
朱砂根叶绿体全基因组解析及系统发育分析
2023-02-02 06:46:54刘雄伟刘畅曾宪法杨小英俸婷婷赵杰宏周英
生物技术通报 2023年1期
刘雄伟 刘畅 曾宪法 杨小英 俸婷婷 赵杰宏 周英
(贵州中医药大学药学院 药食两用资源应用与开发研究中心,贵阳 550025)
朱砂根为紫金牛属植物朱砂根(Ardisia crenata)的干燥根或根茎,至今有多年的民间药用历史,其含有三萜类、黄酮类、香豆素类等多种化学成分,具有清热解毒、散瘀止痛功效,主要用于治疗咽喉肿痛、扁桃体炎等病症。对本草著作中紫金牛属药物进行基原考,《本草纲目》及《图考》记载的“朱砂根”是指叶背红色的红凉伞,其鲜根断面皮部有多个朱砂色小点[1]。红凉伞(Ardisia crenatavar.bicolor)与朱砂根在植株外形上无大差别,仅叶背、花梗、花芬及花瓣均带紫红色而有所不同,Flora of China中红凉伞已合并至朱砂根,但仅仅是从形态学方面判断红凉伞为朱砂根的变种尚缺乏科学的依据。对朱砂根和红凉伞进行性状鉴别、显微鉴别、DNA条形码、遗传变异等研究[2-5],表明朱砂根和红凉伞亲缘关系较近,但已有DNA 条形码序列分辨率较低,并不能很好地明确物种界限,未能理清朱砂根和红凉伞的进化演变,对其起源、扩张和迁移的过程,种内遗传变异空间分布与环境条件关系等尚不清楚,其系统发育关系尚不明朗。
叶绿体基因组序列的结构、大小和顺序相对稳定,并且突变率较缓慢[6],是系统发育研究和物种鉴定有效工具之一[7],其突变热点区域已经被用于物种及物种以下水平(品种)亲缘关系研究[8-9]。随着第二代测序技术(next generation sequencing)的兴起,叶绿体基因组在解析植物系统进化发育关系研究中得到广泛应用,如青牛胆[10]、白头翁[11]、牛蒡[12]、罂粟[13]等已经完成了叶绿体基因组分析,对其系统发育和亲缘关系进行研究。针对紫金牛属物种系统发育关系尚不清楚的问题,对紫金牛属百两金Ardisia crispa、走马胎Ardisia gigantifolia、纽子果Ardisia polysticta、朱砂根A.crenata、紫金牛Ardisia japonica、虎舌红Ardisia mamillata、雪下红Ardisia villosa、皱叶紫金牛Ardisia bullata等进行叶绿体基因组测序,解析其物种进化关系,同时发现紫金牛属与长柱金牛属亲缘关系较近[14-17]。但关于紫金牛属归属于紫金牛科还是报春花科尚且存在争议[14-17],同时2020 版《中国药典》认为药材朱砂根为紫金牛科植物朱砂根的干燥根。植物化学方面研究表明紫金牛科与报春花科在亲缘关系上是相近的科(http://www.iplant.cn/info/Primulaceae),将紫金牛属合并到报春花科,这种分类方法是否科学合理尚缺乏科学依据。
因此,本研究对不同地区朱砂根A.crenata、红凉伞A.crenatavar.bicolor3 个紫金牛属物种的新鲜叶片进行叶绿体基因组测序,解析其叶绿体基因组序列与结构特征,筛选种间高变异序列;同时下载紫金牛科和报春花科代表性叶绿体全基因组序列信息,构建系统进化树,进而为紫金牛属药用植物的分类鉴定、遗传进化关系及资源开发利用等提供基础。
1 材料与方法
1.1 材料
朱砂根(贵州)和红凉伞新鲜叶片采自贵州中医药大学种质资源圃,朱砂根(江西)新鲜叶片采自江西武功山,经贵州中医药大学魏升华教授鉴定为朱砂根A.crenata、红凉伞A.crenatavar.bicolor。取新鲜健康叶片,用无菌水冲洗干净,晾干后置于-80℃冰箱保存。通过NCBI 数据库(https://www.ncbi.nlm.nih.gov/)检索报春花目的紫金牛科和报春花科的23 个叶绿体全基因组序列信息,共检索到紫金牛科紫金牛属14 个种、蜡烛果属1 个种、酸藤子属2 个种、铁仔属3 个种及报春花科报春花属3 个种的信息(表1)。

表1 紫金牛科和报春花科物种的叶绿体基因组GenBank 登记号Table 1 Chloroplast GenBank accession numbers of the species in Myrsinaceae and Primulaceae
1.2 方法
1.2.1 基因组DNA 提取与测序 利用植物基因组DNA 提取试剂盒(TIANGEN,北京)提取总DNA,0.8%琼脂糖电泳检测DNA 样品是否有降解及杂质,Nanodrop 分光光度计检测样品浓度及纯度。检测合格的DNA 样品根据Illumian DNA 文库构建流程,构建插入片段大小为350 bp 的双末端测序文库。对质检合格测序文库,利用Illumina Novaseq6000 高通量测序平台进行测序。测序结果经CASAVA 软件进行碱基识别(base calling)后转化为原始测序序列(raw reads),利用NGS QC ToolKit 软件对raw reads 进行质控,过滤去除接头和低质量序列,得到高质量序列(clean reads)。
1.2.2 叶绿体全基因组序列组装、拼接与注释 采用SPAdes v.3.15.2 软件(http://cab.spbu.ru/software/spades/)对叶绿体基因组序列进行拼接组装,利用CpGAVAS[18]和ORF Finder 对叶绿体基因组进行注释。对注释的初步结果,运用Blastn 和Blastp 的方法与已报道的近缘物种叶绿体基因组编码蛋白和rRNA 进行比对,验证结果的准确性并进行修正。tRNA 的注释采用ARWEN[19]软件进行注释,生成tRNA 二级结构图。利用OGDRAW[20]软件绘制叶绿体基因组环状结构图。
1.2.3 SSR 和重复序列分析 利用MISA 在线软件(https://webblast.ipk-gatersleben.de/misa/index.php)分析微卫星序列(SSR)[21],参数阈值设置为:1、2、3、4、5、6,核苷酸参数为8、4、4、3、3、3,且两个SSRs 之间的距离不小于100 bp,并对SSRs 的类型、数量等进行分析。利用REPuter(https://bibis erv.cebitec.uni-bielefeld.de/reputer)在线工具对长重复序列进行分析[22]。
1.2.4 叶绿体全基因组比较及共线性分析 利用mVISTA 软 件(https://genome.lbl.gov/vista/mvista/submit.shtml)在Shuffle-LAGAN(全局比对模式)下[23],以本研究新获得的朱砂根叶绿体基因组为参考,对紫金牛属其它物种叶绿体全基因组序列同源性进行比较分析;采用 Geneious10.2.2 软件中Mauve多重基因组比对法检测紫金牛物种的叶绿体基因组的重排和共线性[24],包括月月红A.faberi、虎舌红A.mamillata、雪下红A.villosa、走马胎A.gigantifolia和酸苔菜A.solanacea。
1.2.5 系统进化分析 选择已发表的紫金牛科叶绿体基因组为内类群和与紫金牛科亲缘关系较近的3个报春花科植物钟花报春P.sikkimensis、鄂报春P.obconica、腾冲灯台报春P.chrysochlora为外类群,构建最大似然(maximum likelihood,ML)系统发育树。利用MAFFT 7(https://mafft.cbrc.jp/alignment/server/)在线软件对其叶绿体基因组进行全基因组比对[25-26],采用IQ-TREE 2.0.5(http://www.iqtree.org/)软件[27],基于最大似然法,通过比较不同样本叶绿体基因组序列的差异构建系统发育树,25 条序列共包含8 867 个简约性信息位点(parsimonyinformative),6 759 个自裔位点(singleton sites),136 494 个不变位点(constant sites),构树参数设置为:-s .fasta -st DNA -m TEST -bb 1 000 -alrt 1 000-abayes,优构树模型为:TVM+F+I+G4。
2 结果
2.1 朱砂根及红凉伞叶绿体基因组基本特征
叶绿体基因组图谱显示朱砂根及红凉伞叶绿体基因组均为双链环形结构(图1),包括大单拷贝区(large single copy region,LSC)、小单拷贝区(One small single copy region,SSC)、反向重复区(reverse repeat regions,IRa)和反向重复区(reverse repeat regions,IRb)4 个边界区,4 个边界区长度大小基本一致,GC 含量无差异(表2)。相比于LSC 和SSC区,IR 区的GC 含量最高,可能是由于IR 区含有高GC 含量的RNA 基因造成。
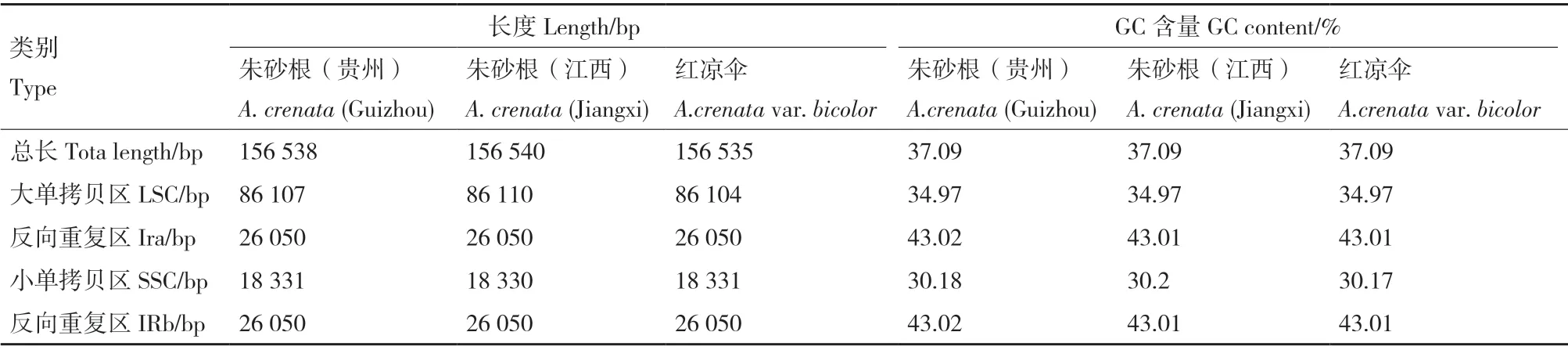
表2 朱砂根及红凉伞叶绿体基因组基本信息统计Table 2 Basic information statistics of chloroplast genomes between A.crenata and A.crenata var.bicolor
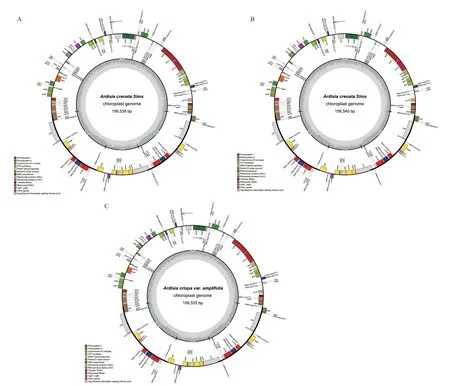
图1 叶绿体基因组图谱Fig.1 Chloroplast genome map
2.2 朱砂根及红凉伞叶绿体基因组功能及分类
在朱砂根和红凉伞叶绿体全基因组序列中注释得到132 个基因(表3),包括87 个蛋白编码基因、37 个tRNA 基因和8 个rRNA 基因,主要分为光合作用基因、复制基因、未知功能基因以及乙酰辅酶A 羧化酶亚基(accD)、成熟酶基因(matK)、其他基因。有 10 个蛋白质编码基因ndhB、rpl2、rpl23、rps12、rps7、ycf15、ycf2,7 个 tRNA 编码基因及4个 rRNA 编码基因位于 IR 区。

表3 朱砂根及红凉伞叶绿体基因组分类Table 3 Chloroplast genome classification between A.crenata and A.crenata var.bicolor
对叶绿体基因内含子进行统计分析发现,共有15 个基因含有内含子,其中ycf3和clpP蛋白质编码基因含有 2 个内含子。
2.3 朱砂根及红凉伞叶绿体基因组长重复序列及简单重复序列分析
利用REPuter 在线工具对的长重复序列进行分析。朱砂根和红凉伞均有50 个长重复序列,包括正向重复(forward,F)、回文重复(palindrome,P)、反 向重复(reverse,R)和互补重复(complement,C),但其类型个数稍有不同(表4)。简单重复序列(simple sequence repeat,SSRs)分析表明(表5),朱砂根共有207 个SSRs 位点,红凉伞有205 个SSRs位点,均为未发现六核苷酸重复基序。

表4 朱砂根及红凉伞叶绿体基因组长重复序列分析Table 4 Analysis of long repeats of chloroplast genome between A.crenata and A.crenata var.bicolor
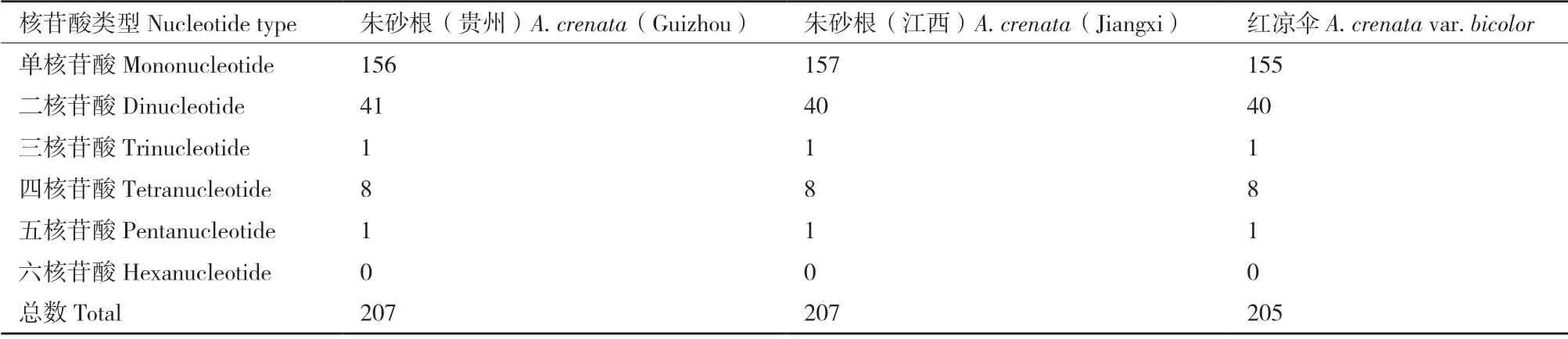
表5 朱砂根及红凉伞叶绿体基因组SSR 分析Table 5 Analysis of SSR of chloroplast genome between A.crenata and A.crenata var.bicolor
2.4 朱砂根及红凉伞叶绿体基因组IR-LSC/SSC 区的收缩和扩张分析
边界分析显示紫金牛属叶绿体基因组虽然在序列长度、基因组成以及GC 含量等方面相对稳定保守,但4 个边界区的过渡区域却存在有多样性(图2)。紫金牛属8 个物种的IRb-LSC 边界均位于rps19的编码区。走马胎、月月红、雪下红、酸苔菜、虎红舌、朱砂根(贵州)IRb-SSC 边界均位于ndhF编码区,朱砂根(江西)距ndhF编码区5 bp。紫金牛属8 个物种IRa-SSC 边界,除了红凉伞位于ndhF编码区外,其他种均位于ycf1基因内。走马胎、月月红、雪下红IRa-LSC 边界均位于rpl2和tRNA 基因间区内,酸苔菜、红凉伞、朱砂根(江西)位于rpl2和trnH间区内,朱砂根(贵州)位于rps1和trnH基因编码区,虎红舌位于rps19和trnH基因编码区。

图2 叶绿体基因组IR 与 SC 边界Fig.2 Chloroplast genome IR and SC boundary
2.5 朱砂根及红凉伞叶绿体基因组序列变异分析
以朱砂根(贵州)作为参考基因组,采用mVISTA 软件检测基因重排和倒位的全局比对模式(Shuffle-LAGAN),对红凉伞、朱砂根(江西)、月月红、虎红舌、雪下红、走马胎和酸苔菜叶绿体全基因组序列同源性进行比较研究(图3)。结果表明,紫金牛属物种叶绿体基因组LSC、IRa、SSC、IRb四个区域排列顺序较为一致、保守性较高,非基因编码区变异程度较高,其中 SSC 区的变异程度最高;基因编码区差异不明显。紫金牛属叶绿体基因组中psaB、psbA、psbC、matK、atpA、atpB、rpoC2、rpoC1、ropB、ndhD、ndhF、ndhK、rbcL、accD、ropA等基因的编码区存在差异。此外,朱砂根与红凉伞叶绿体基因组相比,psbA、matK、rpoC2、ropB、ndhK、accD、ndhF、ndhD、ndhH及ycf1等基因的编码区存在差异,这些位点为红凉伞的分子鉴定提供了新的位点资源。
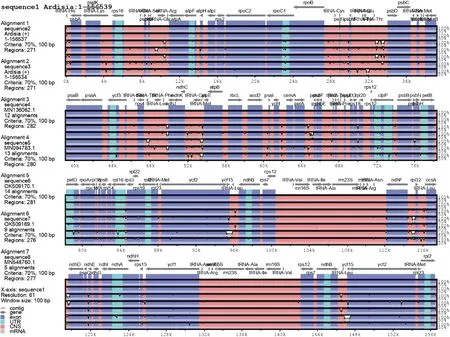
图3 叶绿体基因组全局比对图Fig.3 Chloroplast genome global alignment map
2.6 朱砂根及红凉伞叶绿体基因组共线性分析
采用 Mauve 多重基因组比对法检测红凉伞、朱砂根(贵州)、朱砂根(江西)、月月红、虎红舌、雪下红、走马胎和酸苔菜8 个物种的叶绿体基因组的重排和共线性。通过多重基因组比对法检测出 8 个物种的叶绿体基因组之间有 5 个局部共线块(locally collinear block,LCB)(图4),这表明8 个物种之间的基因组具有高度的相似性。叶绿体全基因组序列的比对显示,叶绿体基因组之间没有重排或倒置,从叶绿体基因组4 个组分上看,IR 区序列变异最低,SSC 区的变异程度最高。

图4 叶绿体基因组的共线性分析Fig.4 Covariance analysis of the chloroplast genome
2.7 朱砂根及红凉伞系统发育分析
选择已发表的紫金牛科叶绿体基因组为内类群和与紫金牛科亲缘关系较近的3 个报春花科植物钟花报春P.sikkimensis、鄂报春P.obconica、腾冲灯台报春P.chrysochlora为外类群,构建ML 系统发育树。在系统发育树中(图5),可确定为不包括外类群物种的3 个主要类群:蜡烛果属和杜茎山属为一个类群,酸藤子属为一个类群,紫金牛属为一个类群,支持率为100%,酸藤子属和紫金牛属可能是在蜡烛果属和杜茎山属基础上进化的。这与前人研究略有不同,Xie 等[14]认为紫金牛属与长柱金牛属亲缘关系较近。基于系统进化树发现朱砂根与纽子果亲缘关系较近,与已有研究结果一致[14]。进一步分析发现,朱砂根与红凉伞亲缘关系最密切,从分子水平为红凉伞作为朱砂根变种提供了科学解释。系统进化树结果表明,叶绿体基因组构建的系统发育树可用于紫金牛属物种植物的鉴定。
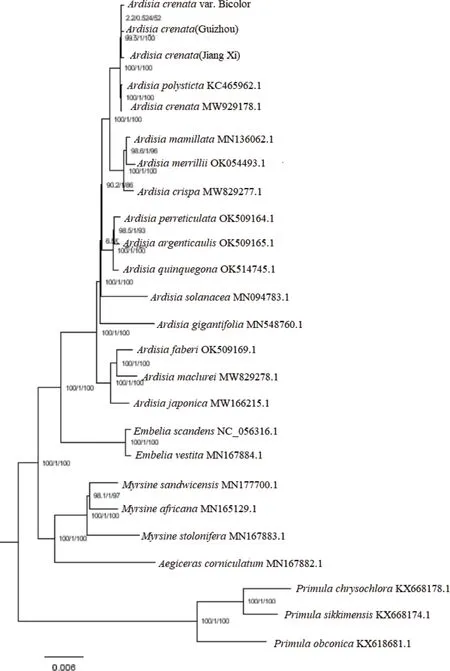
图5 叶绿体基因组序列构建的系统发育树Fig.5 Phylogenetic tree constructed from chloroplast genome sequence
3 讨论
本研究对不同地区朱砂根及变种红凉伞进行叶绿体基因组测序,测序结果表明三者叶绿体基因组均具有典型的环状四分体结构,包括 LSC、SSC、IRa 和IRb 四个区,其序列总长度、CG 含量、基因数量基本一致,且IRs 区序列GC 含量最高。叶绿体基因组包括蛋白编码区和非编码区,编码区包含100-120 类不同的基因,涉及光合作用、复制等作用[28],朱砂根和红凉伞叶绿体全基因组序列中注释得到132 个基因,包括87 个蛋白编码基因、37 个tRNA 基因和8 个rRNA 基因。叶绿体基因组的突变不是随机的,而是存在突变热点区域,这些突变热点区域已经被用于物种及物种以下水平(品种)亲缘关系研究[8-9]。通过对变异位点的分析,发现psaB、psbA、psbC、matK、atpA、atpB、rpoC2、rpoC1、ropB、ndhD、ndhF、ndhK、rbcL、accD、ropA、ccsA、ycf2及ycf1等基因的编码区存在差异,为该属的分子鉴定奠定了基础。
叶绿体基因标记物,如简单重复序列(simple sequence repeats,SSRs)己经被用于基因流、种群分化和遗传多样性研究[29]。本研究从朱砂根叶绿体基因组中共检测到207 个 SSR 位点,SSR 以单核苷酸重复为主,且随着拷贝数目增加,SSR 数量明显减少,朱砂根和红凉伞的SSR 位点类型和数量基本一致,但略有不同,可为朱砂根分子标记开发或基原鉴定提供理论依据。朱砂根叶绿体基因组序列中单核苷酸、二核苷酸、三核苷酸SSR 类型分别以A/T、AT/TA、AAT/ATT 为主,表明朱砂根叶绿体基因组序列中SSRs 主要由polyA 或 polyT 所构成,与已有研究结果结论一致[30]。
Liu 等[31]利用形态特征和matK 分子标记对24种紫金牛属的亲缘关系进行研究,但由于形态特征不稳定和遗传信息不足,仍不能推断紫金牛属物种亲缘关系。而完整的叶绿体基因组序列具有更高的分辨率,可以用于紫金牛属物种鉴定及进化的分析,Xie 等[14]认为紫金牛属与长柱金牛属亲缘关系较近。这与本文的研究结果不同,系统进化树表明紫金牛属可能是在蜡烛果属和杜茎山属基础上进化的,支持率为100%。同时Xie 等[14]认为紫金牛属为报春花科,但亦有说法紫金牛属为紫金牛科。植物化学方面研究表明紫金牛科与报春花科在亲缘关系上相近的科,目前紫金牛属已合并到报春花科(http://www.iplant.cn/info/Primulaceae)。但本研究构建的叶绿体系统发育树,通过支持率可将紫金牛科和报春花科分为两个分支,紫金牛属是否可以划分为报春花科值得进一步商榷,需要进一步结合贝叶斯树及溯祖理论进行分析。
对朱砂根原植物的考证[32],认为红凉伞更符合《本草纲目》记载的朱砂根,吴征镒认为红凉伞与朱砂根在植株外形上无大差别,仅叶背、花梗、花芬及花瓣均带紫红色而有所不同,二者作为两个单独种并不适当,因此将红凉伞作为朱砂根的变种,在最新版的Flora of China中红凉伞已合并至朱砂根。叶绿体基因组系统发育树结果表明,朱砂根与红凉伞亲缘关系最为亲密,在朱砂根的基础上进化,这也为红凉伞作为朱砂根变种提供了科学解释。
4 结论
基于Illumina 测序平台对不同地区朱砂根及变种红凉伞叶绿体全基因组进行测序,psbA、matK、rpoC2、ropB等基因编码区存在差异,为朱砂根和红凉伞的分子鉴定提供了新的位点资源。系统发育表明朱砂根与红凉伞亲缘关系最为亲密,紫金牛属可能是在蜡烛果属和杜茎山属基础上进化,为进一步研究紫金牛属药用植物的分子进化和亲缘关系提供了充分的信息。但需要进一步结合贝叶斯树及溯祖理论探讨紫金牛属与报春花科之间的关系,以明确紫金牛属的划分。
猜你喜欢
英语世界(2023年10期)2023-11-17 09:18:18
今日农业(2021年11期)2021-08-13 08:53:24
科学大众(中学)(2019年3期)2019-05-17 10:04:30
汽车观察(2018年10期)2018-11-06 07:05:26
广西林业科学(2016年1期)2016-03-20 05:33:01
少儿科学周刊·少年版(2015年1期)2015-07-07 17:15:12
遗传(2014年3期)2014-02-28 20:58:49
世界科学(2014年8期)2014-02-28 14:58:31
食品科学(2013年6期)2013-03-11 18:20:13
世界科学(2013年6期)2013-03-11 18:09:33
