泡中花青素的功能[43-44];拟南芥的AtABCC2可能与其他的ABCC转运蛋白共同介导营养组织中花青素和其他类黄酮的液泡运输[45];还有研究表明,部分ABCC转运蛋白具有多种功能,如拟南芥AtABCC5/AtMRP5[46]和水稻 OsABCC13[47]参与种子植酸的转运和含磷产物的积累,其中AtABCC5/AtMRP5还是保卫细胞脱落酸和 Ca2+信号转导过程中保卫细胞离子通道的中央调节器[48];而拟南芥AtABCC4/AtMRP4不仅参与叶酸转运[49],还介导气孔孔径的复杂调节[50]。
二是具有解毒功能,拟南芥AtABCC1、AtABCC2和AtABCC3均定位于液泡膜,其中AtABCC1和AtABCC2被证明赋予对砷、镉和汞的耐受性[51-53],是因为植物螯合素 (PC) 与Cd需要以 PC-Cd 复合物的形式运输到液泡中起到解毒作用,而AtABCC3是一种PC-Cd 复合物的转运蛋白,其活性受Cd 调节并与AtABCC1/AtABCC2 的活性协调;此外,AtABCC6 在重金属Cd胁迫下表达上调,也参与重金属解毒作用[54];苦荞FtABCC9在重金属Pb胁迫下表达上调,说明此蛋白也响应重金属胁迫[40]。这些转运蛋白为植物基因工程提供了有用的工具,可使植物增强对金属耐受性和积累,这是植物修复的理想特征[52]。
2.4 ABCG亚族
在ABC转运蛋白家族中,ABCG是其中最大的一个亚族,其功能广泛,有全分子的PDR和半分子的WBC两种类型。已知的拟南芥、亚麻、水稻、玉米、铁皮石斛、毛果杨、二穗短柄草、葡萄、番茄、菠萝及蜜柚中分别有43、85、50、54、37、74、44、71、70、42和22个ABCG转运蛋白(表2),分别占ABC蛋白的33.08%、42.93%、39.06%、41.54%、42.05%、38.54%、33.08%、39.23%、45.45%、42%和44%,平均数量占ABC家族的40%以上,说明ABCG是ABC家族最为重要的成员。从已报道的研究来看,已鉴定ABCG成员的功能多样,具体类别见表3。
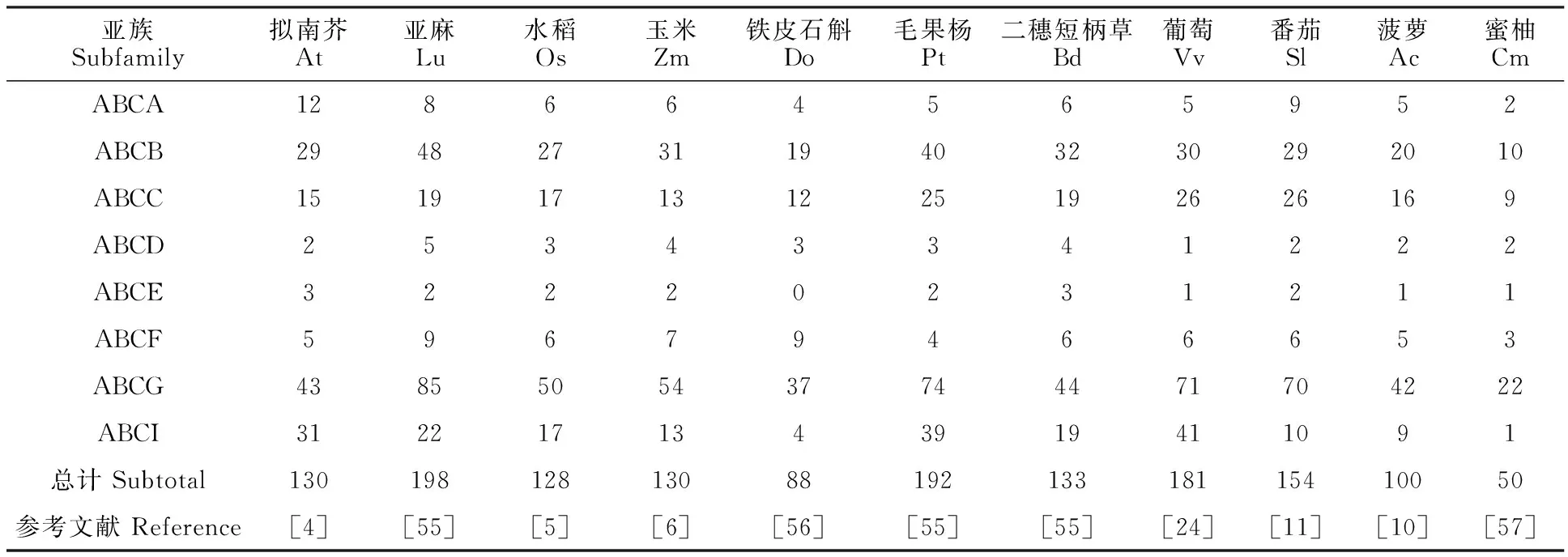
表2 部分植物中ABC转运蛋白数量Table 2 Number of ABC transporters in some plants
由表3可知,ABCG是植物体生命活动的重要参与者。首先,可转运植物体内的代谢物,包括脂类、烯类,生物碱类等,还调节花散发挥发性化合物的功能,除转运内源代谢物外,还可转运外源物质百草枯等除草剂。其次,介导激素类物质的运输(ABA),如AtABCG25、AtABCG31、AtABCG30、AtABCG40、AtPDR12/AtABCG40均具有转运脱落酸(ABA)的作用。此外,ABCG参与重金属胁迫/逆境胁迫响应,可提高植物对非生物胁迫的抗逆性。
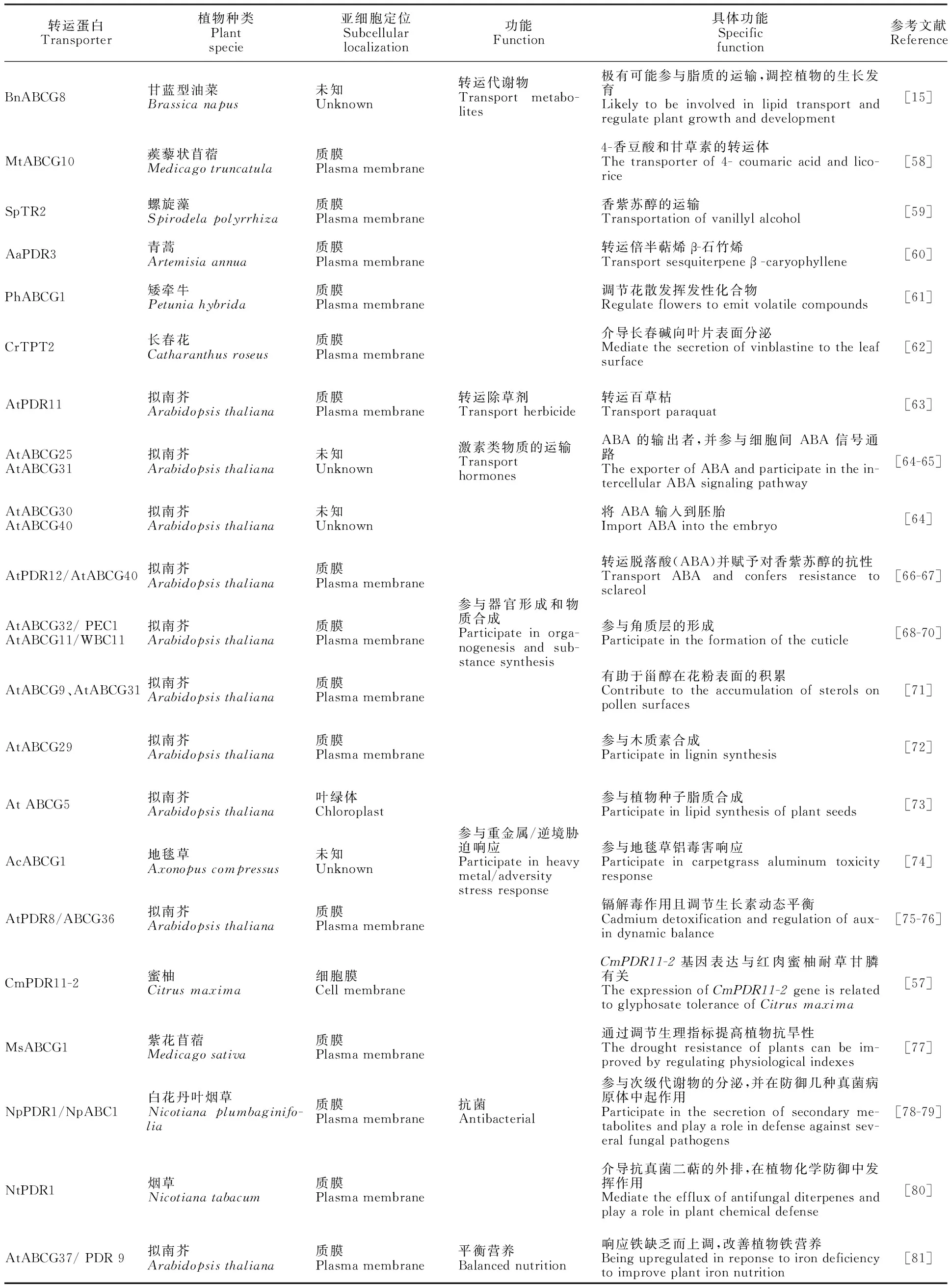
表3 植物ABCG转运蛋白功能Table 3 Functions of plant ABCG transporters
植物角质层不仅减少水分流失,还具有化学防御功能,研究发现AtABCG32/PEC1和AtABCG11/WBC11均参与角质层的形成[68-70],ABCG9 和 ABCG31 通过促进甾醇在花粉表面的积累而支持花粉外壳的成熟,从而增强花粉的适应性[71];木质素作为植物细胞壁的重要组成部分,其转运机制还未可知,Alejandro等[72]研究发现AtABCG29参与拟南芥中木质素合成,填补了人们对木质素生物合成理解的关键空白,这可能为木质素工程开辟新的方向。
此外,ABCG还具有抗菌方面的作用,如白花丹叶烟草(Nicotianaplumbaginifolia)中NpPDR1/NpABC1 通过参与次级代谢物的分泌,在防御真菌病原体中起作用[78-79];烟草NtPDR1通过介导抗真菌二萜的外排,在植物的化学防御中发挥作用[80],这为植物抗菌方面的进一步研究奠定了基础。
2.5 其余亚族
除亚族 B、C、G研究较多外,其他亚家族的报道相对较少,它们是否具有与其他亚族相似的功能未有明确的报道。根据模式作物拟南芥的研究,推测蔬菜作物番茄ABCD可能参与长链脂肪酸的过氧化物酶体输入,ABCE可能在核糖体生物发生、翻译控制和基因沉默调节中发挥作用,ABCI在果实发育期表达,表明其可能在果实发育中有特定功能[11],拟南芥ABCF 亚家族成员 AtABCF3 参与根系生长发育[82];水稻OsABCI7可能涉及到多种叶绿体发育调控网络[83],具体功能还有待验证。
3 总结与展望
综上发现,植物ABC蛋白家族体系庞大,成员众多,这些 ABC 蛋白存在于各类细胞器中,大多定位于质膜和液泡膜、还有些定位于线粒体、叶绿体、内质网等细胞器上,其转运底物形式多样,有激素、离子、重金属、代谢产物等,通过介导底物的转运及平衡,随时调控植物的生理状态。就目前研究可知,ABCB参与生长素及重金属转运,ABCC参与转运代谢产物(花青素、植酸、叶酸等物质)、还具有重金属解毒功能,ABCG功能较多,参与代谢产物和激素转运、植物器官形成和细胞发育、逆境胁迫、化学防御等过程,ABCA报道较少,初步表明其与脂质代谢有关,其余亚族功能还有待进一步研究。总而言之,ABC是一类值得深入研究的转运蛋白,研究其可解析生理机制,研究其可辅助植物育种。通过总结上述进展的同时,也发现以下不足。
研究作物种类单一:目前研究主要集中在模式作物和个别作物中,如蔬菜仅有番茄ABC 转运蛋白进行了全基因组分析[11]。
研究深度不够:大多研究只对某些成员做了初步的鉴定和表达,其转运底物尚不明确,具体功能还有待验证[11,23]。
与应用研究结合不紧密:如品质育种已成为研究热点,代谢产物是影响果实品质的关键因素[84],而ABC家族中有许多成员参与转运代谢产物,但未见到关于此类问题的研究报道。
基于上述研究现状,笔者认为未来研究,需从以下方向着手:(1)开展蛋白互作,挖掘未知功能。对目前功能未知的其他亚族建议开展蛋白互作研究,可有助于加快ABC转运蛋白的研究进程;(2)横向研究,由此及彼。利用已有模式作物的研究基础,加快其他作物ABC蛋白的研究,如拟南芥AtABCA9过量表达可促进种子三酰甘油 (TAG)的生物合成,从而增加油籽脂质产量,这是一种源库调控效应,若应用于其他油料作物,可能有助于提高其品质;(3)深度研究,明确机制,加以应用。针对目标蛋白加深研究,掌握其生理功能,并延伸至实践应用,如NpPDR1/NpABC1在抗菌方面起作用, AtATH14 和 AtATH15响应盐胁迫,MsABCG1响应干旱,若加以深度研究,则可应用于植物抗逆育种;(4)多学科联合,准确解析具体功能。由于其结构的相似性,有许多蛋白功能冗余[27-31],大多研究只是通过模式作物进行推测,应通过代谢组学、转录组学、生物信息学及结构生物学等多学科联合,准确具体的鉴定其功能。
猜你喜欢
江苏农业科学(2022年17期)2022-09-24
江苏农业科学(2022年16期)2022-08-26
四川大学学报(自然科学版)(2021年4期)2021-07-15
科学(2020年2期)2020-08-24
中国农业科学(2020年5期)2020-03-30
山东化工(2019年18期)2019-10-16
科技资讯(2018年16期)2018-10-26
科技资讯(2017年12期)2017-06-09
上海农业学报(2017年3期)2017-04-10
红领巾·探索(2015年9期)2015-09-10
