
22份百脉根种质资源表型数量性状的遗传多样性分析
2023-02-10 03:12:14谢文辉赵文武王雷挺赵丽丽王普昶
草地学报 2023年1期
谢文辉,赵文武,王雷挺,赵丽丽*,王普昶
(1.贵州大学动物科学学院,贵州 贵阳 550025;2.贵州师范大学生命科学学院,贵州 贵阳 550025)
百脉根(Lotuscorniculatus)是豆科百脉根属多年生草本植物,原产于欧亚大陆温带地区[1],在我国广泛分布于华东、华中、西南和西北地区[2]。百脉根具有很高的经济效益与生态效益,它的茎叶营养含量丰富、粗蛋白含量高、富含缩合单宁、适口性好,是牛、马、兔等家畜的优良牧草,也是人工草地建植和放牧草地改良的优良草种[3]。百脉根中的缩合单宁能够促进反刍动物的消化[4-5],这使得其与多数豆科牧草不同,不易造成家畜患膨胀病[6]。此外,百脉根根系发达,侧根众多,茎枝匍匐生长,枝叶茂密,覆盖度大,具有改土肥田、防止水土流失,美化环境的作用[7]。
自上世纪80年代以来,我国陆续从国外引进了一些百脉根栽培品种[8],生产试验表明这些品种有较强的适应性和较高的生产性能,广泛种植于我国西北河西走廊灌溉地区和新疆沙漠绿洲[9]。在我国西南岩溶地区,目前关于百脉根的品种选育研究还不系统,百脉根新品种较少。通过引进新的优良百脉根种质资源,培育适宜不同生态区和用途的百脉根品种,这对百脉根在我国西南岩溶山区的规模化种植及当地生态畜牧业发展具有重要的现实意义。
种质资源的遗传多样性分析是新品种选育的基础,通过表型性状检测遗传多样性是最直接且最简易的方法[10]。表型性状是作物基因和外部环境综合作用的结果,张本瑜等人[11]将69份俄罗斯百脉根种质资源划分为3大类群,并发现这69份材料各具特点,差异明显,类型广泛。郝裕辉等人[12]分析了参试的29份无芒雀麦(Bromusinermis)种质资源表型性状差异,发现无芒雀麦种质间遗传变异较大,是环境因子和遗传物质共同作用的结果,育种潜力巨大。此外,前人对箭筈豌豆(Viciasativa)[13]、苜蓿(Medicagosativa)[14]、胡枝子(Lespedeza)[15]等作物表型性状存在的变异也进行了广泛深入研究,为不同牧草种质资源的收集、保护和遗传改良提供了参考。本研究基于表型性状对来自国内外的22份百脉根种质材料的遗传多样性进行分析和比较,拟为优异百脉根种质资源的创新利用和新品种选育提供初步参考。
1 材料与方法
1.1 试验材料
试验材料为22份百脉根种质种子(表1),其中21份由中国农业科学院草原研究所国家种质资源牧草中期库提供,1份由贵州众志恒生态科技有限公司提供。
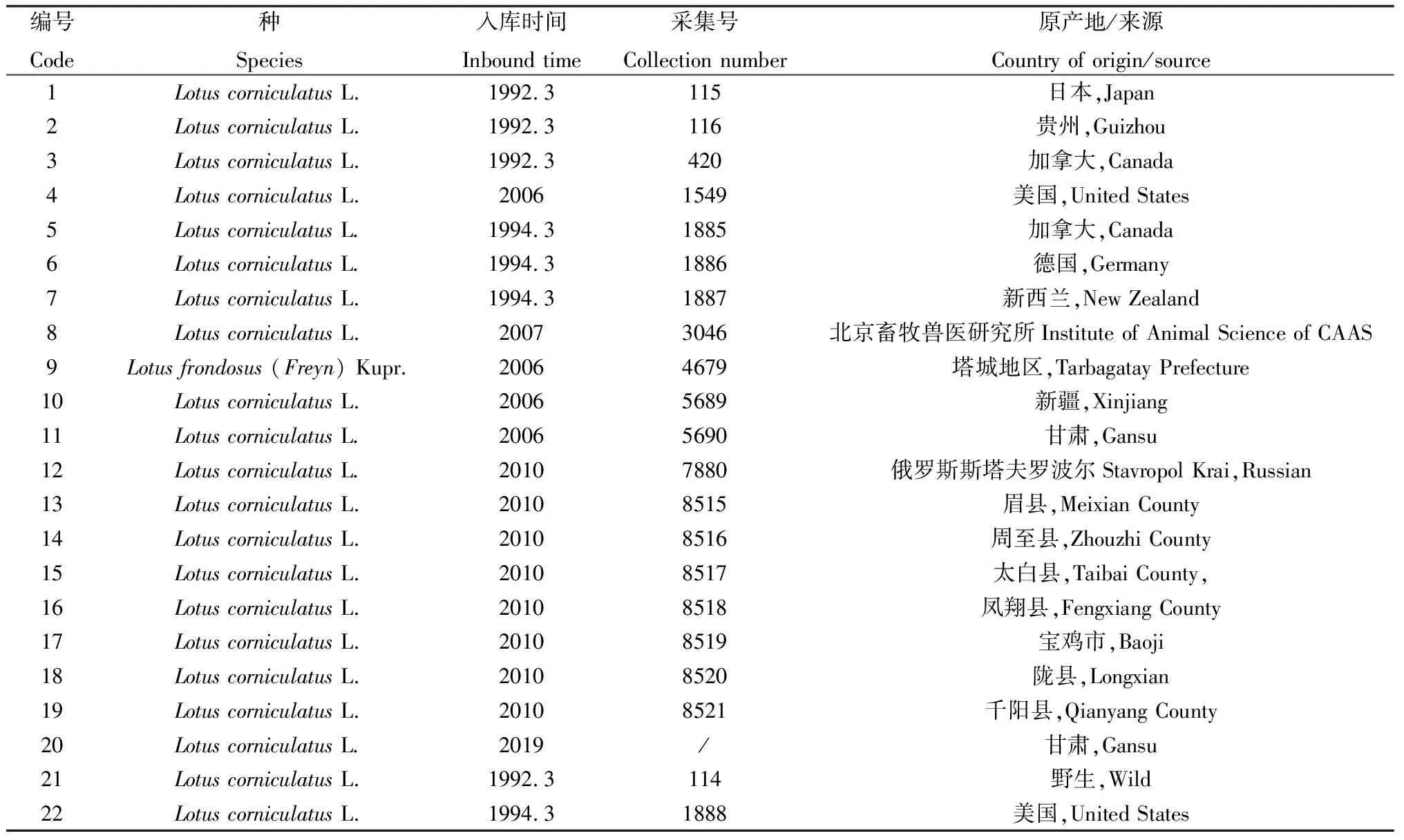
表1 材料名称及来源
1.2 试验方法
1.2.1试验设计与种植 测定完种子表型性状后,先在培养皿上育苗,出苗10 d后,选取长势一致的幼苗移栽至花盆(高30 cm,长35 cm,宽20 cm),每份种质种植3盆,每盆5株。温室适应生长2周后(光照16 h,黑暗8 h,生长温度(22±2)℃),移至室外培养,定期浇水并除草,所有材料均可以完成全部生育期并开花结种。种子形态性状测定包括种荚长,种荚宽,种荚厚,种子周长,种子宽,种子长和种子长宽比;植株表型性状测定包括分枝数,茎长,株高和冠幅,共11个指标。
1.2.2表型数量性状的测定方法 种子形态:使用光学平板扫描仪(V800 PHOTO,Epson,日本)对每份种质种子(每份种子100粒,3次重复)进行扫描,获得种质的种子图像,图像分辨率为300 dpi,像素为1024×1024。使用数字图像分析系统(WinFOLIA 2014,加拿大)对种子图像进行分析,得到种子长、种子宽、长宽比、种子周长。
种荚形态:使用数显游标卡尺(AIRAI SR44,精度0.01 mm,山东青岛)测量种荚长度、种荚宽度、种荚厚度。
在利用直尺(量程为30 cm)测量同一时间不同种质百脉根的茎长、冠幅、株高及记录百脉根的分枝数。其中,茎长:用直尺测量茎顶端至基部的总长度。冠幅:分枝直径的最大值。株高:用直尺测量从地上茎部分到植株花序顶端的自然高度;分枝数:初花期统计并记录百脉根根颈部的一级分枝数。
1.3 数据统计与分析
利用Microsoft Excel 2016对试验数据进行整理,并基于单因素(One-Way ANONA)方差分析的最小显著差(LSD)多重比较法。利用Origin2021软件的模式分析,包括聚类和主成分分析(Principal component analysis,PCA),控制数据质量并以图像方式直观展示本试验研究中22份种质表型数量性状的差异变化情况,选用基于平方欧氏距离的最长距离法对22份种质的植株表型及种子表型性状指标进行聚类分析,为了消除量纲对聚类关系的影响,种质性状数据通过z值标准化后进行聚类,其中,z值标准化计算公式如下:

各表型数量性状之间的相关性和主成分分析在IBM SPSS Statistics 21中分析,其中,基于主成分提取结果,我们构建了1个新的变量y(y1,y2,y3,y4),通过因子分析来反映主成分分析提取到的主成分的得分:
y=∑cx
(1)
公式(1)中,c为特征向量,特征向量为(相应载荷因子/特征根的开方),x为标准化后的种质表型性状值。随后,将提取到的主成分的方差贡献率C作为系数,结合公式(1)构建综合评价模型:
F=C1y1+C2y2+C3y3+C4y4
(2)
最终以评价得分F为基准对各百脉根种质进行综合评价[16]。
2 结果与分析
2.1 百脉根种质资源形态特征多样性
22份百脉根种质表型性状结果表明(表2),除种宽外,22份百脉根种质间各性状均存在显著(P<0.05)或极显著(P<0.01)差异。其中,株高、茎长、分枝数及冠幅等百脉根形态指标的变异系数均超过50%,株高变异幅度最大,为69.48%,种子和种荚性状(种子长、种子宽、种长宽比、种子周长、种荚长、种荚宽、种荚厚)的变异系数较小,在12.11%和20.81%之间。此外,除种子宽外,22份百脉根种质间的种长宽比和种子周长性状存在显著差异(P<0.05),其余各性状(株高、茎长、分枝数、冠幅、种子长、种荚长、种荚宽、种荚厚)均存在极显著差异(P<0.01)。
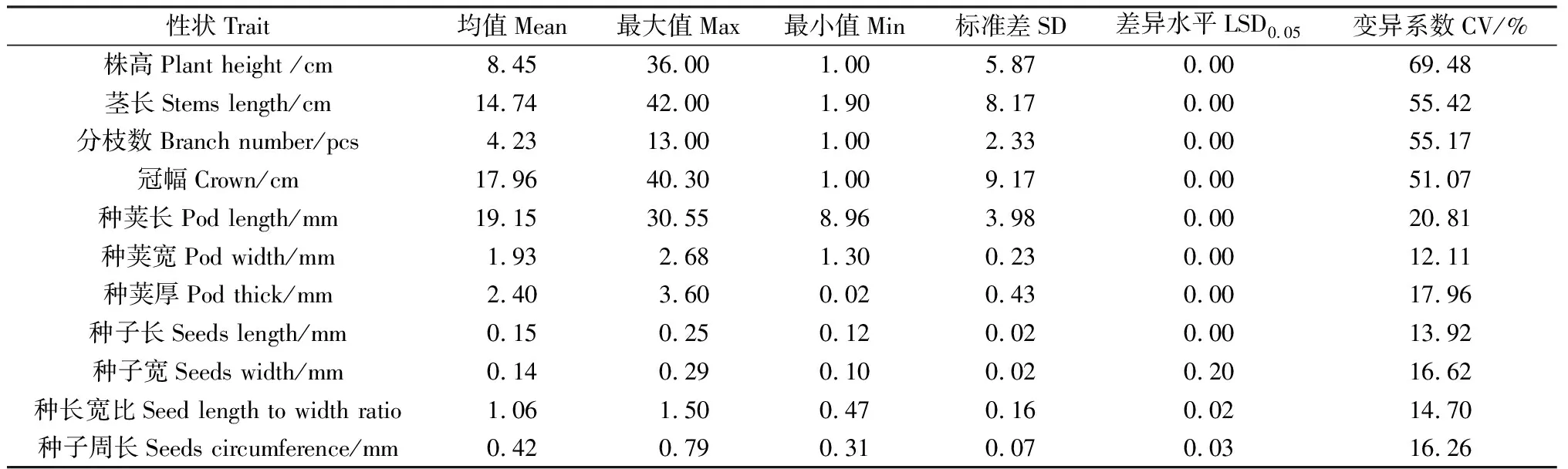
表2 22份百脉根种质表型数量性状变异
2.2 百脉根种质材料不同性状间的相关性分析
对22份百脉根种质材料的11个表型性状进行相关性分析(图1),结果显示分枝数与茎长、株高和冠幅性状之间呈极显著正相关(P<0.01),而分枝数与种荚长呈显著负相关(P<0.05),与种荚宽和种荚厚性状之间呈极显著负相关(P<0.01)。茎长与种荚厚和种子长呈显著负相关(P<0.05)。种荚长与种荚宽和种荚厚之间呈极显著正相关(P<0.01),种子长,种子宽,种周长之间呈极显著正相关(P<0.01)。种子长宽比和种子长之间呈极显著正相关(P<0.01),而与种子宽之间呈极显著负相关(P<0.01)。此外,种荚厚,种子长均与茎长呈显著负相关(P<0.05)。总体而言,不同性状间存在显著相关性。
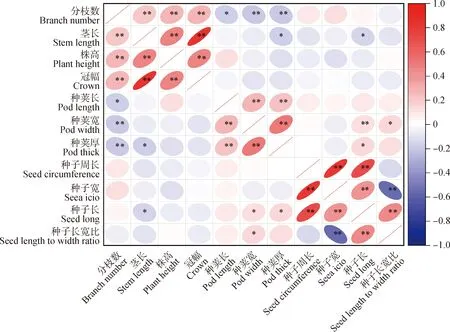
图1 22份百脉根种质不同性状间的相关性分析
2.3 22份百脉根种质资源的聚类分析
通过聚类分析,将22份百脉根种质资源分成3个大的类群(图2)。第I类群仅包含1份种质,种质编号为21,从热图可以看出,该野生百脉根种质的茎分枝数、茎长、株高和冠幅直径高于其它类群种质资源,但该类群种质种子小也是一个主要表型特征,这表明类群I种质的种子小但植株高大,地上生物量高。
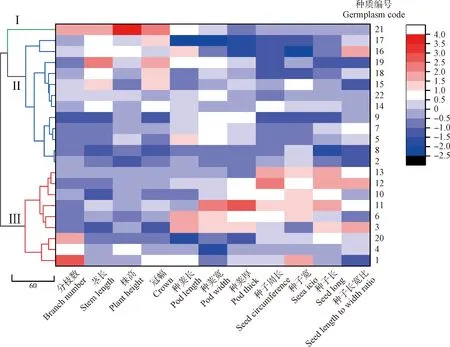
图2 基于表型性状数据的22份百脉根种质材料聚类分析热图
第II类群包含12份种质,种质编号为17,16,19,18,15,22,14,9,7,5,8和2,包含除眉县的所有陕西种质(14,15,16,17,18,19)以及部分国外种质(5,7,22),该类群种质整体表现为种荚和种子小,植株矮小,地上生物量低。第Ⅲ类群包含9份种质资源,种质编号为13,12,10,11,6,3,20,4和1,包含来自甘肃的全部2份种质(11,20)及部分国外种质(12,6,3,4,1)。与I、II类群不同,虽然类群Ⅲ种质的种荚及种子整体大于其它2个类群,但该类群种质整体表现为植株矮小,地上生物量低。
2.4 主成分综合分析及评价
本研究首先对22份百脉根种质的11个性状指标进行了KMO(Kaiser-Meyer-Olkin)检验和Bartlett's球形检验,结果显示,KMO值=0.519>0.5,Bartlett's检验的P值小于0.001,拒绝零假设,因此可以进行主成分分析。
以特征值>1为标准,共提取到4个主成分(表3),累积贡献率达73.181%,这表明这4个主成分能够代表11个表型性状的大部分信息。其中,第1主成分特征值为2.590,贡献率为23.549%,载荷较大的因子包括茎长(-0.752)、株高(-0.581)、冠幅(-0.708)、种子周长(0.516)及种子长(0.554)。第2主成分特征值为2.304,贡献率为20.946%,载荷较大的因子包括种子周长(0.807)和种子宽(0.852)。第3主成分特征值为1.791,贡献率为16.282%,载荷较大的因子包括种荚长(0.500)、种荚宽(0.570)、种荚厚(0.505)及种子长宽比(0.527)。第4主成分特征值为1.364,贡献率为12.404%,载荷较大的因子包括种子长宽比(0.732)。

表3 主成分分析结果
随后,我们将提取的4个主成分的方差贡献率作为系数,结合公式(1)构建了综合评价模型:F=0.23549y1+0.20946y2+0.16282y3+0.12404y4,并以评价得分为基准对各百脉根种质进行综合评价,评价结果显示(表4),11和12号种质的得分最高,综合表现整体优良。
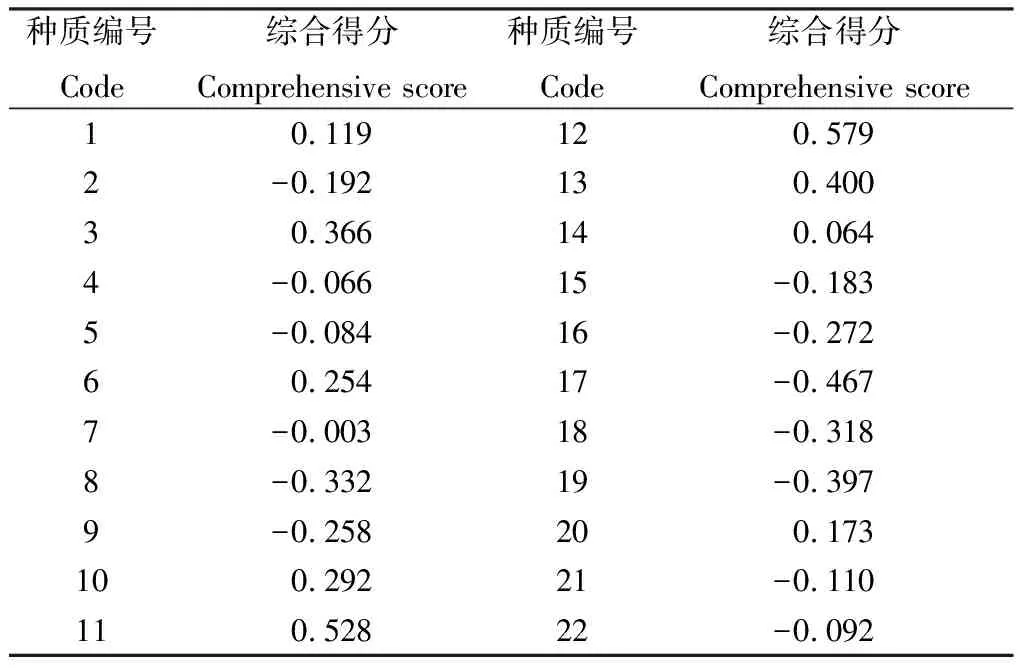
表4 基于主成分分析的22份百脉根种质资源综合得分
3 讨论
种质资源表型性状是植物基因型和环境互相影响的综合体现[17],因此具有相对稳定且丰富的表型变异特征[18]。对优质牧草百脉根种质资源的表型性状进行遗传多样性分析,是百脉根种质资源研究的重要工作[19],也是筛选特异牧草资源作为育种材料的前提[20]。本研究结果表明,除种宽外,22份百脉根种质间各性状均存在显著(P<0.05)的表型性状差异,这些表型性状的变异表明在各种质内存在潜在可用的遗传变异。变异系数(Coefficient of variation,CV)反映不同种质特征性状的变异程度,CV值越大,说明不同种质间相应的特征性状的差异越大,之前的研究表明,当CV达到10%以上时,表明所选种质材料间有显著差异[21-22],这与本研究结果一致。此外,本研究中的22份百脉根种质的CV值均大于50%,可能是因为22份种质来自全球不同国家和地区,海拔,气候等环境差别较大,而环境条件是种质资源遗传变异的重要因素之一[23]。
22份百脉根种质表型性状相关性分析表明,种子长宽比与种子周长间无显著相关性,这与对小麦(Triticumaestivum)、青稞(Hordeumvulgare)的研究结果相似[24]。之前的研究表明,种子长宽比主要用来描述种子形状,而与种子大小性状无关[15],结合本研究,百脉根的种子大小与种子形状这两个数量性状可能也为独立遗传。百脉根种质的分枝数与茎长,株高和冠幅性状间呈正相关,而这些表型性状都是构成株型的重要因素,本世纪以来,包括玉米、水稻和小麦在内的众多作物在株型育种中已经取得了较大的进展[25-27]。对于牧草而言,合理的株型能保证牧草在生长过程中利用良好的生长姿势进行光合作用,进而提升产量[28]。本研究中,百脉根的株型性状如株高、分枝数,都属于数量性状,应考虑各株型性状间的相关性从而更全面的研究株型形成的机制,来改良百脉根株型性状以提高其产量和品质。
聚类分析是植物育种的必要手段之一[29],应用聚类热图分析可以直观呈现多个种质的多个表型性状值的全局大小变化,同时反映不同种质资源间的遗传差异和各类群的特点。本研究中。通过聚类热图分析,将22份百脉根种质划分为3个具有明显差异的类群,第I类群包含的21号种质资源,与其它种质资源具有较远的亲缘关系,该种质株高和冠幅均大于其它种质。第II类群包含了除眉县外的所有来自陕西省的种质及部分国外种质,表明来自同一生态区的百脉根种质表型性状的差异不明显,这也表明,尽管不同种质间的性状特征和地理来源间可能存在一定的相关性,但地理来源并不对表型性状起决定作用[12]。类群Ⅲ种质的种荚及种子随整体大于其它2个类群,但该类群种质整体表现为植株矮小,之前的研究表明,大粒种子的萌发率高于小粒种子,因为相比小粒种子,大粒种子能够储存丰富的营养物质及更完整的酶系统,进而促进种子更快萌发[30],但种子大小对植株形态特征的影响较小,可能是因为幼苗生长至营养物质的积累阶段时,大粒种子的优势不再显著[31-32]。
由于百脉根种质材料表性状指标众多,且各指标间存在很强的相关性。因此,需要通过提取主成分,以降维的方法将指标重新组合,将多个性状指标转化为较少的几个相关性不强的综合指标,从而达到综合和简化的目的[33],该方法在燕麦(Avenasativa)[34],披碱草(Elymusdahuricus)[35]等植物种质资源的遗传多样性评价中广泛运用。本研究中,11个联系相关的指标经主成分分析,转化为4个主成分,累计贡献率达到73.181%,包含了大部分性状信息,茎长、株高、冠幅、种子周长、种子长、种子宽和种子长宽比的主成分载荷较高,是造成百脉根种质资源表型差异的主要因素。在牧草育种研究中应充分利用主成分因子,有效选择育种材料,以缩短育种周期[36]。基于主成分分析的综合评价显示,聚类分析后的第Ⅲ类群的11和12号种质主成分综合得分最高,是2份综合表现整体优良的种质材料,依据这些信息,在育种工作中,可根据实际用途或育种目标从中再进一步选择,为选择优良的育种亲本或栽培材料提供参考,提高育种效率[37]。
4 结论
本研究已初步证明,参试的22份百脉根种质资源遗传变异较大,11个表型数量性状间具有广泛相关性,各指标在各种质之间多样性较丰富,通过聚类分析将22份种质分为3类,基于主成分综合得分筛选出2份综合表现优良的材料11和12号,并且这2份材料均聚类为同一类,具有较大的种荚和种子,可作为选育种子高产品种的目标亲本加以利用。本研究可为百脉根的实际应用提供理论参考。
猜你喜欢
东北林业大学学报(2023年6期)2023-05-31 02:56:16
丽水学院学报(2022年2期)2022-04-19 01:22:10
山东农业科学(2021年8期)2021-09-12 09:43:50
林业科技情报(2021年3期)2021-09-01 02:01:26
草业科学(2019年11期)2019-12-12 07:31:52
江苏农业科学(2019年5期)2019-09-02 14:01:46
海南师范大学学报(自然科学版)(2017年4期)2018-01-22 07:50:52
中南林业调查规划(2017年1期)2017-12-19 00:58:07
广东农业科学(2017年5期)2017-08-29 10:37:31
应用海洋学学报(2014年1期)2014-11-22 07:17:44
