
六斑异瓢虫生物学特性及其对核桃扁叶甲幼虫的捕食能力
2023-02-06 07:41温学达刘玉升
昆虫学报 2023年12期
温学达,刘玉升
(山东农业大学植物保护学院环境昆虫研究所, 泰安 271000)
核桃扁叶甲Gastrolinadepressa,又名核桃叶甲,属鞘翅目(Coleoptera)叶甲科(Chrysomelidae)扁叶甲属Gastrolina。主要危害核桃Juglansregia、核桃楸Juglansmandshurica、枫杨Pterocaryastenoptera(孟庆英, 2006)。该虫分布范围广泛,在四川、云南、黑龙江、辽宁、山东、江苏等地均有发生(虞佩玉等, 1996),以成虫和幼虫取食叶肉,造成叶片网状或缺刻,有时可将全叶食光,仅剩叶脉(孟庆英等, 2008)。长期以来对核桃扁叶甲的防治主要以化学农药为主(李飞等, 2021),但是大量使用化学杀虫剂会对环境和天敌产生负面影响,导致生态系统失衡,天敌控制力下降(Shahetal., 2020; Ghongade and Sangha, 2021)。因此控制该虫的非化学措施逐渐受到重视,充分利用捕食性天敌的控害作用成为主要措施之一。
有益瓢虫是世界重要的天敌资源之一,在害虫防治中起着重要作用(包建中和古德祥, 1998)。六斑异瓢虫Aiolocariahexaspilota,又名奇变瓢虫,属鞘翅目(Coleoptera)瓢虫科(Coccinellidae)(孟祥杰等, 2008)。栖息于多种阔叶树,有时在松树、竹林等植物上也可发现其踪迹。国内主要分布于黑龙江、吉林、内蒙古、甘肃、陕西、北京、河北、河南、山西、湖北、四川、台湾、福建、贵州、云南、西藏;国外主要分布于日本、朝鲜、俄罗斯、印度、尼泊尔、锡金、缅甸(虞国跃, 2010)。六斑异瓢虫喜欢捕食小型昆虫,是叶甲等农业害虫的重要捕食性天敌昆虫,具有体型大、食量大、捕食专化性强的特征,对赤杨叶甲Agelasticacoerulea(林锦枫, 1977)、榆紫叶甲Ambrostomaquadriimpressum(杜文梅, 2017)、漆树叶甲Podontialutea(李连虎和王维, 1984)、杨树叶甲Chrysomelapopuli(曲友, 2012)等,以及漆条跳甲Podontialutea、白杨叶蝉C.popali、柳十星叶蝉C.
liginigpunetata(杜文梅等, 2014)有较强的捕食能力。
目前关于六斑异瓢虫的研究报道相对较少。为更加全面地了解该瓢虫资源、探究其生物防治潜力,本研究对其生物学特性和其对核桃扁叶甲2龄幼虫的捕食能力进行了较系统的观察,以期为今后六斑异瓢虫的识别鉴定、规模化繁殖及生物防治的实施提供一定的指导和借鉴意义。
1 材料与方法
1.1 供试昆虫
核桃扁叶甲幼虫采自山东省泰安市泰山区山东农业大学植物保护学院实验站(36°10′N, 117°09′E);六斑异瓢虫于2022年4月27-29日采自泰安市泰山区药乡林场(36°19′N, 117°07′E),饲养于山东农业大学昆虫资源与环境生物研究室。
1.2 主要仪器
体视显微镜(上海缔伦光学仪器有限公司,型号:XTL-207A);人工气候箱(合肥右科仪器设备有限公司,型号:RGC-500A)。
1.3 六斑异瓢虫生活史调查
试验点位于山东省泰安市泰山区药乡林场。2021年4月至2022年3月,每隔10 d在林间调查1次,观察林间六斑异瓢虫的卵、幼虫、蛹、成虫的分布情况。
1.4 六斑异瓢虫形态特征及生活习性观察
在室内条件下,六斑异瓢虫幼虫用玻璃培养皿(d=10.5 cm)以单头单皿的方式饲养,放入浸湿的脱脂棉保湿,扣紧上盖,防止逃逸(饲养1和2龄幼虫的玻璃培养皿需用保鲜膜封口,并用针在膜上扎多个小孔)。六斑异瓢虫成虫用塑料养虫盒(长×宽×高=16.0 cm×11.0 cm×7.0 cm)饲养,每个塑料养虫盒放入10头成虫(♀∶♂=1∶1)。每日投放核桃扁叶甲1, 2和3龄幼虫,保持饵料充足,达理论捕食量的1.2倍。及时更换湿润棉球、清理饵料尸体、虫粪、残枝叶等杂物,保持容器的洁净和干燥。六斑异瓢虫成虫出现交尾现象后,将该对成虫转移至单独玻璃培养皿中饲养,将滤纸折叠成波浪形铺在玻璃培养皿底部,便于记录、转移卵粒。使用体视显微镜观察记录六斑异瓢虫的形态特征。随机抽取六斑异瓢虫不同龄期的幼虫各20头,使用50分度游标卡尺测量幼虫的头宽和体长。在温度(25±0.5) ℃、相对湿度(RH) 80%±3%和光周期14L∶10D的人工气候箱内,观察记录六斑异瓢虫的产卵、孵化、蜕皮、化蛹、羽化等生活习性。
1.5 以核桃扁叶甲幼虫为食料的六斑异瓢虫的发育历期和存活率观测
试验在人工气候箱(条件同1.4节)中进行。以核桃扁叶甲幼虫(1, 2和3龄)作为饵料,将5对六斑异瓢虫雌雄成虫分别接入玻璃培养皿中饲养。六斑异瓢虫产卵后移去成虫,每天观察3次卵的孵化情况。卵孵化后,将六斑异瓢虫1龄幼虫移入新的玻璃培养皿中,一皿1头,每天更换含有核桃扁叶甲幼虫(1和 2龄)的新鲜核桃叶片作为饵料,每隔12 h观察1次,记录六斑异瓢虫的发育历期及存活情况,每个处理5次重复。
1.6 六斑异瓢虫对核桃扁叶甲2龄幼虫的捕食量测定
分别测定六斑异瓢虫2, 3和4龄幼虫及成虫对核桃扁叶甲2龄幼虫的日捕食量。选择核桃扁叶甲2龄幼虫作为供试猎物,按不同密度梯度接种在盛有新鲜核桃叶的玻璃培养皿内,六斑异瓢虫2龄和3龄幼虫按10, 15, 20, 25和30头/皿接种核桃扁叶甲2龄幼虫;六斑异瓢虫4龄幼虫和成虫按20, 30, 40, 50, 60和70头/皿接种核桃扁叶甲2龄幼虫。将饥饿12 h的六斑异瓢虫2和3龄幼虫及饥饿24 h的六斑异瓢虫4龄幼虫和成虫分别接入上述玻璃培养皿中,每皿1头。随后将玻璃培养皿置于人工气候箱(条件同1.4节)。24 h后检查每个玻璃培养皿中残余的核桃扁叶甲2龄幼虫数量,并计算六斑异瓢虫的捕食量(捕食量=初始核桃扁叶甲2龄幼虫数量-存活核桃扁叶甲2龄幼虫数量)。以只放入核桃扁叶甲2龄幼虫而不放入六斑异瓢虫的玻璃培养皿为空白对照校正实际的捕食量(实际的捕食量=观测捕食量-空白对照死亡量)。每个处理5次重复。
1.7 六斑异瓢虫对核桃扁叶甲2龄幼虫的捕食功能反应试验

1.8 六斑异瓢虫对核桃扁叶甲2龄幼虫的寻找效应分析

1.9 数据分析
实验数据采用Microsoft Excel 2010处理后,利用SPSS 23.0软件进行卡方检验来验证不同处理的六斑异瓢虫对核桃扁叶甲2龄幼虫的取食是否拟合Holling Ⅱ型功能反应模型。采用Duncan氏新复极差检验在显著性水平0.05下比较分析六斑异瓢虫2, 3和4龄幼虫及成虫在不同处理下捕食量的差异显著性。
2 结果
2.1 六斑异瓢虫形态特征
成虫形态特征:体长8.04~11.52 mm,体宽7.03~10.04 mm;体广卵形,圆弧形拱起,体背无毛。前胸背板基色黑色,两侧各有1块白色或浅黄色大斑,外缘有细窄黑边;头黑色,触角褐色,共11节,复眼黑色,近于圆形;小盾片黑色;胸部腹面黑色;腹部腹面黑色,外缘黄褐色,腹板刻点稀疏(图1);雌虫第6腹板后缘弧形突出,雄虫第6腹板后缘平截内凹。

图1 六斑异瓢虫成虫腹面特征Fig. 1 Ventral characteristics of Aiolocaria hexaspilota adults
成虫鞘翅色斑变化较大。鞘翅基色主要为橙红色或黄褐色,斑纹为黑色。鞘缝、 基缘、外缘都具有黑色条纹,外缘黑色条纹较细。通过调查,发现该地区有两种常见的鞘翅色斑类型。第1种鞘翅色斑类型表现为:鞘翅缝纵黑纹较狭窄,黑色斑纹单个分开;肩斑位于肩胛突上,向下伸展,达中线的1/4处向内弯曲成短柄;亚肩斑位于肩斑之后的外缘中部,向内后弯曲成短柄;中斑为较小的近圆形黑斑,位于中线的1/2处且稍偏于外方(图2: A)。第2种鞘翅色斑类型表现为:翅缝纵黑纹宽阔,黑色斑纹毗连成带;肩斑、亚肩斑与第1种类型相似;中斑位于鞘翅的中部稍前且稍偏于鞘缝;端斑位于端部之前且靠近鞘缝;肩斑外缘有一细窄黑色纵纹向下延伸与中斑、端斑顺次相连,形成黑色纵带;亚肩斑、中斑连成横带与鞘翅纵缝黑纹相交(图2: B)。

图2 山东省泰安市两种常见的六斑异瓢虫色斑类型Fig. 2 Two common types of color variations of Aiolocaria hexaspilota in Tai′an City, Shandong ProvinceA: 第1种色斑类型 The first type of color variation; B: 第2种色斑类型 The second type of color variation.
卵形态特征:长卵圆形,橙红色,长1.72~2.03 mm,宽0.71~0.84 mm。
幼虫形态特征:纺锤形,共4龄。1龄初孵幼虫体深褐色,头、足均为黑色,腹面黄褐色; 2龄幼虫前胸背板呈橙红色,侧缘凸起,有1对半圆形黑斑,中、后胸背板各有1对较小黑斑,第1-8腹节各有6个黑色毛疣,横向排列; 3龄幼虫前胸背板前缘中间有一橙红色细线(图3: F); 4龄幼虫前胸背板外缘长有白毛,中间橙红色线变宽,腹部两侧有针状突起(图3: G)。1-4龄幼虫头宽和体长见表1。
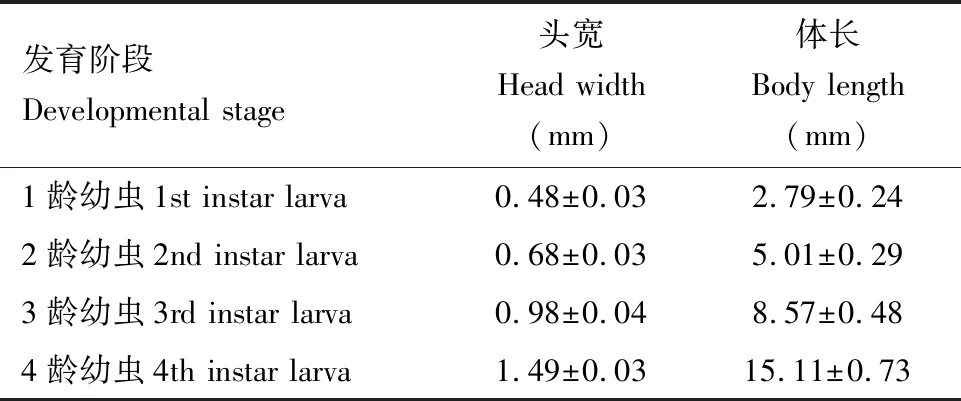
表1 六斑异瓢虫各龄幼虫头宽和体长Table 1 Head width and body length of Aiolocaria hexaspilota at different larval instars

图3 六斑异瓢虫各发育阶段形态特征及生活习性Fig. 3 Morphological characteristics and life habits of Aiolocaria hexaspilota at different developmental stagesA: 成虫交尾Adult mating; B: 卵块Egg mass; C: 1龄幼虫捕食Predation of the 1st instar larva; D: 蜕皮Exuviation; E: 2龄幼虫捕食Predation of the 2nd instar larva; F: 3龄幼虫3rd instar larva; G: 4龄幼虫4th instar larva; H: 蛹Pupa; I: 初羽化成虫Newly emerged adult; J: 成虫捕食Adult predation.
蛹形态特征: 裸蛹,卵圆形,深黄褐色,有黑色斑纹(图3: H);长7.03~10.02 mm,宽6.01~7.50 mm; 背面拱起,腹面前半平扁、后半弯曲。
2.2 六斑异瓢虫生活习性
2.2.1交配及产卵: 越冬成虫取食1~3 d后开始交尾(图3: A)。全天均可交配,无明显交配高峰。最短交尾时间约2 min,最长超10 h,交尾过程中雌虫可继续取食,受较大惊吓时停止交配或雌虫背负雄虫逃离。交尾后十多小时产卵。有隐蔽处产卵的习性,一般产于叶片或滤纸背面。卵竖立成行排列或呈块状(图3: B)。单雌每次产卵1~54粒,一生产卵量约800粒,产卵期约40~60 d。
2.2.2取食及行为: 六斑异瓢虫取食核桃扁叶甲的卵和幼虫,一般不取食核桃扁叶甲成虫,食物缺少时会尝试攻击其腹部。初孵幼虫即可吮食核桃扁叶甲1和2龄幼虫(图3: C)。用口器咬住核桃扁叶甲幼虫躯体,吸取体液,使其干瘪而死(图3: E)。成虫行动敏捷,有假死性;受惊吓时,脚关节缝隙分泌难闻的红黄色液体。易发生自残行为,在饥饿状态下,成虫会取食幼虫和卵,高龄幼虫取食低龄幼虫和卵,同龄幼虫之间也会出现相互咬食的现象,3和 4龄幼虫因种内残杀导致的死亡率最高。
2.2.3孵化及蜕皮: 卵由橙红色逐渐变为深褐色,临近孵化呈黑色。初孵幼虫咬破卵壳后缓慢伸展四肢,经约2 h完全爬出卵壳。初孵幼虫聚集在卵壳周围,取食卵壳,约20 h后分散取食。蜕皮时,胸部背板出现裂缝,幼虫胸部背板逐渐显现,头和四肢缓慢脱出。初蜕皮幼虫头、足均为淡黄色半透明状(图3: D)。
2.2.4化蛹及羽化: 老熟幼虫预蛹历期约1 d,体收缩、颜色变淡,以腹部末端足突将身体固定。全天均可羽化,羽化时蛹体摆动,蛹背裂开,成虫从裂缝中爬出。初羽化成虫行动迟缓,体橙红色,膜翅下垂,约1 h后收缩到鞘翅下(图3: I)。3~6 h后鞘翅斑纹逐渐显现。在蛹壳附近停息1 d左右前往别处捕食(图3: J)。
2.3 六斑异瓢虫生活史
六斑异瓢虫在泰安地区一年发生1代。4月上旬越冬成虫开始活动觅食,4月下旬至5月中旬为产卵盛期,5月上旬至6月上旬为孵化盛期,5月下旬至6月下旬为化蛹盛期,6月上旬至7月中旬为羽化盛期。10月份之后成虫逐渐停止进食,开始向越冬场所迁飞或在原处枯叶下进入越冬期(表2)。

表2 山东省泰安市六斑异瓢虫生活史Table 2 Life history of Aiolocaria hexaspilota in Tai′an City, Shandong Province
2.4 以核桃扁叶甲2龄幼虫为食料的六斑异瓢虫发育历期和存活率
如表3所示,在温度(25±0.5) ℃,RH 80%±3%,光周期14L∶10D的条件下,测得以核桃扁叶甲2龄幼虫为食的六斑异瓢虫的卵、 1-4龄幼虫、预蛹和蛹的平均历期分别为3.84, 2.24, 1.90, 2.85, 4.42, 1.04和5.68 d。除预蛹的存活率较低外,其余各发育阶段六斑异瓢虫的存活率均在93%以上, 3龄幼虫存活率最高(96.67%)。
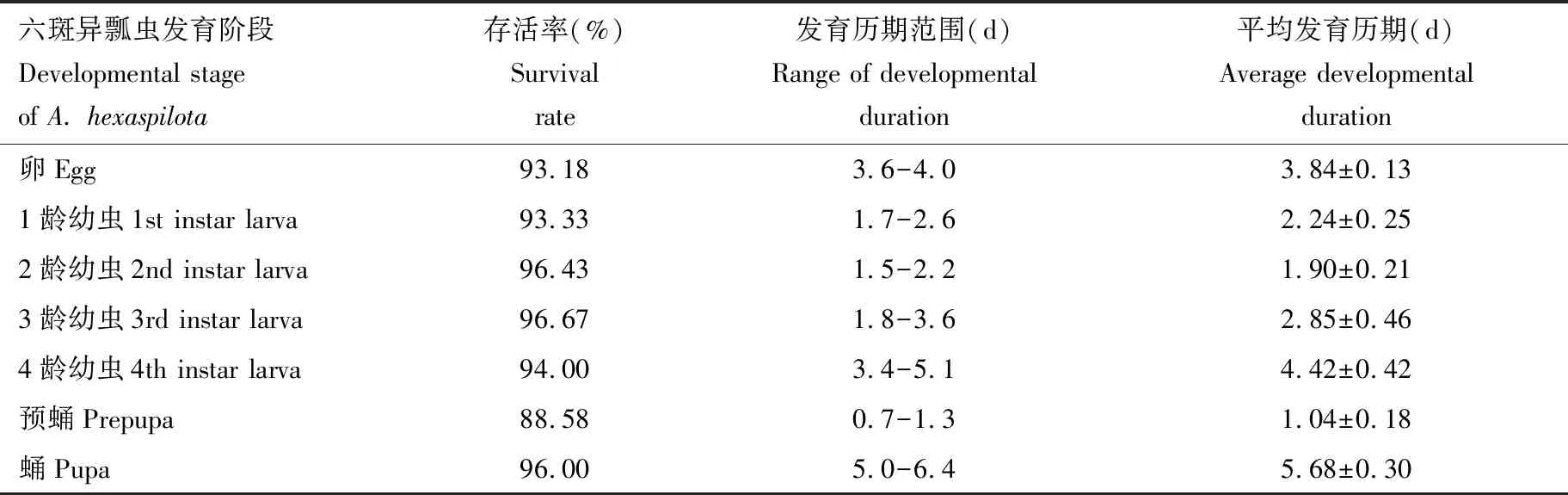
表3 25 ℃下不同发育阶段六斑异瓢虫取食核桃扁叶甲2龄幼虫时的历期及存活率Table 3 Duration and survival rates of different developmental stages of Aiolocaria hexaspilota fed on the 2nd instar larvae of Gastrolina depressa at 25 ℃
2.5 六斑异瓢虫在核桃扁叶甲2龄幼虫不同密度下的日捕食量
六斑异瓢虫2-4龄幼虫及成虫的日捕食量见表4,结果表明:在设定的核桃扁叶甲2龄幼虫密度范围内,六斑异瓢虫2-4龄幼虫及成虫的单头日捕食量均随核桃扁叶甲2龄幼虫密度增加而增加,捕食量增长速度呈逐渐减小趋势。
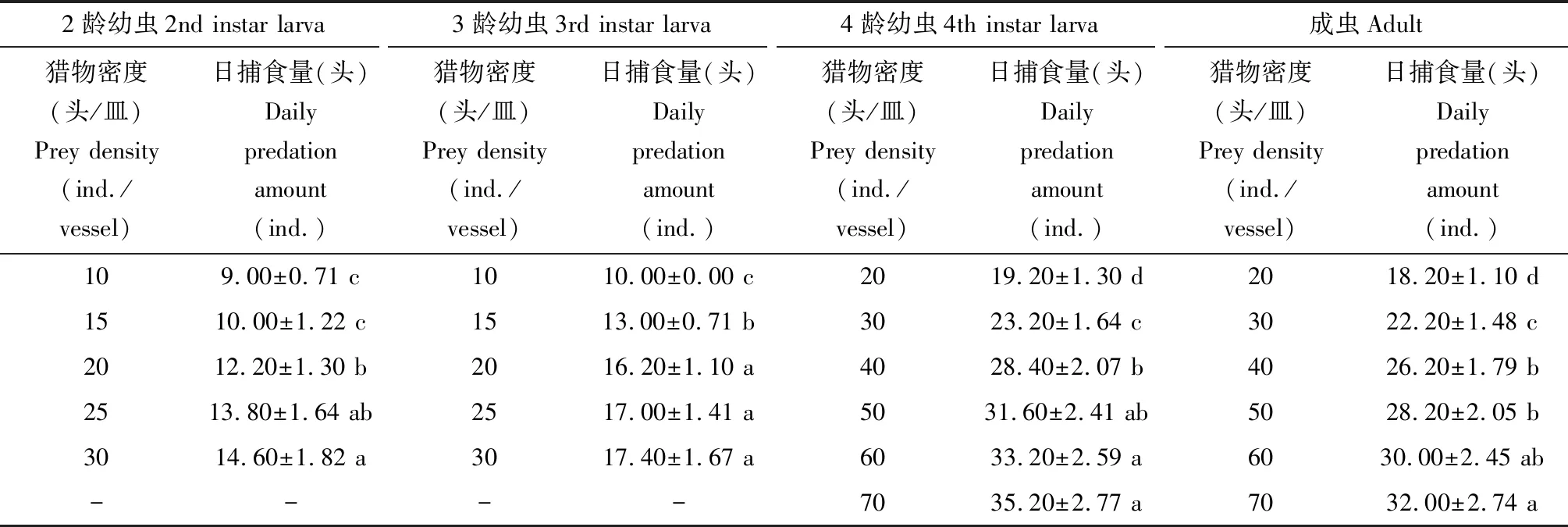
表4 不同发育阶段六斑异瓢虫对不同密度核桃扁叶甲2龄幼虫的单头日捕食量Table 4 Daily predation amount per individual of Aiolocaria hexaspilota at different developmental stages on the 2nd instar larvae of Gastrolina depressa at different densities
2.6 六斑异瓢虫对核桃扁叶甲2龄幼虫的捕食功能反应
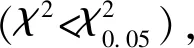
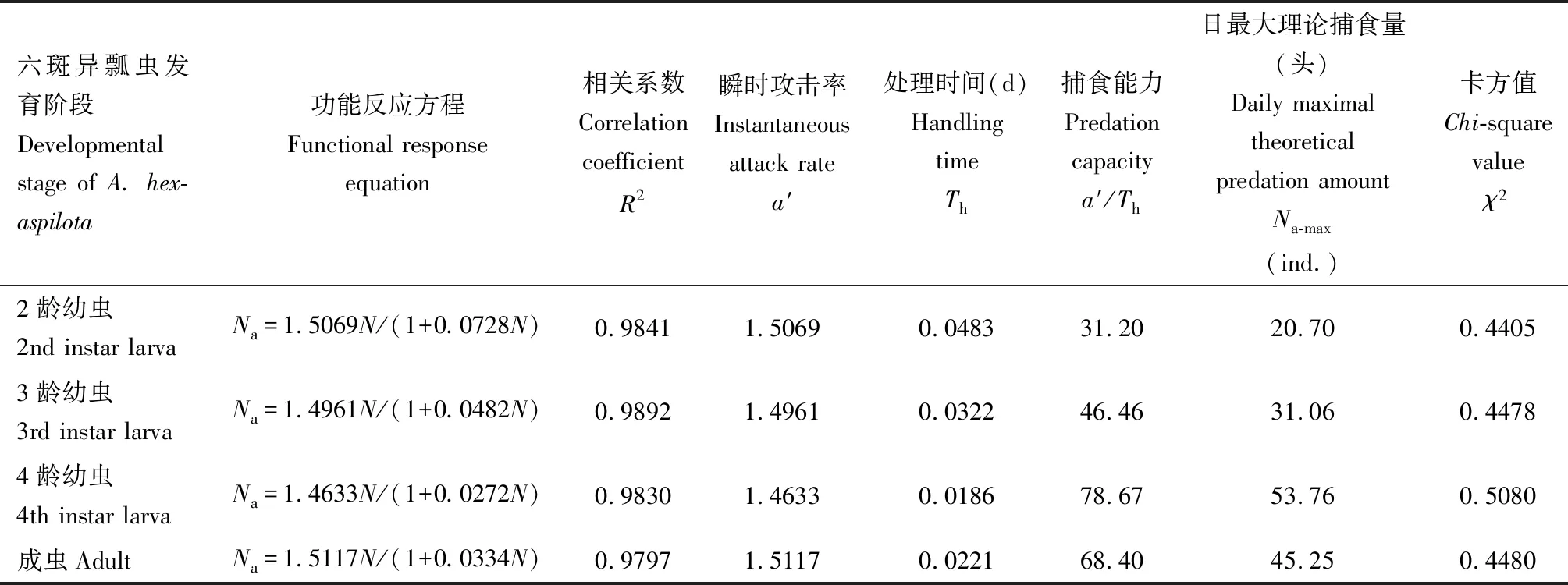
表5 不同发育阶段六斑异瓢虫对核桃扁叶甲2龄幼虫的捕食功能反应Table 5 Predation functional responses of Aiolocaria hexaspilota at different developmental stages to the 2nd instar larvae of Gastrolina depressa

图4 不同发育阶段六斑异瓢虫对核桃扁叶甲2龄幼虫的捕食功能反应拟合曲线Fig. 4 Fitting curves of the functional responses of Aiolocaria hexaspilota at different developmental stages to the 2nd instar larvae of Gastrolina depressaA: 2龄幼虫2nd instar larva; B: 3龄幼虫3rd instar larva; C: 4龄幼虫 4th instar larva; D: 成虫Adult.
2.7 不同龄期六斑异瓢虫对核桃扁叶甲2龄幼虫的寻找效应
Holling(1959)认为寻找效应必须依赖于寄主的种群密度,随着猎物密度的增加,寻找时间减少,捕食率提高。根据表5中的相关参数值及寻找效应公式,可计算出六斑异瓢虫2-4龄幼虫及成虫对核桃扁叶甲2龄幼虫的寻找效应。从图5可见,寻找效应与其猎物核桃扁叶甲2龄幼虫的密度呈负相关关系。六斑异瓢虫2-4龄幼虫及成虫对核桃扁叶甲2龄幼虫的寻找效应随着猎物密度的增加而降低。六斑异瓢虫4龄幼虫与成虫对核桃扁叶甲幼虫的寻找效应比3和2龄幼虫的寻找效应高。
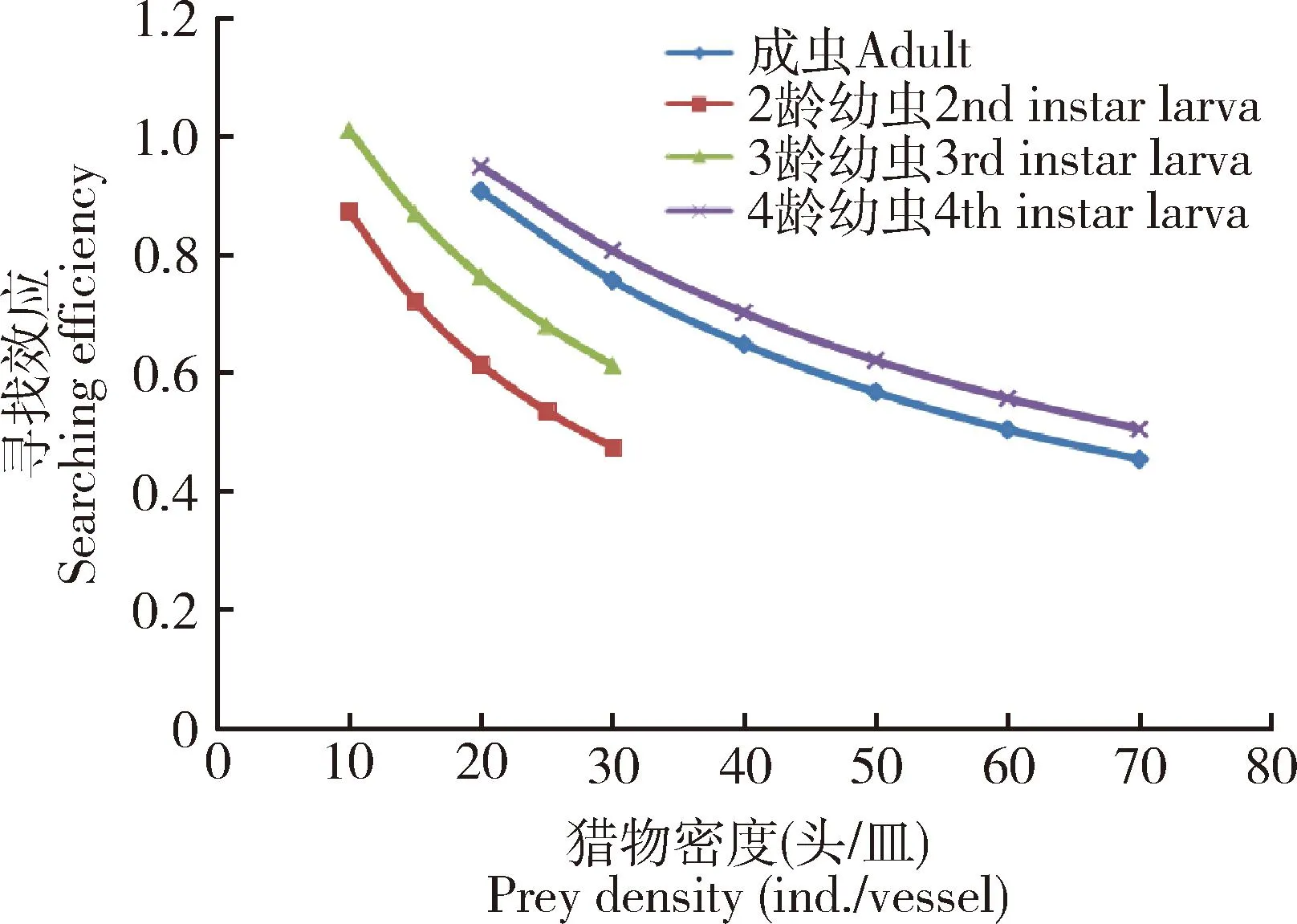
图5 不同发育阶段六斑异瓢虫对核桃扁叶甲2龄幼虫的寻找效应Fig. 5 Searching efficiency of Aiolocaria hexaspilota at different developmental stages to the 2nd instar larvae of Gastrolina depressa寻找效应
3 讨论
在山东省泰安市泰山区药乡林场,六斑异瓢虫有两种主要的色斑类型(图2)。这与虞国跃(2008)记录的常见六斑异瓢虫色斑类型相一致。但本次调查未发现曾在黑龙江抚远柳树上拍摄到的六斑异瓢虫色斑类型(虞国跃, 2010)。鉴于调查时间、范围有限,可能该地区还存在未被发现的色斑类型,或是因为山东泰安与黑龙江抚远两地区的气候等地理环境因素存在差异,一定程度上使六斑异瓢虫种群的多态结构发生较明显的空间变异。
六斑异瓢虫在各地的发生期有明显差异,整体呈现出由南向北逐渐推迟的现象。在云南省漾濞地区,3月中旬越冬成虫开始活动觅食;4月中旬至5月上旬为产卵盛期;4月下旬至5月中旬为孵化盛期; 5月上旬至5月下旬为化蛹盛期;5月中旬至5月下旬为羽化盛期(杨源, 1983)。在湖北省恩施地区,越冬成虫4月下旬开始活动觅食,交尾、产卵、孵化以及羽化的始期、盛期均比云南省漾濞地区推迟约1个月,且羽化盛期持续时间更长(李连虎和王维, 1984)。在吉林省长春地区,越冬成虫于4月下旬至5月上旬才开始活动(鄢淑琴等, 1994; 曲友, 2012)。但本次调查中,我们在4月上旬发现了六斑异瓢虫越冬成虫的活动迹象(表2),该时间明显早于纬度较低的湖北省恩施地区的记录。其原因可能与两地的海拔、气候等环境因素差异有关。昆虫在生长发育过程中,各生物学参数存在地理差异(王政等, 2020),这种差异是由于昆虫在不同环境下为达到最大适应值所产生的(邹超等, 2017)。
据杜文梅等(2017)报道,在25 ℃条件下,六斑异瓢虫的卵、 1-4龄幼虫和蛹历期分别为: 3.88, 1.74, 1.55, 1.90, 3.51和 5.11 d。本研究同样在25 ℃条件下进行,记录的卵、1-4龄幼虫和蛹历期分别为3.84, 2.24, 1.90, 2.85, 4.42和 5.68 d(表3),这与其报道略有不同。原因可能是六斑异瓢虫取食的猎物以及相对湿度不同所导致。
本研究通过研究六斑异瓢虫对核桃扁叶甲幼虫的功能反应模型、寻找效应模型,明确了六斑异瓢虫对核桃扁叶甲幼虫的控害能力。天敌对猎物的捕食功能反应是衡量天敌捕食能力的重要指标之一,也是评价天敌对害虫控制作用的重要依据(刘树生, 2004)。本研究结果表明,六斑异瓢虫2-4龄幼虫和成虫对核桃扁叶甲2龄幼虫的捕食量均随猎物密度增加而增大,捕食量增长的速率随密度增加而逐渐减小,拟合功能反应符合Holling Ⅱ模型(图4)。类似的结果在六斑异瓢虫对榆紫叶甲卵捕食能力的研究中也有报道(杜文梅, 2014)。天敌对害虫的攻击系数与处理时间之比是衡量天敌作用的参数之一,a′/Th越大,对害虫的控制能力越强(牟吉元等, 1997)。六斑异瓢虫4龄幼虫和成虫对核桃扁叶甲2龄幼虫的a′/Th值分别为78.67和 68.40(表5)。从反映捕食能力指标理论日最大捕食量来看,六斑异瓢虫4龄幼虫的日最大捕食量也高于成虫的(表5)。这说明前者具有较强的捕食能力。但是4龄幼虫生活史较短,因此六斑异瓢虫对核桃扁叶甲幼虫的控制作用主要集中在成虫阶段。
寻找效应是捕食者在捕食过程中对寄主攻击的一种行为效应。六斑异瓢虫对核桃扁叶甲2龄幼虫的寻找效应随猎物密度增加而减弱,当猎物密度相同时,六斑异瓢虫高龄幼虫的寻找效应大于低龄幼虫,成虫仅次于4龄幼虫, 按高低排序为: 4龄幼虫>成虫>3龄幼虫>2龄幼虫(图5)。
自然条件下,瓢虫的捕食行为除了与其自身特性有关外,还与温度、湿度、光照、风力等环境因子、天敌的集团竞争、其他生物的干扰以及活动空间大小等有很大关系(杨帆等, 2016)。本研究是在人工气候培养箱内进行的,属于恒温恒湿的半封闭条件,因此统计的瓢虫捕食量很可能与其在外界复杂多变环境下的捕食量存在一定差异。同时由于条件所限,研究采用的六斑异瓢虫是未分雌雄的混合种群,对雌雄个体各自的捕食功能反应尚有待进一步研究。但本研究仍可为采用核桃扁叶甲幼虫饲养六斑异瓢虫以及利用该瓢虫防控核桃扁叶甲提供借鉴与参考。
猜你喜欢
阅读(科学探秘)(2021年9期)2021-05-30
红蜻蜓·低年级(2020年9期)2020-09-26
考试与评价·高二版(2020年4期)2020-09-10
环境昆虫学报(2020年3期)2020-07-07
小学生作文(低年级适用)(2018年4期)2018-11-29
现代农村科技(2018年8期)2018-08-23
橡塑技术与装备(2018年14期)2018-07-20
金色少年(奇趣科普)(2017年6期)2017-07-25
销售与市场(管理版)(2017年3期)2017-03-28
小朋友·快乐手工(2015年10期)2015-11-02
