
伪狂犬活/灭活病毒接种小鼠诱导免疫效力及差异因子比较
2023-02-02 11:33章晨昕于晓明侯立婷张元鹏乔绪稳侯继波黄克和郑其升陈瑾
南京农业大学学报 2023年1期
章晨昕,于晓明,侯立婷,张元鹏,乔绪稳,侯继波,黄克和,郑其升,陈瑾*
(1.南京农业大学动物医学院,江苏 南京,210095;2.江苏省农业科学院动物免疫工程研究所/国家兽用生物制品工程技术研究中心,江苏 南京,210014;3.江苏省食品质量安全重点实验室-省部共建国家重点实验室培育基地,江苏 南京,210014)
伪狂犬病是由伪狂犬病病毒(Pseudorabiesvirus,PRV)感染引起的一种急性传染病,可感染猪、鼠、犬、牛、羊和兔等多种哺乳动物[1]。伪狂犬病多宿主传播及高致病性给各国养殖业带来很大威胁,其作为重要的繁殖障碍类疾病,已被列入国家疫病净化中长期规划[2]。
疫苗免疫是防控伪狂犬病最有效的方式之一,目前商品化伪狂犬病灭活疫苗和活疫苗在国内外广泛应用,为猪场提供了强有力保护。与灭活疫苗相比,活疫苗可刺激机体产生更强劲的免疫应答,促进机体更早地产生抗体,且抗体持续时间长,能够为机体提供更有效的免疫保护[3]。但活疫苗普遍存在散毒及毒力返强等安全隐患,且伪狂犬病毒具有嗜神经性,活疫苗免疫后,疫苗毒会在免疫动物的三叉神经处潜伏感染,导致利用活疫苗免疫的猪场一直处于隐性感染的状态,影响猪群健康,也不利于该病的净化[4]。灭活疫苗安全性高,不会引发散毒,也不会带来潜伏感染的问题。但其免疫原性差,不能诱导细胞毒性 T淋巴细胞(CTL)反应,一般需要高剂量、多次免疫,且有致敏风险[5]。因此,研究改进灭活疫苗的免疫效力,使其能激发机体产生更强劲免疫应答,甚至达到活疫苗免疫保护效力水平,将有利于疫病的安全防控。
2018年8月非洲猪瘟首次传入我国,疫情迅速大范围蔓延,对我国养猪业造成极大的威胁,防控形势异常严峻[6]。利用猪作为实验动物开展相关试验难度较大,利用小鼠作为实验动物可以更好控制外源病毒干扰,且组内和组间差异较小可以更好控制变量[7]。Egawa等[8]利用Miyazaki-Bali/2007(PRV-MB)和PRV-Samal-24E两种毒株滴鼻感染9周龄的BALB/c小鼠,通过测定病毒载量以及监测肺部感染情况,以组织病理学和免疫组化学成功建立小鼠PRV感染模型;臧素芳等[9]利用1×105TCID50PRV疫苗经皮下注射免疫小鼠建立伪狂犬人工感染模型,为进一步研究PRV感染小鼠后的天然免疫机制提供试验依据。因此,本研究利用PRV Bratha-K61株,建立PRV人工感染小鼠模型;通过评估不同滴度PRV活病毒感染小鼠后的存活率,探究活/灭活病毒接种小鼠后其体内抗体水平差异及差异显著的细胞因子,从而确定PRV感染安全剂量以及筛选最佳感染方式,建立体内评价动物模型,为后续免疫增强剂的筛选和增强机制研究提供理论基础。
1 材料与方法
1.1 材料
PRV Bratha-K61株由国家兽用生物制品工程技术中心保存。6~8周龄SPF级BALB/c小鼠购自扬州大学比较医学中心。Angle Gene PRV gB、IL-6、IL-1β、TNF-α试剂盒和IFN-β ELISA 试剂盒均购自南京奥青生物公司。噻唑蓝(MTT)、植物血凝素(PHA)购自北京索莱宝生物科技有限公司;二甲基亚砜(DMSO)购自美国Sigma-Aldrich公司。
1.2 PRV Bartha-K61疫苗株灭活
实验室保存的PRV Bartha-K61株经猪睾丸细胞(ST)扩增后收获,于96孔培养板中测定病毒滴度。分别用甲醛和二乙烯亚胺(BEI)灭活病毒。将适量甲醛加入待灭活病毒EP管中,使甲醛终含量为0.2%(体积分数),充分混匀后放入振荡培养箱中,37 ℃、180 r·min-1孵育24 h;将环化BEI加入待灭活病毒EP管中,调整其含量到0.05%(体积分数),充分混匀后放入振荡培养箱中,30 ℃、100 r·min-1孵育48 h[10]。灭活病毒后进行灭活效果检测,ST细胞连续盲传2代,观察细胞不出现病变,即灭活成功。
1.3 小鼠感染模型安全剂量及最佳感染方式筛选
不同滴度的PRV Bratha-K61株活病毒(1×106、5×105、1×105、5×104和2.5×104TCID50·mL-1)经皮下注射、肌肉注射和腹腔注射(1 mL)感染小鼠;连续观察7 d,统计各组小鼠存活率,确定活病毒的感染安全剂量;用安全剂量的活病毒/灭活病毒接种小鼠7 d后检测各组小鼠血清抗体及细胞因子水平,筛选最佳感染途径。
1.4 活病毒和灭活病毒接种小鼠早期免疫差异因子的检测
将最佳感染剂量(5×104TCID50)的PRV Bartha-K61活病毒和灭活病毒(分别用甲醛和BEI灭活)按照最佳感染途径(皮下)接种小鼠,并设空白对照组,每组8只。小鼠接种病毒7 d后,从眼球静脉丛采血,分离血清,冷冻保存,待测。
1.4.1 抗体水平检测在抗体检测酶标板中先加入100 μL待测样品稀释液,再加入4 μL待检血清,振荡混匀,放入25 ℃温箱孵育30 min;配制的新鲜洗涤液(PBST)并洗涤酶标板4次;配制含30 g·L-1BSA的PBST,加入0.2%山羊抗鼠HRP-IgG,充分混匀,每孔加入100 μL混合液,放入37 ℃温箱孵育40 min;用洗涤液重复洗版4次;每孔依次加入50 μL显色液A和显色液B,轻轻振荡混匀,放入37 ℃温箱避光显色 10 min,再加入50 μL终止液后,以空白孔调零,450 nm波长测定各个孔吸光值(D450)。
1.4.2 免疫相关细胞因子水平的检测取待检血清样本,按照Angle Gene检测试剂盒说明书测定血清中相关细胞因子(IL-6、IL-1β、TNF-α和IFN-β)水平。
1.4.3 淋巴细胞增殖试验小鼠接种病毒7 d后,各试验组随机选取3只,分离脾脏,研磨过滤去除红细胞后,利用含10%胎牛血清、100 IU·min-1青霉素和100 μg·mL-1链霉素的RPMI1640培养基制备脾细胞悬液。将脾细胞悬液加入96孔板中,用100 μL灭活PRV抗原刺激(灭活前滴度为5×105TCID50·mL-1,PRV加0.2%甲醛在37 ℃温箱摇床灭活48 h);设PHA刺激的阳性对照组和细胞对照组。37 ℃、5% CO2培养44 h后,每孔加入20 μL 5 mg·mL-1MTT,继续培养4 h,离心,加入DMSO避光振荡,测量D570值。
淋巴细胞刺激指数(SI)=(刺激孔D570值-细胞对照D570值)/(未刺激孔D570值-细胞对照D570值)。
1.5 数据处理与统计分析
利用SPSS 20.0软件的two-ways ANOVA进行方差分析,用t测验法检验各试验组间细胞因子浓度差异。采用GraphPad Prism 5绘图。
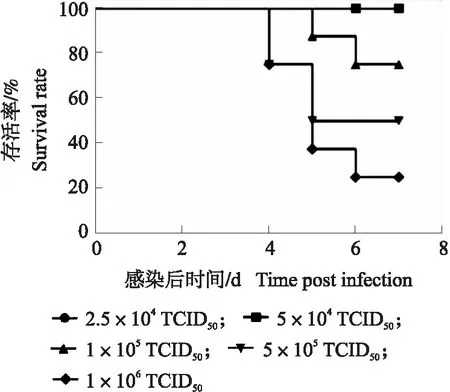
图1 PRV活病毒皮下注射感染小鼠的存活率Fig.1 Survival rate of mice infected with PRV Bratha-K61 by subcutaneous infection
2 结果与分析
2.1 筛选最佳感染剂量和感染途径
将不同滴度的PRV Bratha-K61活病毒分别通过皮下、肌肉和腹腔注射的方式感染小鼠,每天观察小鼠感染情况,7 d后计算小鼠存活率。结果显示(图1),感染剂量为1×106TCID50组小鼠感染后4 d开始出现死亡;感染剂量为5×105和 1×105TCID50组小鼠感染后4 d开始出现脱毛、瘙痒等症状,5 d开始出现死亡;免疫后 7 d,感染剂量为1×106TCID50组小鼠存活率为20%;5×105TCID50组小鼠存活率为50%;1×105TCID50组小鼠存活率为75%;5×104TCID50组小鼠和2.5×104TCID50组小鼠存活率均为100%。
从表1可知:皮下注射时,2.5×104和5×104TCID50病毒滴度为小鼠感染的安全剂量;而腹腔注射和肌肉注射组小鼠在病毒剂量为2.5×104TCID50时存活率仅0和25%。选择感染剂量为2.5×104TCID50分别经皮下注射活病毒和甲醛灭活、BEI灭活PRV Bratha-K61。7 d后,3组存活率均为100%,但激活机体血清抗体水平较低,活/灭活病毒组间抗体水平差异不显著,如继续降低病毒浓度无法激活机体足够抗体水平,不利于后续试验开展。为建立小鼠伪狂犬人工感染模型,探究活/灭活病毒接种小鼠早期免疫相关因子差异,需要存活率达到100%,故确定5×105TCID50·mL-1为最佳感染剂量,皮下注射为最佳感染方式。

表1 PRV活病毒不同注射方式免疫后小鼠的存活率Table 1 Survival rate of mice infected by different immunization methods of live PRV %
2.2 活/灭活病毒接种7 d后小鼠血清细胞因子和抗体水平比较
2.2.1 血清细胞因子水平比较利用Angle Gene ELISA试剂盒检测各组血清TNF-α、IL-1β、IFN-β和IL-6水平(图2)。甲醛灭活组、BEI灭活组、活病毒组和空白对照组TNF-α平均水平分别为57.7、56.4、75.9和47.5 pg·L-1。统计分析表明,甲醛灭活组与BEI灭活组组间血清TNF-α差异不显著(P>0.05),活病毒组与甲醛/BEI灭活组间血清TNF-α水平差异极显著(P<0.001);甲醛/BEI灭活组组间血清IL-1β水平差异不显著(P>0.05),活病毒组与甲醛/BEI灭活组间血清IL-1β水平均差异显著(P<0.05)。甲醛灭活组、BEI灭活组、活病毒组和空白对照组血清IFN-β平均水平分别为263.9、283.4、296.6和236.6 pg·L-1。统计分析表明,甲醛/BEI灭活组组间血清IFN-β水平差异不显著(P>0.05),活病毒组与甲醛/BEI灭活组间血清IFN-β水平差异极显著(P<0.01)。以上结果表明,活病毒/灭活病毒接种后,小鼠血清中TNF-α、IL-1β 和IFN-β水平差异显著,IL-6水平差异不显著,但不同灭活方式对抗体水平和细胞因子水平无显著影响(P>0.05)。

图2 PRV活病毒与灭活病毒接种后(7 d)小鼠血清细胞因子水平比较Fig.2 Comparison of serum cytokine levels in mice inoculated with live and inactivated PRV(7 d) F-inactivated:甲醛灭活组Formaldehyde inactivated inoculated;BEI inactivated:二乙酰亚胺灭活组BEI inactivated inoculated;Live virus:Bratha-K61活病毒接种组Bratha-K61 live virus inoculated;Control:空白对照组Blank control. *P<0.05,**P<0.01,***P<0.001. 下同The same below.
2.2.2 血清抗体水平比较从图3可知:甲醛灭活组、BEI灭活组、活病毒组和空白对照组D450平均值分别为0.24、0.29、0.42和0.10。统计学分析表明,甲醛灭活组与BEI灭活组抗体效价差异不显著(P>0.05),活病毒组与BEI灭活组抗体效价差异显著(P<0.05),活病毒组与甲醛灭活组抗体效价差异极显著(P<0.01)。结果表明,活病毒和灭活病毒接种小鼠后的血清PRV 特异性 gB抗体(IgG)水平存在显著差异,且活病毒组抗体水平显著高于灭活病毒组。

图4 病毒接种7 d后小鼠脾脏淋巴细胞 刺激指数(SI)的比较Fig.4 Comparison of spleen lymphocyte stimulation index(SI)in mice inoculated with live and inactivated PRV
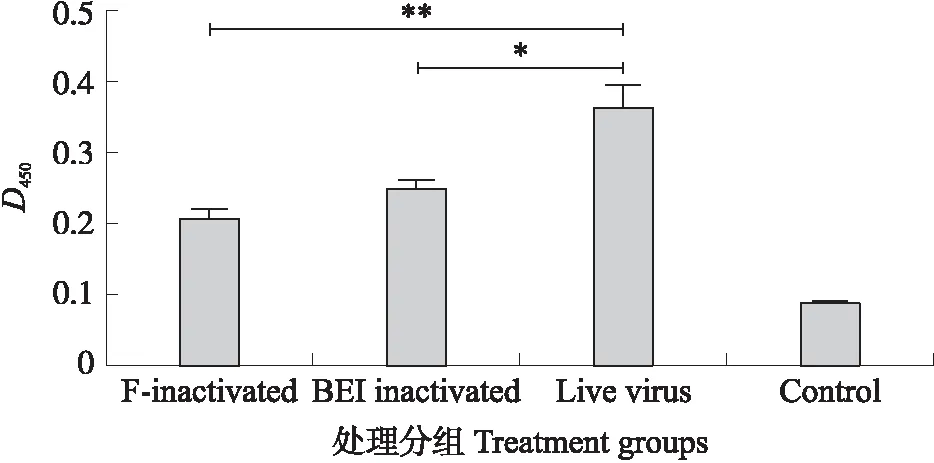
图3 PRV活病毒/灭活病毒接种7 d后小鼠血清 PRV gB抗体(IgG)水平比较Fig.3 Comparison of serum PRV gB antibody(IgG)levels in mice inoculated with live and inactivated PRV
2.3 活/灭活病毒接种7 d后小鼠脾脏淋巴细胞增殖刺激指数(SI)比较
淋巴细胞刺激指数(SI)可以初步反映细胞免疫活化情况。各组SI值结果见图4。对照组SI值设为1,甲醛灭活组、BEI灭活组、活病毒组SI平均值分别为1.75、2.08和5.75。统计学分析表明,甲醛灭活组与BEI灭活组SI值差异不显著(P>0.05),活病毒组与甲醛/BEI灭活组组间SI值均差异极显著(P<0.001)。结果表明活病毒感染与灭活病毒接种小鼠相比能够更好地激活小鼠细胞免疫,且不同灭活方式之间差异不显著。
3 讨论
为了更好地改进伪狂犬灭活疫苗的免疫效力,提高灭活疫苗保护效力。本试验利用小鼠建立伪狂犬病毒的人工感染模型,为伪狂犬灭活病毒免疫增强剂的筛选和效力评价建立高效平台,为后续伪狂犬灭活疫苗效力的改进和其他灭活疫苗免疫增强剂的研发奠定基础。
不同剂量病毒经不同途径进入机体后,在激活免疫应答方面有极大差异[11]。本研究显示,同等剂量下不同免疫途径对小鼠存活率的影响较大,与皮下注射5×104TCID50PRV(存活率100%)相比,肌肉注射(存活率为12.5%和25%)和腹腔注射(存活率均为0)小鼠存活率显著下降。范克伟等[12]利用皮下腹股沟注射不同剂量PRV病毒感染小鼠,发现接种高剂量病毒的小鼠体重下降更快,更早出现临床症状且症状更明显,死亡率更高,伪狂犬病毒具有典型的神经嗜性,腹腔、肌肉组织中以及免疫细胞、神经和血管都非常丰富,有利于抗原的扩散,导致发病和死亡。本试验在保证免疫源性足够,且小鼠存活率达到较高水平下,成功建立了活/灭活抗原的感染和差异评价模型。活病毒/灭活病毒进入机体后,在激活免疫应答方面有极大差异[13]。Sander等[14]发现灭活菌与活菌感染小鼠后,检测小鼠血清相关细胞因子水平,其中TNF-α、IL-1β和IFN-β水平差异显著,并得出差异因子与生存能力相关模式识别受体相关的结论。本试验中,活病毒与灭活病毒免疫小鼠7 d后,检测小鼠血清中与先天免疫相关细胞因子水平,其中活病毒组小鼠血清中TNF-α、IL-1β和IFN-β水平显著高于灭活病毒组(P<0.05)。这3类细胞因子均与vita-PAMP引起的差异因子一致,而与vita-PAMP 无关的炎性细胞因子 IL-6 的水平差异不显著。甲醛通过破坏病毒蛋白及核酸达到灭活病毒目的;BEI只破坏病毒核酸而不破坏蛋白质。BEI灭活与甲醛灭活相比可更好保持病毒免疫原性,但试验结果显示不同灭活方式间细胞因子水平差异不显著。vita-PAMP作为一种仅存在于活性微生物中的危险信号,进入机体后使得先天免疫细胞能够区分活的和死的微生物,并仅对活微生物动员产生高效免疫反应[15]。本试验中,当活病毒进入机体后能够高效激活机体天然免疫应答,这与vita-PAMP的存在密切相关[15-16]。这为后续在伪狂犬灭活苗中免疫增强剂研制中提供了研究方向。
PRV gB蛋白是PRV最保守的蛋白之一,可产生有效保护易感动物感染的中和抗体,具有较高的特异性和敏感性,也是国际贸易检测PRV的指定方法[17-18]。本试验比较活病毒和灭活病毒免疫小鼠后其体内短期血清gB抗体水平的结果显示,活病毒组小鼠产生抗体水平显著高于灭活病毒组(P<0.01)。试验结果为后续评价免疫增强剂对伪狂犬灭活疫苗抗体水平的影响提供了有的效评价方法。
淋巴细胞增殖和分化是机体免疫应答过程的一个重要阶段,检测淋巴细胞增殖水平是反映细胞免疫应答的常用方法之一。淋巴细胞刺激指数(SI)升高提示机体产生更强劲的细胞免疫反应[19]。本研究证明活病毒组SI极显著高于灭活病毒组。免疫增强剂发挥作用可通过SI值显著增加来反映细胞免疫水平的提升。这说明活病毒和灭活病毒进入机体后,活病毒能够更高效的激活细胞免疫应答。类似活病毒免疫后产生的细胞免疫应答,是更有效和期待的免疫应答方向。
综上,本研究成功建立了安全的活/灭活病毒小鼠感染模型,并确定了活/灭活病毒进入机体后,活病毒会刺激机体产生的更高水平抗体和相关免疫因子,并且与vita-PAMPs分子所引起的免疫差异一致。这为后续伪狂犬灭活疫苗免疫效力的改进提供了指导方向。
猜你喜欢
中国病理生理杂志(2022年11期)2022-12-03
建材发展导向(2021年11期)2021-07-28
生物学通报(2021年4期)2021-03-16
当代水产(2020年10期)2020-03-17
江苏安全生产(2020年1期)2020-03-16
当代水产(2019年8期)2019-10-12
中华神经外科疾病研究杂志(2018年4期)2018-08-20
中华肺部疾病杂志(电子版)(2018年6期)2018-03-11
当代医药论丛(2017年1期)2017-11-30
中央民族大学学报(自然科学版)(2014年3期)2014-06-09
