
山西省不同地区柴胡根部可培养内生真菌的多样性分析
2023-02-02 13:18申小营刘心雨任广喜刘春生
微生物学杂志 2023年6期
申小营, 姜 丹, 李 萌, 刘心雨, 任广喜, 刘春生
(北京中医药大学 中药学院,北京 102488)
内生真菌是指某一生命阶段或整个生命周期内,生活在植物体内,但不引起植物表现出疾病症状的一类真菌[1]。植物体内蕴含着丰富的内生真菌资源,不同植物体内的优势菌群差异较大。西洋参根部内生真菌优势菌属为一种柔膜菌(Unclassifiedhelotiales)和假裸囊菌属(Pseudogymnoascus)微生物等[2];云南重楼内生真菌以头孢霉属(Cephalosporium)为优势种群,占总数的24.69%[3];黄芩叶和茎的内生真菌优势属为链格孢属(Alternaria),根的内生真菌优势属为镰刀菌属(Fusarium)[4];可以看出不同植物体内及相同植物不同器官的优势内生真菌种群存在显著差异。同时,不同地理-气候环境对植物的内生真菌菌落结构也有明显影响,取样地点和取样组织类型在决定药西瓜中内生真菌的组成起到了重要的作用[5];不同产地杜仲树皮内生真菌的生物多样性不同[6]。环境强烈影响了沙漠中锁阳内生真菌种类的数量和多样性[7]。内生真菌不仅在生态系统发挥着重要作用,还可以产生多种有生理活性的药效成分[8]。马钱内生真菌Exserohilumrostratum次生代谢产物中的化合物(+)-monocerin具有潜在的抗炎活性[9]。狭叶柴胡内生真菌CHS4菌株发酵液的乙醇粗提物对肝癌细胞有抑制作用,内生真菌CHS3可以产生柴胡皂苷[10-11]。近年来植物内生真菌与植物之间的互作关系受到广泛关注,植物内生真菌对植物具有促生、提高抗病害能力等多种有益影响。有学者在金线莲内生真菌中发现两株可以促进金线莲生长发育的促生菌和两株可以拮抗金线莲茎腐病的拮抗菌[12],苹果内生真菌PG06、LG12、LG06、LY09对苹果组培苗均具有显著促生作用[13];野生大豆内生真菌ZX-WD-S4、WDS-6、Z1L-5对大豆苗期生长起到显著的促生作用[14];深色有隔内生真菌OchroconisguangxiensisX22及其多糖和寡聚糖提取物对香蕉苗具有促生与抗枯萎病作用[15]。有关研究表明,野生甘草的质量稳定优于栽培型甘草[16],而野生型甘草中的内生真菌多样性也明显多于栽培甘草[17];由不同内生真菌合成的人工群落可以促进丹参的生长,提升丹参的产量与品质[18]。因此可以得出,内生真菌不但自身可以合成生物活性物质,也能够对药用植物的质量产生一定影响。柴胡(BupleurumchinenseDC.)是一种多年生药用植物,根部入药,其含有的皂苷类、黄酮类、挥发油类、甾醇类等多种活性成分具有广泛的药理活性;2020版《中华人民共和国药典》规定,柴胡皂苷a+d 的含量不得低于0.30%[19]。现代药理研究证明柴胡中的挥发油具有解热作用[20],柴胡中的皂苷类成分具有镇痛和保护肝脏等作用[21-23]。临床上柴胡可用于治疗感冒发热、肝郁气滞、胸肋胀痛、脱肛、子宫脱垂、月经不调等症。柴胡分布地区较广,黑龙江、甘肃、陕西、山西、河北、河南、四川等地均有栽培种植[24],研究发现不同产区的柴胡皂苷含量差异较大[25-27],有学者在测定甘肃、山西、河北、赤城四个产地柴胡的柴胡皂苷a和柴胡皂苷d含量时,发现赤城产地柴胡的柴胡皂苷含量最高,甘肃产地柴胡的柴胡皂苷含量最低[28]。研究表明,海拔、温度、光照等生态因子影响药用植物体内次生代谢产物的积累[29],晋东北大同市浑源县、广灵县与晋中部吕梁市汾阳县显著高于晋南临汾市、长治市、晋城市、运城市的柴胡皂苷 a+d含量,主成分分析表明柴胡皂苷含量与经纬度、海拔呈正相关[30]。北京中医药大学药用植物与分子生药课题组(本课题组)前期研究发现晋南的柴胡挥发性气味比晋北丰富,晋南与晋北柴胡中羧酸、酯类、烯烃、醇类的相对质量分数接近,但晋北醛类相对质量分数高于晋南,晋北酮类、芳香类、烷烃的相对质量分数远远低于晋南[31]。山西省是我国柴胡种植面积和市场影响最大的省份之一[32],山西省的地势狭长,地形多样,海拔落差大,南北的气候具有明显差异,通过对山西省内不同地区的经度、纬度、海拔、年均温、年降水量等地理-气候数据分析发现,山西省北部区、中部区与南部区三个自然地理区的地理-气候因子存在显著性差异[33]。综上所述,地理-气候因子对植物中活性成分和植物中内生真菌的多样性都有不同程度的相关性,植物中内生真菌也是直接或间接影响植物中活性成分的因素之一。那么不同产地柴胡根部内生真菌多样性是否与内生真菌多样性相关?哪些类群的内生真菌可能对柴胡质量产生影响?要想对以上问题进行研究,就需建立丰富的柴胡内生真菌库。因此,本研究通过对不同产地柴胡内生真菌进行分离和鉴定,建立柴胡中内生真菌菌库,研究柴胡根部可培养内生真菌的微生物群落结构,为开发促进柴胡皂苷 a+d 含量积累的内生真菌或产生柴胡皂苷 a+d 的内生真菌奠定基础。
1 材料与方法
1.1 材料
1.1.1 植物材料 综合考虑山西省的地理-气候特性与山西省柴胡栽培基地分布,于2020年10~11月分别从山西省北部与南部地区各采集35批样品,共采集70批次健康柴胡(BupleurumchinenseDC.),样品编号见表1,采集点分布图见图1。

图1 样品采集地分布图(山西省)Fig.1 Distribution of sampling sites (Shanxi Province)
1.1.2 培养基 马铃薯葡萄糖琼脂培养基(PDA):取马铃薯200 g,切成1 cm3小方块,加入适量去离子水煮沸30 min,纱布过滤,向滤液中加入琼脂17~20 g、葡萄糖20 g,加蒸馏水定容至1 L。放入锥形瓶中,121 ℃灭菌30min,倒板备用。
1.1.3 主要试剂与仪器设备 真菌基因组 DNA 提取试剂盒(北京索莱宝科技有限公司);2×TaqMaster Mix(南京诺唯赞生物科技有限公司);ITS1(5′-TCCGTAGGTGAACCTGCGG-3′);ITS4(5′-TCCTCCGCTTATTGATATGC-3′)(北京全式金生物技术股份有限公司)。梯度聚合酶链式反应(PCR)仪(Veriti96,美国 ABI 公司);洁净工作台(SW-CJ-1D,苏州苏洁净化设备有限公司);台式小型离心机(1-14,德国 Sigma 公司);凝胶成像系统(GelDoc,美国 Bio-Rad 公司);水平电泳槽(JY-SP3,北京君意东方电泳设备有限公司);冷冻混合球磨仪(MM400,德国 Retsch 公司);立式压力蒸汽灭菌器(LSB35,江阴滨江医疗设备有限公司)。
1.2 方法
1.2.1 内生真菌的分离 新鲜柴胡洗净后,用流动水冲洗30 min,将根切成2 cm小段,在超净台中进行根部表面消毒,步骤为75%乙醇消毒3 min,用无菌水冲洗1次,5%次氯酸钠消毒5 min,用无菌水冲洗1次,75%乙醇消毒30 s,无菌水冲洗5次。用无菌滤纸吸干柴胡小段表面水分,无菌手术刀将其分割为5 mm组织块,放入PDA平板,每个平板放入4个组织块,每个批次设置10个重复,设置两个空白对照组:一组为组织印记法,将吸干水分的柴胡小段在PDA培养基上来回滚动,印记1 min;另一组为漂洗液检验法,取最后一次漂洗液200 μL涂布于平板上。以上平板均在28 ℃培养箱中避光培养7 d,若空白对照组无真菌菌落生长,则将所得菌落转移至新的PDA平板中进行纯化,否则重复上述步骤。纯化的菌落放入15%的甘油中,-80 ℃保存备用。
1.2.2 内生真菌DNA的提取 在超净台中用无菌竹签将培养皿中的菌丝收集在2 mL离心管中,放入两个无菌小钢珠,液氮速冻离心管,用球磨仪磨碎菌丝,随后使用真菌基因组DNA提取试剂盒提取真菌的基因组DNA。
1.2.3 内生真菌ITS序列的PCR扩增与测序 采用真菌通用引物ITS1与ITS4扩增真菌的ITS序列,聚合酶链式反应(PCR)体系为30 μL(1 μL DNA模板、1 μL ITS1、1 μL ITS4、15 μL 2×TaqMaster Mix、12 μL ddH2O),PCR反应参数:94 ℃预变性 5 min,94 ℃变性 1 min,55 ℃退火1 min,72 ℃延伸1 min,30~35次循环;72 ℃ 10 min,4 ℃停止。取5 μL PCR 产物进行琼脂糖凝胶电泳跑胶检测,得到明亮、单一的条带,送Songon Biotech (中国上海)公司测序。
1.2.4 内生真菌ITS序列BLAST比对和分析 将样品测序结果的序列在CongtingExpress软件拼接,在NCBI官网(https://blast.ncbi.nlm.nih.gov/Blast.cgi) 进行同源序列比对,下载相似
度最高序列建立系统发育树,用 MEGA 5.0 应用邻接法(Neighbor-joining,NJ)构建系统进化树,两个或两个以上菌株序列与同一个参考菌株序列在系统发育树中聚为一支且与参考序列相似度大于等于97%,则该菌株被鉴定到种水平,其余菌株为属水平[34]。
1.2.5 数据分析 分离率(%)=(分离得到的菌株数/组织块总数)×100%。分离率(Isolation Rate,IR)可以反映出样本中可培养内生真菌的丰富度。RStudio计算各地区的Chao1指数、ACE指数、香农(Shannon)指数、辛普森(Simpson)指数等α多样性指数,GraphPad Prism 9进行地区的多样性指数的差异性分析与绘图。
2 结果与分析
2.1 柴胡根部可培养内生真菌的分子鉴定
将ITS测序结果进行拼接后,在 NCBI 官网(https://blast.ncbi.nlm.nih.gov/Blast.cgi) 上进行同源序列比对,下载相似度最高的菌株序列,同等条件下选择已有相关文献报道的菌株序列,构建系统发育进化树分析亲缘和进化关系。部分菌株鉴定结果见表2、图2,如菌株1P 14-3与Fusariumoxysporum的同源相似性为99.82%,且在系统发育树中聚为一支,则1P 14-3被鉴定为Fusariumoxysporum。菌株1P 12-3与Phomasp.的同源相似性为99.27%,且在系统发育树中聚为一支,则1P 12-3被鉴定为Phomasp.。
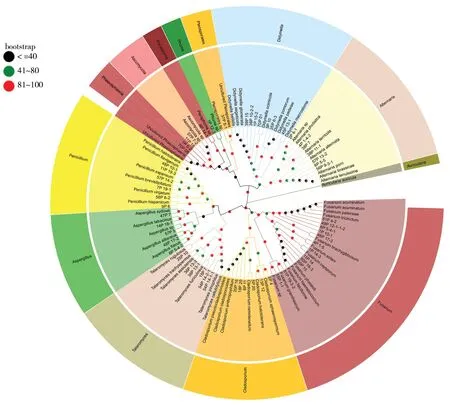
图2 柴胡根部可培养内生真菌中部分菌株与同源菌株的系统发育进化树Fig.2 Phylogenetic evolution tree of some strains and homologous strains of endophytic fungi that can be cultivated in the root of Bupleurum chinenseAuricularia auricula为外源菌株Auricularia auricula is exogenous strain
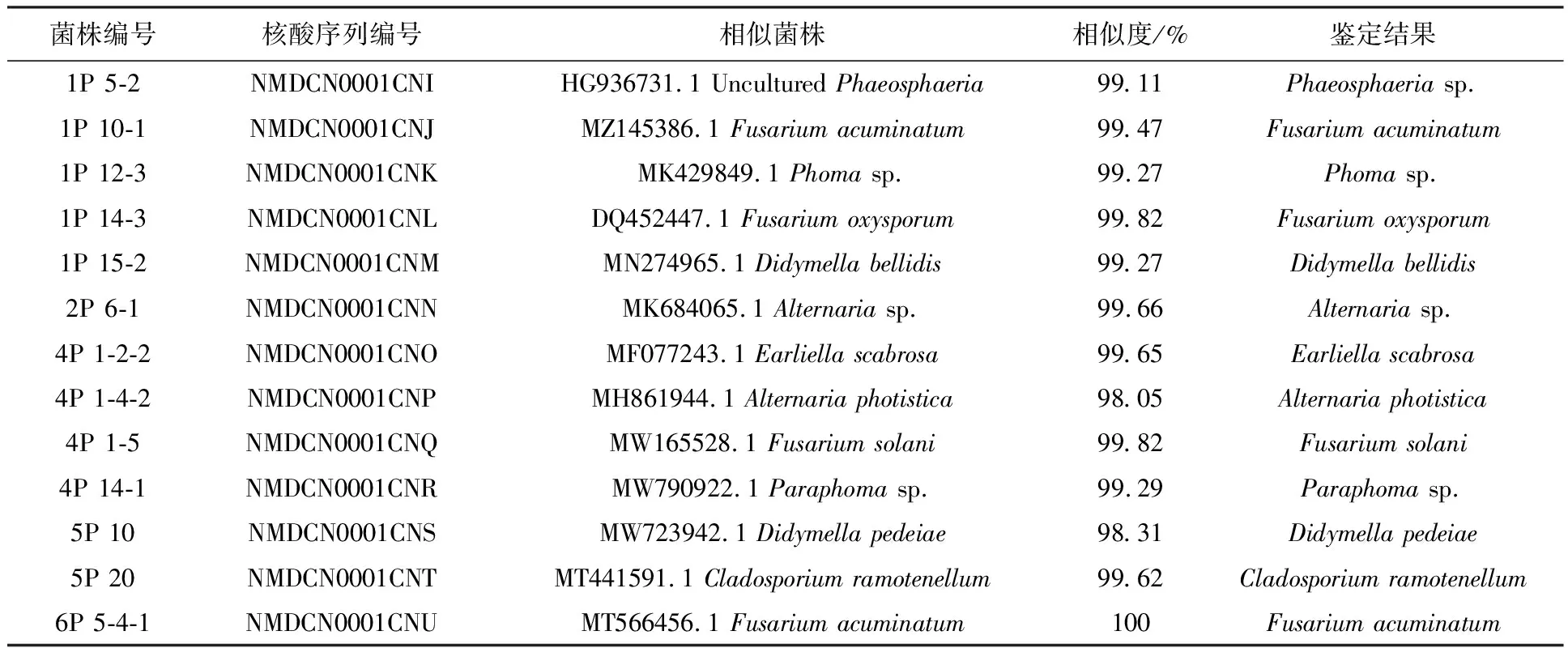
表2 柴胡根部可培养内生真菌中部分菌株BLAST比对结果
2.2 柴胡根部可培养内生真菌菌群的组成分析
全部样品中共分离得到705株内生真菌,根据ITS分子测序结果分析,其中695株真菌归属为4门14纲23目32科55属119种(表3),其余10株为Uncultured fungus,无具体鉴定结果。在对柴胡根部可培养内生真菌菌群进行不同分类水平分析时,相对丰度小于1%的菌群归为others,相对丰度大于5%的菌群归为优势菌群。在门水平,主要由半知菌门、子囊菌门、担子菌门、接合菌门等菌株组成,其中半知菌门(51.06%)、子囊菌门(29.50%)的相对丰度超过25%,为优势菌门。在纲水平主要由丝孢纲、腔孢纲、不整囊菌纲、担子菌纲、座囊菌纲、层菌纲、粪壳菌纲等菌株组成,其中丝孢纲(35.89%)、腔孢纲(14.18%)、不整囊菌纲(13.76%)、担子菌纲(12.20%)、座囊菌纲(11.21%)相对丰度均超过10%,为优势菌纲。在目水平主要由瘤唑孢目、球壳孢目、散囊菌目、假球壳目、格孢腔目、丝孢目、多孔菌目、假球壳目、伞菌目等菌株组成,其中瘤唑孢目(31.49%)、球壳孢目(14.04%)、散囊菌目(13.76%)、假球壳目(12.20%)相对丰度均超过10%,为优势菌目。在科水平主要由瘤唑孢科、球壳孢科、隔孢假壳科、散囊菌科、孢腔菌科、暗丛梗孢科、多孔菌科、葡萄座科、丝裂孢科、发菌科等菌株组成,其中瘤唑孢科(31.49%)、球壳孢科(13.90%)、隔孢假壳科(12.20%)、散囊菌科(11.35%)相对丰度均超过10%,为优势菌科。在属水平上主要由Fusarium、Didymella、Aspergillus、Paraphoma、Alternaria、Phoma、Cladosporium、Ascomycota、Phaeosphaeria、Pleosporales、Penicillium、Talaromyces、Earliella、Botryosphaeria等菌株组成,通过统计各菌属的相对丰度发现Fusarium(31.94%)、Didymella(12.37%)、Aspergillus(11.51%)、Paraphoma(7.19%)、Alternaria(6.91%)的相对丰度均超过5%,故柴胡根部可培养内生真菌的优势菌属为Fusarium、Didymella、Aspergillus、Paraphoma、Alternaria。在种水平主要由Fusariumacuminatum、Didymellabellidis、Fusariumtricinctum、Aspergillusfumigatus、Fusariumsolani、Paraphomachrysanthemicola、Alternariaalternata、Fusariumoxysporum、Cladosporiumcladosporioides、Didymellaglomerata等菌株组成,Fusariumacuminatum(9.50%)、Didymellabellidis(8.65%)、Fusariumtricinctum(8.51%)、Aspergillusfumigatus(8.51%)、Fusariumsolani(5.96%)、Paraphomachrysanthemicola(5.11%)的相对丰度均超过5%,为柴胡根部可培养内生真菌的优势菌种。

表3 柴胡根部可培养内生真菌的分类结果
2.3 不同地区的柴胡根部可培养内生真菌的分离率分析
本研究从70批样品2 800个组织块中,共分离出705株内生真菌,柴胡根部可培养内生真菌的总体分离率为25.18%。从图1可知,样品采集地主要分布在山西省的南部区与北部区,分别简称为晋南与晋北。晋南区域的分离率(29.07%)高于晋北区域(20.49%),晋南区域内生真菌丰富度要高于晋北区域,对晋南与晋北两个地区总的分离率进行差异分析(图3)结果表明,晋南区域与晋北区域的可培养内生真菌的分离率没有显著差异,表明两个区域的柴胡根部可培养内生真菌丰富度没有显著差异,从这两个区域的柴胡根部可分离出的真菌总量没有显著差异。
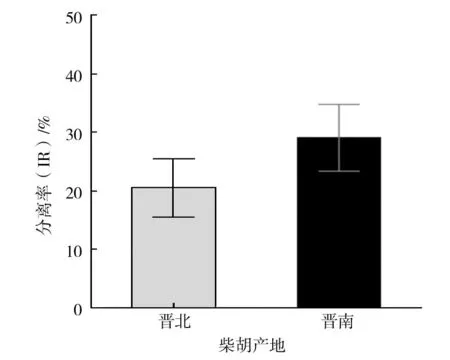
图3 晋南与晋北区域分离率差异分析Fig.3 Difference analysis of regional isolation rate difference between southern and northern Shanxi
晋南与晋北各地区分离率如图4所示,晋南地区中LC(65.00%)、HT(64.17%)、WX(53.75%)、HM(40.00%)、YC(31.88%)的分离率高于晋南地区的平均分离率(29.07%)。晋北地区中GL(65.00%)、LQ(40.00%)、SY(23.33%)、HY(22.50%)、PL-1(22.50%)的分离率高于晋北地区的平均分离率(20.49%)。将晋南与晋北各地区分离率分别进行anova分析(表4),HT(64.17%)地区的分离率显著(P<0.05)高于HY(22.50%)、ZY(17.08%)、NW(9.17%)、PG(5.00%)、BD(5.00%)五个地区;表明HT地区的内生真菌丰富度高于HY、ZY、NW、PG和BD地区。GL(65%)地区的分离率极显著(P<0.01)高于XF(17.00%)、JS(9.17%)地区,显著(P<0.05)高于 J(22.00%)、X(15.00%)和PL-2(3.75%)地区,表明GL地区的内生真菌丰富度高于XF、JS、J、X和PL-2地区。以上数据显示不同地区的分离率各有不同,且差异较大,表明各地的柴胡根部可培养内生真菌丰富度差异较大。
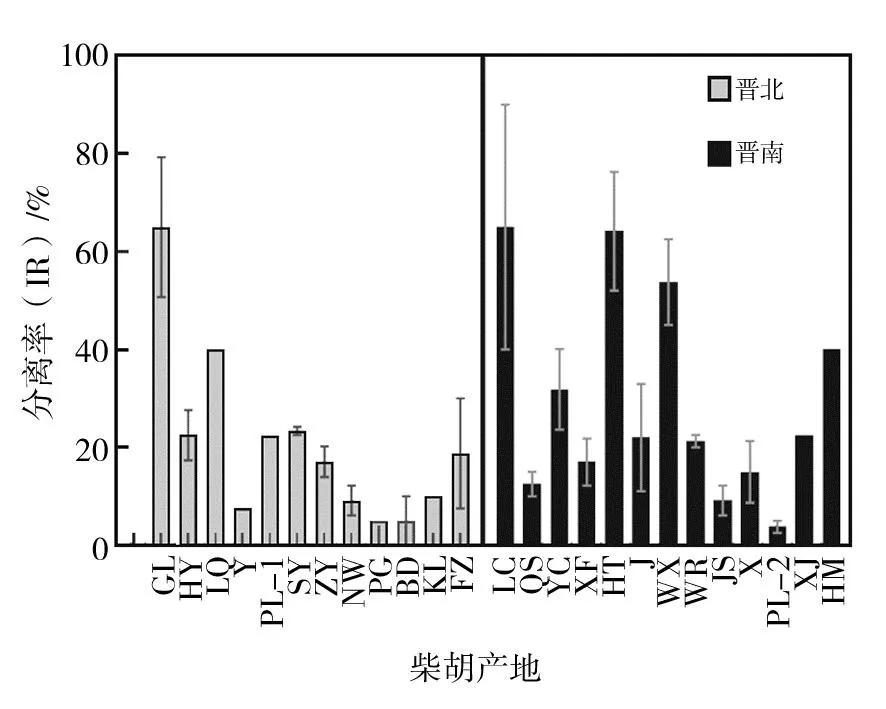
图4 各地区分离率Fig.4 Isolation rate in different regions

表4 晋南与晋北各地区分离率anova分析
2.4 不同地区的柴胡根部可培养内生真菌的优势菌群分析
分析不同地区的柴胡根部可培养内生真菌的物种组成(图5),晋南地区的优势菌属为镰刀菌属(Fusarium, 32.18%)、亚隔孢壳属(Didymella, 15.96%)、曲霉属(Aspergillus, 8.24%)、异茎点霉属(Paraphoma, 6.91%)、茎点球属(Phoma, 5.32%)、链格孢属(Alternaria, 5.05%),在晋南地区柴胡根部可培养内生真菌中的相对丰度均高于5%。晋北地区的优势菌属为Fusarium(31.66%)、Aspergillus(15.36%)、Alternaria(9.09%)、Didymella(8.15%)、Paraphoma(7.52%),在晋北地区柴胡根部可培养内生真菌中的相对丰度均高于5%。


图5 各地区不同分类水平下物种相对丰度Fig.5 Relative abundance of species in different regionsa:属水平物种相对丰度;b:种水平物种相对丰度;GL~FZ:晋北;LC~HM:晋南a: genus-level relative abundance; b: species-level relative abundance; GL-FZ: The northern part of Shanxi Province; LC-HM: The southern part of Shanxi Province
晋南地区的优势物种为Didymellabellidis(11.44%)、三线镰刀菌(Fusariumtricinctum, 7.98%)、腐皮镰刀菌(Fusariumsolani, 7.45%)、锐顶镰孢菌(Fusariumacuminatum, 5.85%)、烟曲霉(Aspergillusfumigatus, 5.85%),在晋南地区柴胡根部可培养内生真菌的相对丰度均高于5%。晋北地区的优势物种为Fusariumacuminatum(14.11%)、Aspergillusfumigatus(11.91%)、Fusariumtricinctum(9.40%)、菊异茎点霉(Paraphomachrysanthemicola, 5.64%)、Didymellabellidis(5.64%)、链格孢菌(Alternariaalternata, 5.64%),在晋北地区柴胡根部可培养内生真菌中相对丰度均高于5%。
2.5 不同地区柴胡根部可培养内生真菌的α多样性分析
α多样性也被称为生境内多样性(within-habitat diversity),是指一个特定地区或生态系统内的多样性。α多样性指数包括Chao1指数、ACE指数、香农指数、辛普森指数等。Chao1指数和ACE指数越大,表明某群落物种数目越多;香农指数越大,表明生物多样性越好;辛普森指数越大,说明群落中种数越多,各种个体分配越均匀,物种多样性程度越好。
对晋南地区与晋北地区进行共有属种分析(图6)发现,晋南地区和晋北地区共有菌属25个,包括Fusarium、Aspergillus、Alternaria、Didymella、Paraphoma、Phoma、Cladosporium、Phaeosphaeria、Pleosporales、Ascomycota、Earliella、Penicillium等;晋南地区特有菌属23个,包括Sarocladium、Macrophomina、Irpex等;晋北地区特有菌属8个,包括Peroneutypa、Ascochyta、Ophiobolus等。晋南地区和晋北地区共有菌种33个,包括Fusariumacuminatum、Aspergillusfumigatus、Fusariumtricinctum、Paraphomachrysanthemicola、Didymellabellidis、Alternariaalternata、Fusariumsolani、Cladosporiumcladosporioides、Didymellaglomerata等;晋南地区特有菌种57个,包括Phomabellidis、Fusariumproliferatum、Fusariumfalciforme、Macrophominaphaseolina、Torulaligniperda等;晋北地区特有菌种32个,包括Alternariaphotistica、Cladosporiumramotenellum、Penicilliumflavigenum、Alternariacarotiincultae、Aspergillusaculeatus等。

图6 不同地区柴胡根部可培养内生真菌的Venn图Fig.6 Venn diagram of cultivable endophytic fungal species from roots of Bupleurum chinense in different regions a:属水平;b:种水平a: genus level; b: species level
对晋南地区与晋北地区进行α多样性指数差异性分析(图7)发现,在属水平上,晋南地区的各α多样性指数均高于晋北地区,且晋南的Simpson指数显著高于晋北地区。在种水平上,晋南地区的各α多样性指数均高于晋北地区,且晋南的Shannon指数与Simpson指数均显著高于晋北地区。表明晋南地区柴胡根部可培养内生真菌的生物多样性要高于晋北地区,物种分配的均匀度也好于晋北地区。
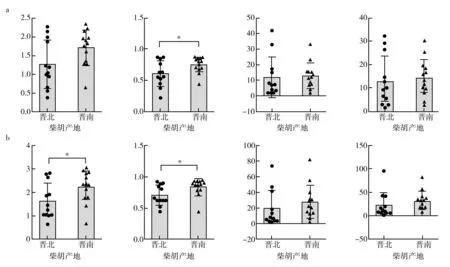
图7 晋南区域与晋北区域柴胡根部可培养内生真菌α多样性指数差异分析Fig.7 Endophytic fungi can be cultivated in the root of Bupleurum chinense in southern and northern Shanxi α Diversity index difference analysisa:属水平;b:种水平;*P<0.05a: genus level; b: the species level;*P<0.05
对不同地区的柴胡根部可培养内生真菌进行α多样性分析。①属水平:J(2.340)、ZY(2.269)、XF(2.222)3个地区的Shannon指数最高,且J>ZY>XF;Y(0.637)、PL-2(0.637)、KL(0.562)、LQ(0.377) 四个地区的Shannon指数最低,且Y=PL-2>KL>LQ。J(0.871)、XF(0.869)、ZY(0.868)三个地区的Simpson指数最高,且J>XF>ZY;Y(0.444)、PL-2(0.444)、KL(0.375)、LQ(0.219)四个地区的Simpson指数最低,且Y=PL-2>KL>LQ。表明J、XF、ZY 三个地区的生物多样性最好,且物种分配的均匀度高;Y、PL-2、KL、LQ 四个地区的生物多样性最差,且物种分配的均匀度较低。ZY(42.000)、WX(33.000)、HY(33.000)三个地区的Chao1指数最高,且ZY>WX=HY;Y(2.000)、PL-2(2.000)、KL(2.000)、LQ(2.000)四个地区的Chao1指数最低,且Y=PL-2=KL=LQ。HY(32.517)、X(30.529)、SY(29.667)三个地区的Ace指数最高,且HY>X>SY;Y(3.000)、PL-2(3.000)、LQ(2.000)三个地区的Ace指数最低,且Y=PL-2>LQ。表明ZY、WX、HY、X、SY 五个地区的群落物种数目最多;Y、PL-2、KL、LQ 四个地区的群落物种数目最少(表5)。②种水平:J(3.048)、HT(2.912)、XF(2.838)三个地区的Shannon指数最高,且J>HT>XF;PL-1(1.003)、Y(0.637)、PL-2(0.637)三个地区的Shannon指数最低,且PL-1>Y=PL-2。J(0.946)、HT(0.928)、XF(0.926)三个地区的Simpson指数最高,且J>HT>XF;PL-1(0.519)、Y(0.444)、PL-2(0.444)三个地区的Simpson指数最低,且PL-1>Y=PL-2。表明J、HT、XF 三个地区的生物多样性最好,且物种分配的均匀度高;PL-1、PL-2、Y 三个地区的生物多样性最差,且物种分配的均匀度较低。WX(81.000)、HY(74.167)、XF(56.000)三个地区的Chao1指数最高,且 WX>HY>XF;PG(3.500)、BD(3.500)、KL(3.500)、Y(2.000)、PL-2(2.000)五个地区的Chao1指数最低,且PG=BD=KL>Y=PL-2。HY(95.804)、WX(81.258)、XF(57.486)三个地区的Ace指数最高,且HY>WX>XF;PG(6.000)、BD(6.000)、KL(6.000)、Y(3.000)、PL-2(3.000)五个地区的Ace指数最低,且PG=BD=KL>Y=PL-2。WX、HY、XF 三个地区的群落物种数目最多;PG、BD、KL、 PL-2、Y 五个地区的群落物种数目最少(表6)。

表5 各地区柴胡根部可培养内生真菌α多样性指数(属水平)
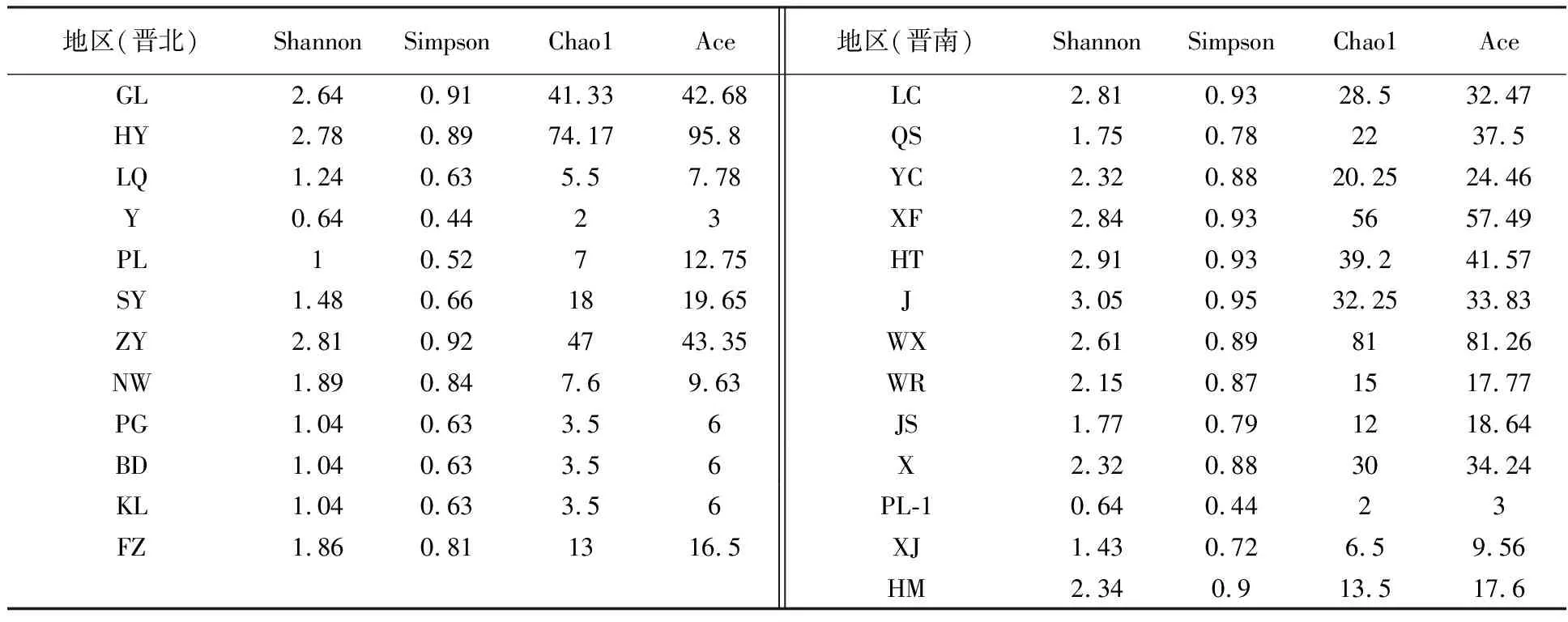
表6 各地区柴胡根部可培养内生真菌α多样性指数(种水平)
3 讨 论
本研究发现柴胡根部可培养内生真菌的优势菌属为Fusarium、Didymella、Aspergillus、Paraphoma、Alternaria。但不同地区的优势菌属有所不同,晋南地区的优势菌属为Fusarium、Didymella、Aspergillus、Paraphoma、Phoma、Alternaria;晋北地区的优势菌属为Fusarium、Aspergillus、Alternaria、Didymella、Paraphoma。结合已有报道及本研究分析发现,Fusarium、Alternaria中部分菌种常为柴胡中的病原菌[35-37],而实验过程中所用实验样品植株并未表现出病态,说明此时柴胡根部微生物菌群还处于动态平衡状态,随着柴胡生长年限的延长,生活力的衰退,植物体内菌群失衡,病原菌可能逐渐占据优势,使植株出现病态[38-39],这可能是柴胡随着栽培年限变长,病害逐步加重的主要原因。
本研究通过对不同地区的柴胡根部可培养内生真菌的分离率和α多样性进行差异分析,发现晋南地区与晋北地区的柴胡根部可培养内生真菌丰富度与物种多样性存在一定的差异。晋南地区柴胡根部可培养内生真菌丰富度要高于晋北地区,晋南地区柴胡根部可培养内生真菌多样性要显著高于晋北地区,物种分配的均匀度也好于晋北地区。这与相关学者对杜仲及锁阳等药用植物内生菌多样性的研究结果类似。同时有研究发现不同地理-气候环境和土壤类型是造成植物内生真菌物种丰度和群落组成差异的主要因素[40-41],而晋南与晋北在地理-气候环境和土壤类型等方面的差异可能也是造成晋南与晋北地区柴胡根部可培养内生真菌的生物多样性存在差异的主要原因。
本课题组前期通过测定山西省北部与南部产地的柴胡中柴胡皂苷a、柴胡皂苷c、柴胡皂苷d、柴胡皂苷e、柴胡皂苷f、总柴胡皂苷的含量和挥发性物质,发现山西省北部产地的柴胡中柴胡皂苷含量高于山西省南部产地的柴胡[42],而山西省南部产区的柴胡中挥发性物质种类多于山西省北部产区的柴胡[31]。研究发现黄芩中黄酮类物质的含量与黄芩中链格孢菌的相对丰度有显著性关系[4],提示不同产地柴胡中内生真菌的种类与数量可能也是影响柴胡中活性成分产生差异的原因。还有研究发现植物挥发性物质有抑菌效果[43]。而本研究发现,晋南地区柴胡根部可培养内生真菌的优势物种中有Fusariumsolani(7.45%),因此推断Fusariumsolani感染柴胡后,有可能刺激柴胡产生挥发性物质进行抗菌实现自我防御。故Fusariumsolani的存在可能是导致柴胡质量下降,挥发性物质增加的主要原因之一。
本研究分析评价了柴胡根部可培养内生真菌的群落结构和物种多样性,建立了柴胡根部内生真菌库,为后期研究内生真菌与柴胡的互作,筛选促生菌株、生防菌和可以产生特定次级代谢产物的菌株提供了资源。
猜你喜欢
遵义医科大学学报(2023年1期)2023-02-06
山西林业(2021年2期)2021-07-21
今日农业(2020年22期)2020-12-25
邯郸学院学报(2020年2期)2020-08-11
中成药(2018年4期)2018-04-26
中成药(2017年8期)2017-11-22
国土与自然资源研究(2016年5期)2017-01-04
广州文博(2016年0期)2016-02-27
大众考古(2015年8期)2015-06-26
农产品市场周刊(2014年22期)2014-08-22
