
Bacillomycin D合成酶系NRPS硫酯酶结构域位移对脂肽合成的影响
2023-02-02 11:41张平陈美容马文杰陆兆新吕紫岩吕凤霞赵海珍别小妹
南京农业大学学报 2023年1期
张平,陈美容,马文杰,陆兆新,吕紫岩,吕凤霞,赵海珍,别小妹*
(1.南京农业大学食品科学与技术学院,江苏 南京 210095;2.中国药科大学药学院,江苏 南京 211198)
抗菌脂肽Bacillomycin D属于伊枯草菌素(Iturin)家族,是一类由枯草芽孢杆菌、解淀粉芽胞杆菌等芽胞杆菌属微生物合成的脂肽类物质,由脂质和脂肽2部分构成,脂质部分为β-氨基脂肪酸链(C11—C17),脂肽氨基酸序列为Asn-Tyr-Asn-Pro-Glu-Ser-Thr,2部分通过苏氨酰-β-氨基键连接。Bacillomycin D对黄曲霉菌、赭曲霉菌、匍枝根酶菌等病原真菌具有很强的抑制效果[1-4],有潜力成为高效安全的食品防腐剂,应用于食品和粮食的防腐。
Bacillomycin D属于非核糖体肽,其合成由bmy基因簇控制,包含bmyD、bmyA、bmyB和bmyC4个基因[5]。在bmyC编码的Bmy C亚基C末端含有1个硫酯酶TE结构域,属于α/β水解酶家族,能断裂多肽载体蛋白(PCP)和酰基分子之间的共价硫脂键[6],负责环脂肽中间体的环化和水解。TE结构域具有典型的Ser-His-Asp催化三联体,Asp对His残基有稳定作用,通过His接受Ser羟基的质子H,Ser作为亲核试剂。活性位点区域上方有1个lid 区域,作用是控制底物与活性位点的结合,以防止不需要的底物结合[6],此外lid结构域还可以根据底物寡聚程度而采用多种构象,并最终控制环化[7]。
TE结构域具有区域特异性、化学特异性和立体特异性[6]。目前针对大环内酯TE结构域的研究大多集中于在TE结构域的底物特异性和晶体结构表征:体外表达gramicidin S合成酶中的迭代双模块体系PCP-TE,具有二聚化和三聚化活性,但随着底物肽链长度的不断增加,PCP-TE的二聚化和三聚化反应能力降低[8]。Liu等[9]分析了PCP-TE的晶体结构,指出疏水作用力介导了两结构域结合的相互作用,而酰基-TE中间体的结构表征[10]有利于确定假定的氧阴离子空穴存在,并证明了TE结构域中lid 区域与底物的相互作用。非核糖体肽合成酶(nonribosomal peptide synthetases,NRPS)具有模块化生物合成规则,有利于对其进行改造,增强其生物催化特性或天然产物多样性。Niquille等[11]对酪氨酸合成途径中识别L-Phe的A结构域进行随机突变与高通量筛选,筛选出的A结构域能以近天然的特异性和效率接受和处理底物(S)-β-Phe。Steiniger等[12]依次使用cyclosporine合成途径中的模块2、4、7和10取代了ennitain和bassianolide生物合成途径中的第2个模块,生成了新型环脂肽类似物。但是关于Bacillomycin D合成酶系及其TE结构域的研究国内外鲜见报道。
本试验对硫脂酶TE结构域进行前移,在体内改变其在Bacillomycin D合成酶系中的位置,构建新的合成酶系,形成一系列截短的Bacillomycin D脂肽类似物,分析其结构和抑菌活性等,明确硫酯酶结构域位移对Bacillomycin D类似物合成的影响,旨在探索合成酶结构修饰与新型抗菌脂肽的合成及构效规律。
1 材料与方法
1.1 试验材料
1.1.1 试验菌株和质粒本试验所用菌株解淀粉芽胞杆菌(Bacillusamyloliquefaciens)fmbJ、克隆宿主大肠杆菌(Escherichiacoli)JM109、黄曲霉菌(Aspergillusflavus)均由南京农业大学酶工程实验室保藏。克隆宿主大肠杆菌(Escherichiacoli)JM110 购买自北京全式金生物公司;克隆载体 pMD19-T购买自TaKaRa生物公司,温敏型质粒pKS2由本课题组保存。
1.1.2 工具酶和试剂细菌基因组提取试剂盒购自美国OMEGA公司;DNA凝胶回收试剂盒、质粒提取试剂盒购自南京诺唯赞生物科技有限公司;T4DNA连接酶购自大连宝生物公司;氨苄青霉素、红霉素、卡那霉素及低熔点琼脂购自上海生工生物工程公司;酵母提取物、胰蛋白胨等购自英国Oxoid公司;其他化学试剂均为分析纯,购自国药集团公司。
1.1.3 培养基和溶液配制LB培养基:胰蛋白胨10.0 g,酵母提取物5.0 g,氯化钠10.0 g,1 000 mL去离子水。LBS培养基:在LB培养基中加入山梨醇91.0 g。种子培养基:牛肉浸膏3.0 g,蛋白胨10.0 g,NaCl 5.0 g,1 000 mL去离子水,pH(7.0~7.2)。发酵培养基:葡萄糖20.0 g,L-谷氨酸5.0 g,酵母浸膏1.0 g,KH2PO40.5 g,MgSO4·7H2O 0.5 g,KCl 0.5 g,CuSO4·5H2O 0.15 mg,FeSO4·7H2O 1.2 mg,MnSO45 mg,1 000 mL去离子水,pH7.0。PDA培养基:马铃薯200.0 g,葡萄糖20.0 g,琼脂20 g,1 000 mL去离子水。电转化液:0.5 mol·L-1海藻糖,0.5 mol·L-1山梨醇,0.5 mol·L-1甘露醇,10%(体积分数)甘油。
1.1.4 主要仪器PTC-100TM PCR仪购于MJ Research 公司;电转化仪MicroPulserTM购于Bio-Rad 生命医学产品有限公司;Ultimate3000 高效液相分析系统购于美国 Dionex 公司;NanoDrop 2000 微量紫外-可见分光光度计购于美国 Thermo 公司;HYG-A全温摇瓶柜购于太仓实验设备厂;Eppendorf 5418 离心机购于德国 Eppendorf 公司;G2-XS-QTof高分辨液相质谱购于Waters公司。
1.2 试验方法
1.2.1 重组整合质粒pKS2-M5-TELong和pKS2-M6-TELong的构建根据B.amyloliquefaciensfmbJ全基因组序列,利用Primer 5.0 软件设计PCR反应特异引物,序列见表1,由金斯瑞生物科技有限公司合成。

表1 用于质粒构建的PCR引物Table 1 Primers for plasmids construction
参照OMEGA细菌基因组提取试剂盒提取B.amyloliquefaciensfmbJ基因组DNA,以基因组DNA为模板,分别采用M5-TELong-U-F/R扩增上游同源臂M5-TELong-U;采用M5-TELong-D-F/R扩增下游同源臂 M5-TELong-D,上、下游同源臂有部分序列同源。以上述同源臂为模板,重叠延伸PCR扩增目的基因,进行TATA克隆,获得T-M5-TELong质粒;同理获得T-M6-TELong质粒,送苏州金唯智生物科技有限公司测序验证。
将测序正确的质粒和温敏型质粒pKS2进行SalⅠ和KpnⅠ双酶切处理,琼脂糖凝胶电泳后回收目的基因和线性pKS2片段,采用T4DNA连接酶进行连接,构建重组整合质粒。测序验证后,转入E.coliJM110,获得可用于电转化的去甲基化重组整合质粒,pKS2-M5-TELong和pKS2-M6-TELong。
1.2.2 解淀粉芽胞杆菌前移菌株fmbJ-M5-TELong和fmbJ-M6-TELong的构建制备B.amyloliquefaciensfmbJ电转感受态细胞和电转化的具体过程参照文献[13]。温敏型质粒pKS2介导的同源重组过程参照文献[14],将经PCR和测序验证正确的重组突变菌株-80 ℃保存,并分别命名为fmbJ-M5-TELong和fmbJ-M6-TELong。
1.2.3 抗菌脂肽Bacillomycin D类似物的HPLC和MS分析将原始菌株fmbJ以及突变菌株活化后的单菌落,接种于种子培养基,在37 ℃、180 r·min-1培养至对数生长期,按照5%(体积分数)的接种量转接发酵培养基,33 ℃、180 r·min-1发酵培养120 h后即得发酵粗提液。Bacillomycin D提取方法参照Qian等[2]的方法进行,高效液相色谱(HPLC)检测条件和液质联用(LC-MS)条件参照Gong等[1]的方法进行。
1.2.4 抗菌脂肽Bacillomycin D类似物抑菌试验将黄曲霉菌接种PDA培养基,于 30 ℃ 培养至产生孢子。加入5 mL无菌生理盐水,清洗孢子,用血球计数板调整孢子悬液浓度至 1×106mL-1。添加1 mL孢子悬液至50 ℃左右的PDA中,倒入平板中,晾干后打孔,加入50 μL Bacillomycin D类似物粗提液,以甲醇作为对照,30 ℃培养48~72 h,观察并记录抑菌圈大小。
1.2.5 TE结构域前移对PCP-TE双结构域的影响与预测采用AlphaFold蛋白结构数据库(https://colab.research.google.com/github/deepmind/alphafold/blob/main/notebooks/AlphaFold.ipynb)对结构域前移菌株和野生菌株的Bacillomycin D合成酶末端的PCP-TE双结构域的蛋白质构象进行预测,利用Ligplot+软件[15]分析PCP-TE结构域之间的相互作用。
1.3 数据分析

2 结果与分析
2.1 解淀粉芽胞杆菌突变菌株fmbJ-M5-TELong和fmbJ-M6-TELong的构建
通过温敏型质粒pKS2参与的两步法同源重组敲除Bacilomycin D pKS/NRPS合成酶系末端2个模块(8 540 bp)和最后1个模块(4 563 bp),将负责环化和水解的硫酯酶结构域TE分别前移至模块5末端和模块6末端,改造后的合成酶系见图1。
两步法同源重组双交换后,平板法筛选出在抗生素平板上不能生长但在无抗性平板能够生长的抗生素敏感菌株,分别以M5-TELong-U-F/R和M6-TELong-U-F/R为引物对进行PCR验证。图2-A中,泳道2、3和泳道5~7有略低于2 000 bp的条带,产物测序验证为前移菌株fmbJ-M5-TELong;泳道1、4和8无条带,为回复型突变菌株。图2-B中,泳道1、2和泳道4~6有略低于2 000 bp的条带,产物测序为前移菌株fmbJ-M6-TELong;泳道3和7没有条带,为回复型突变菌株。

图1 Bacillomycin D合成酶系中硫酯酶结构域TE前移示意图[5,16]Fig.1 A schemmatic represention of thioesterase domain TE forward in Bacillomycin D synthetases[5,16] A. 原始的Bacillomycin D合成酶Natural Bacillomycin D synthetases;B. TE结构域前移至模块6末端后的Bacillomycin D合成酶Bacillomycin D synthetases after TE moved forward to the end of module 6;C. TE结构域前移至模块5末端后的Bacillomycin D合成酶Bacillomycin D synthetases after TE moved forward to the end of module 5.

图2 抗生素敏感菌株菌落的PCR凝胶电泳检测Fig.2 Gel electrophoresis detection of colony PCR of antibiotic sensitive strain M.2000 DNA marker;A. Lane 1-8:fmbJ-M5-TELong抗生素敏感菌株Antibiotic sensitive strains of fmbJ-M5-TELong;B. Lane 1-7:fmbJ-M6-TELong抗生素敏感菌株Antibiotic sensitive strains of fmbJ-M6-TELong.
2.2 抗菌脂肽Bacillomycin D类似物的HPLC-MS分析
突变菌株fmbJ-M5-TELong和fmbJ-M6-TELong发酵粗提液HPLC检测结果显示,突变菌株产物的保留时间与fmbJ一致,初步推测为截短的Bacilomycin D类似物(图3)。高分辨率LC-ESI-MS/MS检测结果显示,在fmbJ-M5-TELong发酵粗提液中,检出了预测的Bacilomycin D衍生脂肽(线性五肽和环状五肽)的分子量,通过MS/MS分析进一步确定了上述Bacilomycin D衍生脂肽的结构(表2)。以线性五肽离子分子量(m/z)861.5为母离子进行MS/MS 碎片离子分析(图4),肽键断裂后形成b-和y-这2种电荷离子,从实际检测到的b-和y-离子分子量确定了母离子分子量为861.5的脂肽的肽端部分为Asn-Tyr-Asn-Pro-Glu,脂肪酸链部分为含有14个碳的β-NH2脂肪酸,分子式为C41H64O12N8。此外还检测出母离子峰分子量875.5,与之前的线性五肽分子量间相差14,是由C15的脂肪酸链组成的线性五肽同系物。母离子m/z843.5和857.5与线性五肽的分子量相差18,选取母离子m/z843.5进行MS/MS分析,由生成的二级碎片特征离子峰227.102 6、617.365 4和212.103 2,确定其结构为环状五肽C14β-NH2FA(Asn-Tyr-Asn-Pro-Glu),分子式为C41H62O11N8。

图3 突变菌株产物的HPLC检测Fig.3 HPLC detection of products of mutant strains1~4.发酵产物的色谱峰Chographic peaks of fermentation products.

表2 从突变菌株中检测到的 Bacillomycin D衍生新型脂肽Table 2 Novel Bacillomycin D derivatives detected in crude extracts isolated from mutants
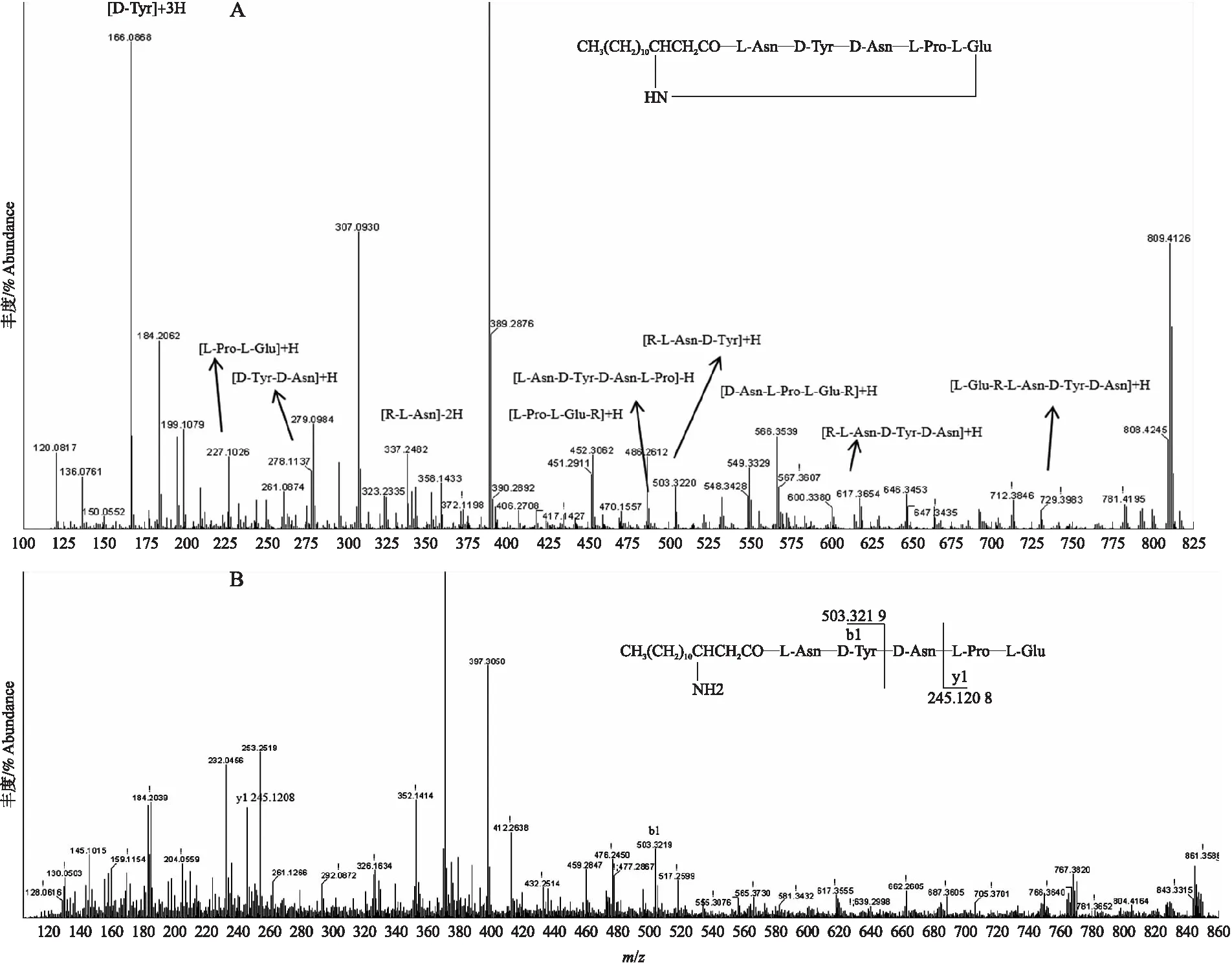
图4 环状脂五肽离子m/z为843.5(A)和线性脂五肽离子m/z为861.5(B)的MS/MS碎片离子分析Fig.4 MS/MS fragment ion analysis of the cyclic pentapeptide at m/z 843.5(A)and linear pentapeptide at m/z 861.5(B)
突变菌株fmbJ-M6-TELong发酵粗提液中,检测出m/z948.5、962.5、976.5、990.6,m/z分别相差14,依次为C14、C15、C16和C17的同系物,以m/z948.5为母离子进行MS/MS分析(图5),b-特征离子峰340.257 8→503.321 5→617.362 7→697.393 4→843.457 5,y-特征离子峰332.145 0→446.188 4→609.250 3,由此确定肽端为Asn-Tyr-Asn-Pro-Glu-Ser,脂肪酸链部分为含有14个碳的β-NH2脂肪酸,分子式为 C44H69O14N9,此外还检测出了m/z902.5、916.5、930.5、958.5和972.5,与线性六肽相差18,以m/z930.5为母离子,由生成的二级碎片特征离子峰341.143 8、590.354 9,确定其结构为环状脂六肽C14β-NH2FA(Asn-Tyr-Asn-Pro-Glu-Ser),分子式为C44H67O13N9。

图5 环状六肽离子m/z为930.5(A)和线性六肽离子m/z为 948.5(B)的 MS/MS 碎片离子分析Fig.5 MS/MS fragment ion analysis of the cyclic hexapeptide at m/z 930.5(A) and linear hexapeptide at m/z 948.5(B)
2.3 抗菌脂肽Bacillomycin D类似物抑菌效果
如图6所示:fmbJ-M5-TELong发酵合成的线性五肽和环状五肽Bacillomycin D类似物的抑菌圈直径(22.89±0.20)mm显著大于fmbJ的抑菌圈直径(21.05±0.18)mm,但随着黄曲霉的生长,抑菌效果逐渐变差,圈内出现部分菌丝,不能完全抑制黄曲霉菌的生长;fmbJ-M6-TELong发酵合成的线性六肽和环状六肽抗菌脂肽类似物的抑菌圈直径(23.91±0.28)mm大于fmbJ和 fmbJ-M5-TELong合成的脂肽类似物,但抑菌效果也随着黄曲霉菌的生长逐渐降低。综上所述,尽管新型脂肽前期的抑菌圈直径显著大于天然Bacillomycin D,但是后期并不能完全抑制。
2.4 TE结构域前移对PCP-TE双结构域的影响
TE结构域前移至模块5和模块6末端后,Bacillomycin D合成酶末端的PCP-TE双结构域相对构象发生变化。如图7所示:PCP蛋白由4个α-螺旋和1个短螺旋(4个氨基酸)构成,TE蛋白则由7个α-螺旋和7个β-折叠构成,此外,天然的PCP-TE双结构域蛋白中TE结构域的第1个β-折叠能被正确预测,而TE前移后的双结构域中TE蛋白在第1个本应是β-折叠处显示为loop区域。使用Ramachandran plot法[17]对3个PCP-TE双结构域三维结构进行评估,图8显示了3个结构模型中α-C原子和N原子间的二面角φ以及α-C原子和羰基C原子间键的二面角ψ的分布情况,M7-PCP-TE、M6-PCP-TE和M5-PCP-TE中分别有99.7%、100%和99.3%的φ和ψ二面角分布在合理区域内,均属于正常分布,且可信度较高。

图6 fmbJ(A)、fmbJ-M5-TELong(B)和fmbJ-M6-TELong(C)发酵产物抑菌图谱Fig.6 Antibacterial spectrum of fermentation products of fmbJ(A),fmbJ-M5-TELong(B) and fmbJ-M6-TELong(C)
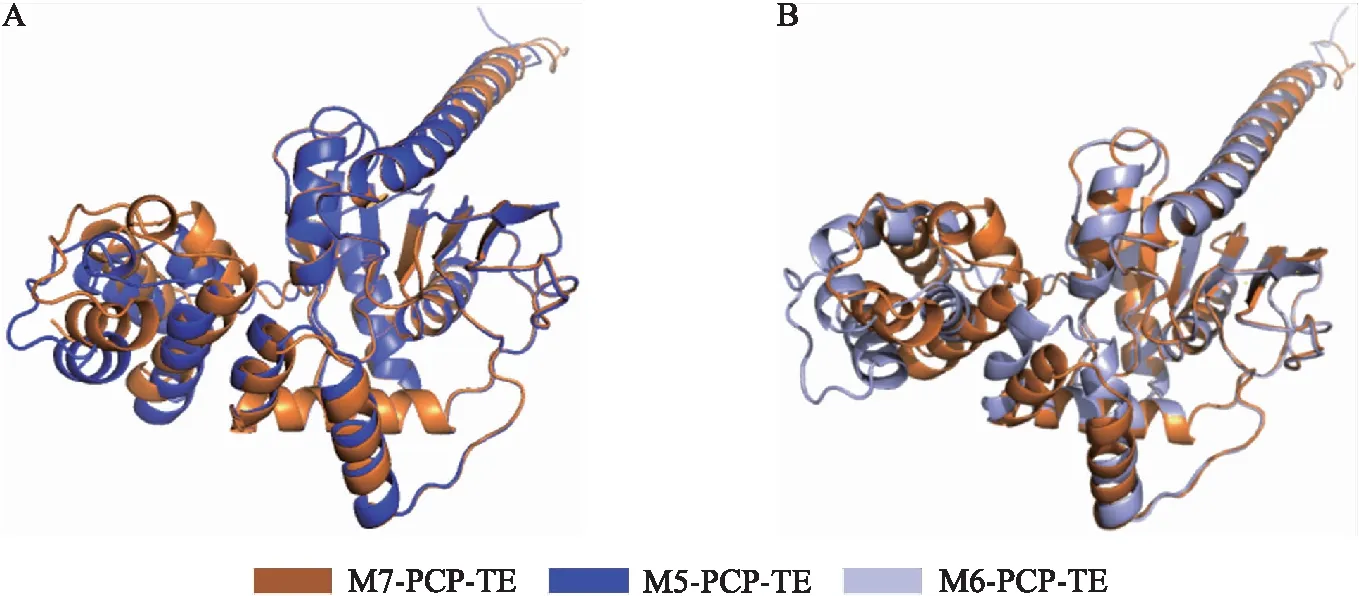
图7 Bcillomycin D合成酶末端PCP-TE双结构域三维结构比对示意图Fig.7 Schematic diagrams of three-dimensional structure comparison of PCP-TE didomains at the end of Bcillomycin D synthases A.M7-PCP-TE和M5-PCP-TE示意图Diagrams between M7-PCP-TE and M5-PCP-TE;B. M7-PCP-TE和M6-PCP-TE示意图Diagrams between M7-PCP-TE and M6-PCP-TE.

图8 PCP-TE三级结构的拉氏构象图Fig.8 Ramachandran plots of three-dimensional structure of PCP-TE A. M7-PCP-TE的拉氏构象图Ramachandran plots of M7-PCP-TE;B. M5-PCP-TE的拉氏构象图Ramachandran plots of M5-PCP-TE;C. M6-PCP-TE的拉氏构象图Ramachandran plots of M6-PCP-TE.
使用Ligplot+软件分析天然的模块7末端的PCP-TE双结构域之间的相互作用力,如图9-A所示,包括α2PCP/α3PCP和β1TE/α1TE之间的疏水作用力,α2PCP和TE结构域lid regions(α5TE/α6TE)之间的疏水作用力,PCP结构域上His48和TE结构域上Asp119形成的一个盐桥相互作用;当TE结构域前移至模块5末端时,PCP-TE双结构域之间的相互作用见图9-B,包括α2PCP和TE结构域lid regions(α5TE/α6TE)之间的疏水作用力,α3PCP和β1TE/α1TE之间的疏水作用力。PCP结构域上Thr36和TE结构域上Ser217形成的一个氢键相互作用;当TE结构域前移至模块6末端时,只有α2PCP和TE结构域lid regions(α5TE/α6TE)之间的疏水作用力与PCP结构域上Ser30和TE结构域上Tyr209形成的一个氢键相互作用(图9-C)。综上所述,硫酯酶TE易位前后,PCP-TE双结构域蛋白的相对构象发生变化,双结构域之间的相互作用力也随之变化,但α2PCP和TE结构域lid regions(α5TE/α6TE)之间的疏水作用力无明显差异,表明此疏水作用是维系PCP-TE双结构域三维结构和功能的主要作用力。
3 讨论
非核糖体肽合成方式广泛存在于细菌和真菌中,合成一些具有生物活性的次级代谢产物,如达托霉素、表面活性剂、环孢菌素A等,相当多的天然产物为大环化合物,化学合成此类物质难度较大,而NRPS具有酶复合物-亚单位-模块-结构域的分级结构[18],其模块化生物合成逻辑和共线性规则激发了研究人员对其结构域和模块重新编程的热情[19-20],设计新的NRPS装配线,以建立无法化学合成的新化合物库。位于NRPS装配线末端的TE结构域负责水解和环化中间产物,形成线性或大环产物,有研究指出TE结构域的N-端延伸结构,即天然的PCP-TE-linker,是联系PCP结构域和TE结构域的有效通讯工具[21],对维持TE蛋白稳定性和正确执行相应水解或环化功能有不可或缺的作用。
本研究采用组合生物学方法对Bacillomycin D合成酶进行改造,将TE结构域分别前移至模块5和模块6的末端,同时保留了天然的module7-PCP-TE-linker。在突变菌株fmbJ-M5-TELong和fmbJ-M6-TELong发酵液中,分别检出了截短的新型脂肽,线性脂五肽(C14-15β-NH2FA-Asn-Tyr-Asn-Pro-Glu)和线性脂六肽(C14-17β-NH2FA-Asn-Tyr-Asn-Pro-Glu-Ser)。产生线性脂肽的原因可能是水分子作为亲核试剂捕获acyl-O-TE中间产物并进行水解作用,而水解作用很大程度上取决于肽底物的立体化学和空间特征[22]。同时,我们还在突变菌株发酵液中检测出了截短的环状脂五肽[C14-15β-NH2FA(Asn-Tyr-Asn-Pro-Glu)]和环状脂六肽[C12-14,16-17β-NH2FA(Asn-Tyr-Asn-Pro-Glu-Ser)]。这与环脂肽 Plipastatin的NRPS复合酶系中TE结构域的易位结果相一致[21],Bacillomycin D合成酶系中TE结构域能够识别新合成酶系的末端底物氨基酸:谷氨酸(Glu)和丝氨酸(Ser),催化它们与脂肪酸链的β-NH2形成酰胺键,从而生成环状脂五肽和脂六肽。同时也表明了相较于高选择性的A结构域和C结构域[12,23],TE结构域具有一定的底物耐受性。研究表明,硫酯酶TE的体外表达可有效催化Skyllamycin B 底物类似物进行环化反应[24],Surugamide生物合成途径中的硫酯酶SurE更是能够环化来自2条不同NRPS装配线的结构不相似的底物,体现了SurE广泛的底物耐受性[25],本文结果与之相同。上述结果表明,硫酯酶TE前移后,Bacillomycin D合成酶仍能够合成新型脂肽,TE结构域的底物耐受性则可能是新NRPS合成酶系仍保留脂肽合成功能的原因之一。
除了TE结构域的底物耐受性,本研究还从蛋白质相互作用角度探讨了硫酯酶易位后Bacillomycin D合成酶系仍具有脂肽合成功能的原因。对M5-PCP-TE、M6-PCP-TE和M7-PCP-TE这3个预测结构模型进行分析比较,原始合成酶末端的M7-PCP-TE之间的疏水相互作用与前人研究基本一致[9];硫酯酶TE易位后,PCP-TE双结构域的相对构象和蛋白间相互作用都发生变化,但α2PCP与TE结构域lid regions(α5TE/α6TE)之间的疏水作用力一直存在,是维系PCP-TE双结构域三维结构和功能的主要作用力,也可能是新NRPS合成酶系仍保留脂肽合成功能的原因之一。研究表明原本与C结构域相互作用的PCP结构域在新NRPS装配线中不能有效地与TE结构域相互作用[26],而本研究中模块5和模块6中PCP结构域均能与TE结构域建立起有效的通讯,合成新型脂肽。此外,本研究还观察到硫酯酶TE易位缩小了PCP-TE之间的疏水作用界面,对NRPS合成酶复杂空间构象造成了较大程度的干扰,这也从侧面解释了硫酯酶易位后的Bacillomycin D合成酶合成大环产物的能力下降。研究人员对此提出了交换单元Xus[27]和XUCs[28]的概念,精准定位交换接头的保守基序[19,29],以保持受体和供体单位之间的蛋白质相互作用,最小化链延长期间复杂构象变化的干扰,并保持聚酮合成酶系PKS和非核糖体肽合成酶系NRPS的整体结构[30]。
Bacillomycin D具有两亲性,能有效抑制病原真菌,破坏其脂质双分子层,导致细胞膜的通透性、流动性和完整性发生变化,从而使细胞质成分外泄,最终导致细胞死亡[31]。本研究中,硫酯酶易位后产生的Bacillomycin D衍生物,并不能完全抑制黄曲霉的生长,除了新型衍生物产量低外,推测原因还可能是:一方面为线性脂五肽和线性脂六肽的空间结构较为灵活,未经环化作用将其限制在生物活性构象中,而与线性脂肽相比,环状脂肽具有更高的特异性和亲和力,其蛋白水解抗性也更强[32];另一方面为丝氨酸和苏氨酸都是极性氨基酸,硫酯酶TE前移后,环肽的空间构象发生较大变化,天然Bacillomycin D的亲水面遭到破坏,而亲水区负责与细胞膜的磷脂相结合[33],关系着环肽的生物活性,所以环状脂五肽和脂六肽无法完全破坏黄曲霉菌孢子和菌丝体的细胞壁及细胞膜[1]。
综上所述,本研究将Bacillomycin D合成酶系末端的TE结构域分别前移至模块5和模块6末端,新NRPS合成酶系仍能合成Bacillomycin D类似物,TE结构域的底物耐受性与其易位前后PCP-TE之间一直存在的疏水相互作用是新NRPS合成酶系仍保留脂肽合成功能的原因,可围绕TE结构域进一步探索合成酶结构修饰与新型抗菌脂肽的合成及构效规律。
猜你喜欢
肝博士(2022年3期)2022-06-30
科学(2020年2期)2020-08-24
山东化工(2020年4期)2020-03-30
分析科学学报(2019年2期)2019-04-25
天然产物研究与开发(2019年1期)2019-03-01
物理化学学报(2015年7期)2015-12-30
电源技术(2015年10期)2015-08-01
医学研究杂志(2015年6期)2015-07-01
天然产物研究与开发(2015年2期)2015-01-11
郑州大学学报(理学版)(2013年3期)2013-03-11
