
狗牙根小尺度克隆多样性及空间遗传结构研究
2023-02-10 03:12:08苟甜甜马艳娜吉轶楠臧国长郑轶琦
草地学报 2023年1期
苟甜甜,马艳娜,吉轶楠,臧国长,郑轶琦
(河南科技大学园艺与植物保护学院,河南 洛阳 471000)
“克隆植物”是指在自然条件下能自发地利用根、匍匐茎或横生根茎等器官产生具有潜在的独立性个体的植物[1]。在高等植物中,克隆植物占到了35%~80%,并且它们在许多生态系统中处于优势地位[2]。克隆多样性是指一个克隆植物种群基株的数量水平,能够反映该克隆种群的有性繁殖程度以及初始建群种的数量。近年来,国内外学者已经对糙叶苔草(Carexscabrifolia)[3]、剑叶金鸡菊(Coreopsislanceolata)[4]、海菖蒲(Enhalusacoroides)[5]、穗状狐尾藻(Myriophyllumspicatum)[6]、羊草(Leymuschinensis)[7]、竹叶眼子菜(Potamogetonmalaianus)[8]等植物进行了克隆多样性研究,表明不同植物间克隆多样性存在着较大差异。
空间遗传结构(Spatial genetic structure,SGS)[9],是物种繁育机制、繁殖体的散播方式、克隆繁殖模式、环境异质性等因素共同作用的结果,它反映了植物种群适应环境的能力[10],有助于理解不同干扰因子对植物种群自然更新、遗传格局以及种间关系的影响作用[11-12],因此对克隆植物空间遗传结构的研究具有重要的意义。李钧敏等[13]发现3个不同斑块蛇莓(Duchesneaindica)的小尺度克隆结构差异明显,是因为其受到环境、干扰、突变等因素的强烈影响造成的。左威等[14]发现在较为单一的生境内,活血丹(Glechomalongituba)采取克隆繁殖的模式,其空间遗传结构较强;而当生境较为复杂时,活血丹采取种子繁殖的模式,其空间遗传结构较弱。Rico等[15]研究了放牧对石竹(Dianthuscarthusianorum)空间遗传结构的影响,发现与未放牧草原上的种群相比,轮流放羊的种群显示出较弱的空间遗传结构。Ohsako利用7个微卫星位点,研究了克隆植物筛草(Carexkobomugi)的空间遗传结构,发现在小距离范围内筛草的空间遗传结构显著正相关,克隆繁殖在一定程度上对空间遗传结构有很大贡献[16]。因此,开展对克隆植物在不同生境条件下空间遗传结构的研究,有助于阐明该克隆植物空间遗传结构的形成机制。
狗牙根(Cynodondactylon)隶属于禾本科(Gramineae),自然条件下以营养繁殖为主[17]。由于其抗逆性强,生命力旺盛,适应性广,匍匐茎发达,侵占能力强,不仅能运用于园林绿化、固土护坡还可以作为牧草[18-19]。我国狗牙根种质资源丰富,主要分布在我国南方地区,另外华北省份、新疆、青海等地也有分布[20],其种内变异程度很大[21]。目前,对狗牙根的克隆多样性和空间遗传结构等方面研究较少。马艳娜等[22]对采自不同生境的狗牙根进行了克隆构型和生物量的研究,发现狗牙根种群的克隆构型分为“密集型”和“游击型”。孙宗玖等[17]对新疆地区的狗牙根无性系构件进行了研究比较,由于原始生境的多样性,导致狗牙根的表型性状表现出较大的可塑性。赵玉等[23]发现狗牙根的无性系构件的持续更新主要依靠根茎芽来实现。由于环境异质性会导致克隆植物的无性系构件,为了生存和发展,调整其生殖策略导致克隆多样性的差异,从而形成不同的空间遗传结构,因此本研究通过SSR分子标记对3个不同生境的狗牙根进行了克隆多样性和空间遗传结构的分析,旨在探讨各种干扰和生境异质性对其克隆多样性及空间遗传结构的影响,阐明狗牙根在不同生境中的生长繁衍机制,了解狗牙根对环境的适应机理。
1 材料与方法
1.1 取样方法
在河南省唐河县境内采集了131份狗牙根材料。样地概况参见马艳娜等[22]的报道,具体生境详见表1。所选样地的狗牙根种群都呈带状分布,在每个样地的中间位置设置1个160 cm×80 cm的矩形样方(图1),每个样方都被划分为20 cm×20 cm的正方形网格,在横纵线的交点处,选取长势良好,无病虫害植株的叶片,编号并记录每个样本的空间坐标,将采集的叶片放入装有变色硅胶的塑封袋中保存备用。
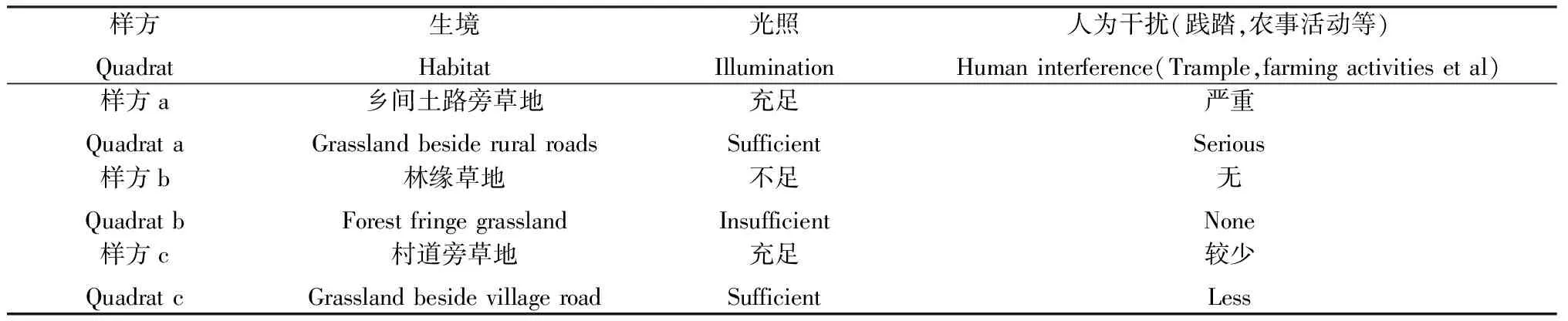
表1 3个狗牙根样方的生境概况
1.2 总DNA的提取与检测
采用高效植物DNA提取试剂盒(DP350-02,北京天根生化科技有限公司)提取DNA。所提取DNA的质量用1%的琼脂糖凝胶电泳进行检测,DNA的纯度和浓度利用紫外分光光度仪(T6新世纪,北京普析通用仪器有限责任公司)进行检测,并将DNA溶液稀释至50 ng·μL-1,放置-20℃保存备用。从本实验室前期已经筛选的狗牙根SSR引物中,选择多态性丰富、重复性较好、条带清晰的9对引物(表2)进行分析,引物序列、扩增反应体系以及扩增程序参考Wang等[24]的报道。

表2 本研究所采用的SSR引物
1.3 数据分析
根据产物在电泳图谱中的位置,有带记为“1”,无带记为“0”,得到0/l矩阵。使用R软件‘Polysat’程序包‘assignClones’功能对数据进行分析,设置阈值,阈值范围内的个体被分配到同一克隆组[25]。遗传多样性等参数如Nei’s遗传多样性指数(h)、Shannon’s信息指数(I)、遗传分化系数(Gst)以及基因流(Nm)使用POPGENE 1.32 软件[26]获得。
克隆多样性水平通过基株总数(G)、平均克隆大小(NC)、基因型比率(PD)、基因型分布的均匀度Fager指数(E)、Simpson多样性指数(D)这5个参数[27]进行估算。克隆分布范围通过计算基株数(G)、分株数(ni)、同一克隆内分株间的最大距离(lmax)、最小距离(lmin)、总距离(L)以及平均距离(M)进行分析,其中同一克隆内分株间的总距离为各分株的直线距离之和,平均距离M=L/ni[28]。利用GenAlEx 6.5[29]计算3个样方中每个分株的遗传距离矩阵,计算空间自相关系数r,并对遗传距离和地理距离进行Mantel检验[30]。利用SPAGeDi 1.2软件[31]计算Sp值以进一步量化群体空间结构的水平。参考Erichsen等[32]的报道计算3个样方的混交度(Mi)。
2 结果与分析
2.1 SSR扩增结果与遗传多样性
利用9对SSR引物对采自3个样方的131份狗牙根材料进行扩增,结果见表3。各遗传参数表明供试狗牙根材料的遗传多样性较为丰富。遗传分化系数Gst为48.47%,表明48.47%的遗传变异来自样方间,51.53%来自样方内部,样方内的遗传分化大于样方之间。由Gst估算的狗牙根的基因流较小,仅为0.53。
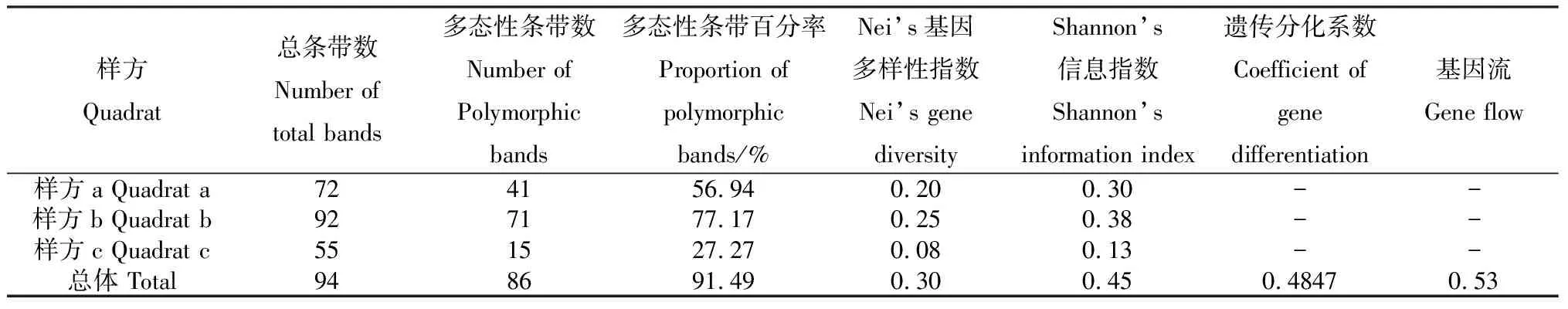
表3 SSR扩增结果及遗传多样性
2.2 克隆多样性与克隆大小
3个样方的克隆空间分布情况如图1所示。结果显示3个样方都是由多克隆组成,共鉴别出46个基株,其中样方a共检测到17个基株,样方b检测到22个,样方c检测到7个。狗牙根各样方的克隆多样性结果见表4,从各项指标反应,供试的3个样方中,克隆多样性水平由高到低依次为:样方b>样方a>样方c。
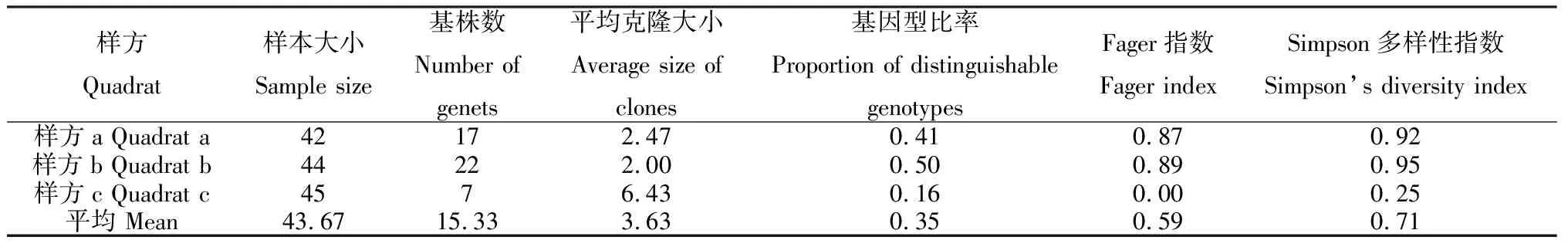
表4 狗牙根3个样方的克隆多样性
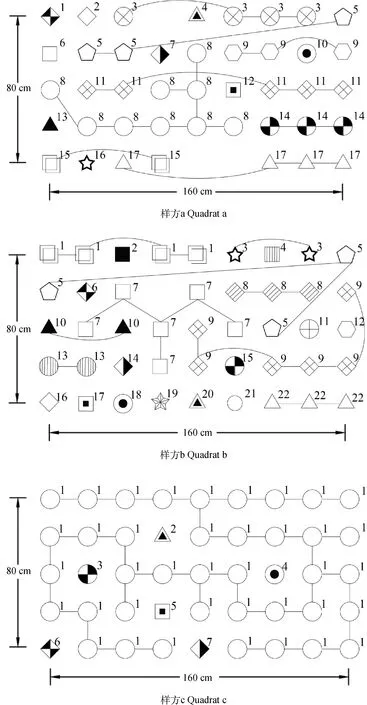
图1 狗牙根3个样方的个体基因型及克隆空间分布
46个基株通过克隆繁殖共形成了84个分株(表5)。分株间的总距离为2340.59 cm,分株间的最大距离为178.89 cm,最小距离为20.00 cm,平均距离为37.10 cm。样方a的最大克隆有8个分株,分株总数为25,克隆大小为3.13,同一克隆内分株间的最大和最小距离分别为141.42 cm和20 cm,总距离的范围40.00~161.42 cm,平均距离的范围为18.54~80.71 cm。样方b的最大克隆有5个分株,分株总数为22,克隆大小为2.44,同一克隆内分株间的最大和最小距离分别为161.25 cm和20.00 cm,总距离的范围20.00~217.82 cm,平均距离的范围为20.00~108.91 cm。样方c的最大克隆、分株总数、克隆大小均为38,同一克隆内分株间的最大和最小距离分别为178.89 cm和20 cm,总距离为760 cm,平均距离为20 cm。
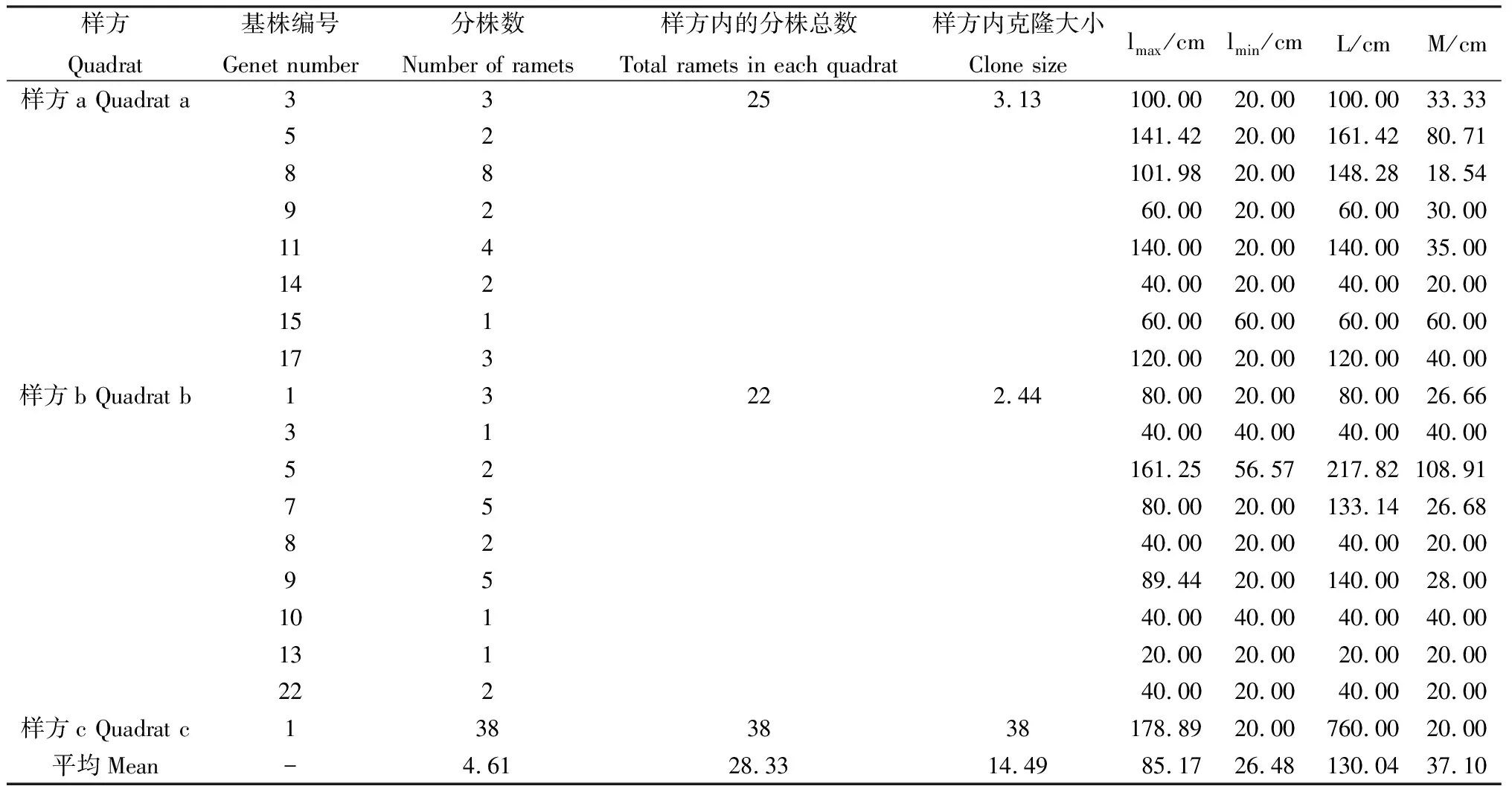
表5 狗牙根3个样方内的克隆分株数和分株间的距离
2.4 空间遗传结构分析
狗牙根3个样方的空间自相关分析结果如图2所示。样方a在空间距离小于20 cm时存在显著的正空间遗传结构,在20 cm处空间自相关系数最大(r=0.24),X-轴截距为47.17 cm。样方b在空间距离小于50 cm时存在显著的空间遗传结构,在40 cm处出现最大空间自相关系数(r=0.22),X-轴截距为66.27 cm,在空间距离大于50 cm时不存在显著的空间遗传结构。样方c在所研究的距离等级下不存在显著的空间遗传结构,X-轴截距为15.26 cm。Mantel检测表明样方b(r=0.15,P=0.01)的遗传距离和空间距离之间存在显著正相关性,而样方a(r=0.01,P=0.36)和c(r=-0.04,P=0.21)的遗传距离和空间距离之间均不相关。
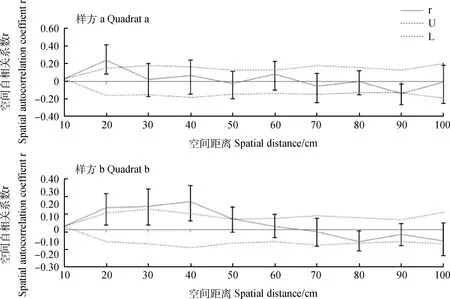
图2 3个样方狗牙根的空间自相关分析
利用SPAGeDi软件进一步量化各样方的SGS水平,Sp值表明样方b的空间遗传结构最强(Sp=0.13),样方c的空间遗传结构次之(Sp=0.12),而样方a的空间遗传结构最弱(Sp=0.06)。
2.5 混交度分析
3个狗牙根样方混交度在0.37~0.81之间,表明3个样方中的克隆分布差异较大。样方a的混交度为0.67,呈现中度混交结构状态,样方b的混交度为0.81呈现出强度混交结构,说明样方a,b狗牙根的分株间竞争激烈,内异质性很强,为游击性策略。样方c的混交度为0.37,基株混杂程度较低,边缘明显,为密集型策略。
3 讨论
3.1 克隆多样性分析
本研究结果表明,供试狗牙根的克隆多样性水平高于蛇莓[13](PD=0.20,D=0.64),也高于Ellstrand等[27]总结的克隆植物的平均值(PD=0.17,D=0.62),说明供试狗牙根克隆多样性水平较高。供试的3个样方中,样方b的克隆多样性水平最高(PD=0.50,D=0.95),样方a略低(PD=0.41,D=0.92),样方c远低于前2个样方(PD=0.16,D=0.25)。生境异质性、基因流、体细胞突变、繁育方式、取样策略等会影响植物的克隆多样性水平[13,27]。样方a的狗牙根生长在乡间小路上,有行人和车辆的践踏,样方b位于林缘草地,遮荫较严重,这2个样方的狗牙根生长环境受到较多的干扰,狗牙根分配更多的资源给有性繁殖,以逃避不良的生存环境,因此样方内的基株数目较多,克隆多样性和基因型比例较高。样方c阳光充足,且无人为干扰,生存环境相对稳定,因此该样方的狗牙根克隆繁殖比例较高,基株数目少,克隆多样性较低。Brzosko等[33-34]的研究也表明在干扰生境中,有性繁殖占据优势,有利于克隆多样性的提高;而在相对稳定的生境中,植物将更多的繁殖资源分配给无性繁殖,其克隆多样性水平越低。此外,在3个狗牙根样方中均存在单个分株组成的克隆,这可能是由于体细胞突变造成的,从而使该植物能够更好的适应环境[35],李钧敏等[13]对蛇莓克隆结构的研究中也存在单株组成克隆的现象。
3.2 空间遗传结构分析
本研究对狗牙根3个不同生境的空间自相关分析,表明a样方和b样方分别在20 cm和50 cm处存在显著性正相关。狗牙根为典型的克隆植物,无性繁殖可以使相同基因型的个体聚集,导致短距离内呈现正的空间自相关,从而对植物种群的空间遗传结构产生显著的影响[36-38]。此外,本研究中狗牙根的空间遗传结构形成距离低于活血丹,左威等[14]认为活血丹较大的空间遗传结构形成距离可能与其较高的种子萌发率有关,狗牙根种子的萌发率约为48%[39],远低于活血丹(79.3%),可能导致其空间遗传结构的形成距离较小。3个样方狗牙根的Sp值存在差异,b(Sp=0.13)、c(Sp=0.12)样方的狗牙根存在较高水平的空间遗传结构,而样方a(Sp=0.06)的空间遗传结构较弱。小尺度空间遗传结构存在与否以及空间遗传结构的强度大小与物种分布的生境、基因流以及外界干扰状况密切相关[40-41]。样方c光照、养分等资源充足,不存在任何干扰,克隆繁殖占据优势,克隆分株呈聚集分布,从而导致小尺度空间遗传结构较强[42-43],样方b较高的空间遗传结构可能是因为种子传播受限导致的,在四叶重楼(Parisquadrifolia)空间遗传结构的研究中也发现种子传播受限导致了其较强的空间遗传结构[44],而样方a由于人类和动物的频繁活动,导致种子的有效传播距离增加,从而削弱了其空间遗传结构的强度。
3.3 混交度分析
混交度反映了基因型之间的聚集程度,如果各基因型个体在样方里各自聚集,不同基因型之间没有交集,边缘明显,混交度低,则为密集型生长策略;如果各基因型个体相互混杂,混交度高,则为游击型生长策略[45]。Erichsen等[32]利用9对微卫星标记对小叶椴(Tiliacordata)克隆结构进行了研究,其混交度为0.48~0.58,居群内不同的基因型处于混合状态,并采取游击型的生长策略。本研究中a,b样方处于中强度混交结构(Mi>0.5),说明样方内基因型混杂程度较高,这是因为2个样方的生境均受到了不同程度的干扰,如遮荫、车辆碾压以及人类践踏,狗牙根通过克隆整合,呈现出游击型生长策略,样方内的狗牙根表现出强烈的觅食反应[1],该生长策略便于对分散分布资源的获取和利用,从而占据更多的生存空间,获取更多的资源。样方c的混交度较低,同一基因型聚集较多,这是因为样方c光照充足,并靠近农田,环境条件较好,狗牙根呈现出密集型的生长策略[1],该生长策略对获取集中分布的资源占优势。
4 结论
本研究应用9对SSR引物分析了3个不同生境小尺度格局狗牙根的克隆多样性和空间遗传结构,发现供试3个样方的狗牙根材料克隆多样性较高,空间遗传结构与繁育方式、人为干扰以及生境异质性密切相关,同时,狗牙根可采取不同的克隆构型来应对异质性生境。因此,本研究对了解狗牙根进化潜力、生态适应性机制以及生态稳定性的维持能力等方面具有十分重要的意义。
猜你喜欢
林业勘查设计(2022年1期)2022-02-15 05:35:12
乡村科技(2021年17期)2021-10-20 08:50:56
安顺学院学报(2021年4期)2021-09-16 08:11:08
落叶果树(2021年6期)2021-02-12 01:29:10
林业调查规划(2020年3期)2020-06-03 07:02:36
体育科技(2016年2期)2016-02-28 17:06:07
中国当代医药(2015年20期)2015-03-01 02:04:33
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:17
西南军医(2015年1期)2015-01-22 09:08:26
中外医疗(2013年23期)2013-08-21 01:45:16
