
不同稻蟹共作模式下的氮磷平衡及生态经济效益研究
2023-01-19 07:45:32吕明睿高子健张欣雨李艳和宋光同赵玉华
华中农业大学学报(自然科学版) 2023年1期
吕明睿,高子健,张欣雨,李艳和,宋光同,赵玉华
1.华中农业大学水产学院/农业农村部淡水生物繁育重点实验室/长江经济带大宗水生生物产业绿色发展教育部工程研究中心, 武汉 430070;2.安徽省农业科学院水产研究所,合肥230001
我国稻渔共作种养模式的发展历史悠久,但直至近十年才有相对快速的发展,并逐步形成了如今规范、标准、有一定规模的稻渔一体化种养模式[1]。随着共作模式日趋成熟,稻蟹[2]、稻虾[3]等共作模式在全国范围内得到推广,使得稻蟹/虾养殖系统的潜在时空价值和营养结构得到充分利用,为农户带来水稻和水产品的双丰收[4]。这种稻渔共作的生态循环农业模式有着高效的资源利用,良好的经济、生态效益,因而备受农户喜爱。
相关研究表明,稻渔综合种养模式在生产上能够提高田间土壤养分[5],增加氮磷含量,促进水稻生长;相对水稻单作,稻渔共作能有效减少60%以上农药和20%以上化肥的使用,减少环境的污染[6],丰富田间生物多样性[7],并能够充分利用藻类、浮游生物、昆虫等系统内资源[8],减少投入,增产增收;相对于水稻单作,稻虾、稻蟹共作模式总经济产出分别提高了156%和710%[9]。
稻渔共作模式除能提升农户经济收益外,还可改善环境[3,10],如Zhang等[11]研究发现稻虾共作可以增加稻田土壤有机碳的含量;Bashir等[12]研究表明:与水稻单作相比,稻蟹共作不仅可以提高水稻产量、品质及经济效益,还能调控稻田排水中污染物的浓度,保护环境;安辉等[13]发现施用有机肥的稻蟹共作模式可在一定程度上提高稻蟹产量、改善稻米品质。上述研究主要集中于某一种共作模式对生态环境、经济产量或产品品质的探讨,而针对不同稻蟹共作模式下的氮磷平衡及生态、经济效益的对比研究较少。因此,本研究通过对不同稻蟹共作系统中氮磷及浮游生物的变化分析、产物收益比较来寻找最佳的绿色生态的稻蟹共作模式,以期为制定安全优质高效的稻渔共作生产技术规范、探求可持续发展的水产养殖技术提供理论支持。
1 材料与方法
1.1 试验区概况
试验区位于安徽省宿州市泗县丁湖镇水产养殖基地。该地属暖温带季风气候,光照充足、雨量适中、雨热同期,年平均气温17.2 ℃,年平均降雨量1 214 mm[14],养殖区土壤以壤土为主。
1.2 试验设计与田间管理
于试验区设立精养蟹(intensive crab, IC)、稻蟹共作(rice-crab co-culture, RC)、稻虾蟹共作(ricecrayfish-crab co-culture, RCC)3种种养模式,并于2021年2月下旬开始养殖试验,10月下旬收获结束试验。3块养殖田面积均为2.33 hm2,每块田地以“田”字分为4个区域形成处理内的重复。稻、虾、蟹品种分别为金香玉1号(Oryza sativa)、克氏原螯虾(Procambarus clarkii)、中华绒螯蟹(Eriocheir sinen⁃sis)。精养蟹田整田挖掘,形成小型池塘,水深1.5 m,坡比1∶1.5,专养蟹,于2月投放蟹苗,蟹苗规格为120只/kg,投放总量为335 kg,放养密度约为17 250只/hm2;稻蟹田与稻虾蟹田四周挖掘形成回型沟,沟渠面积为0.53 hm2,沟深1.2 m,沟宽7 m,沟渠水深1.5 m,坡比1∶1.5。稻蟹田蟹与稻共作种养,于2月投放蟹苗,蟹苗规格为120只/kg,投放总量为185 kg,蟹放养密度约为9 520只/hm2,6月上旬种植水稻,人工插秧,种植间距为40 cm,于10月下旬干田机械收割;稻虾蟹田蟹、虾和稻共作种养,于4月投放虾苗300 kg,规格为100只/kg,6月投放幼蟹800 kg,规格为20只/kg,虾蟹放养密度分别约为12 870、6 870只/hm2,水稻种植、收割情况与稻蟹田相同。
各养殖田均不施用化肥,每14 d换水1次,每次进排水高度为20 cm。精养田和稻蟹田在3月投喂蟹饲料和冰鲜鱼,蟹饲料1次/6 d,冰鲜鱼1次/3 d,饲料投喂量为蟹质量的5%,鲜鱼投喂量为蟹质量的10%。精养田、稻蟹田和稻虾蟹田在4月开始改投冰鲜鱼、玉米、大豆、青豆,每天投喂,冰鲜鱼日投喂量分别为100、50、50 kg,玉米、大豆、青豆各种养田投喂量相同,分别为20.0、11.7、11.7 kg。
1.3 样品的采集与测定方法
试验开始前对试验田的基本情况进行调查,各养殖田情况基本一致。在种养期间于6月、7月、9月(插秧期、水稻快速生长期和水稻成熟期)分别采集养殖水体、稻田土壤、沟渠底泥、水稻植株、虾、蟹、野杂鱼、幼蟹商品饲料、饲料原料(玉米、青豆、大豆、冰鲜鱼)等样本,测定其氮磷含量、分析浮游生物种类与生物量。具体分析方法如下:
1)水样的采集与测定。每次进水、排水时记录水位变化,计算进排水量,并在进出水口采集水样。于稻田四角分别用1 L采水器、25#浮游生物网于水深0.5 m处采集水样及浮游生物。用采水器采集250 mL水样,-4 ℃保存用于总氮总磷测定;采集500 mL水样加10 mL鲁哥氏液固定,沉降48 h后取浓缩样品,蒸馏水定容至50 mL,添加少量35%甲醛溶液固定用于浮游植物、原生动物和轮虫的定量分析;采集10 L水样经25#浮游生物网过滤,蒸馏水定容至50 mL,加少量35%甲醛溶液固定用于枝角类和桡足类的定量分析;25#浮游生物网采集的浮游生物用35%甲醛溶液固定用于浮游生物定性分析。水体总氮测定采用碱性过硫酸钾消解紫外分光光度法(HJ 636-2012),水体总磷测定采用钼酸铵分光光度法(GB 11893-1989),浮游生物的定性定量分析于显微镜下进行。
2)土壤、底泥样品的采集与测定。于稻田四角用彼得逊采泥器采集底泥,用铁锹采集土壤样品,每次200~500 g。样品混匀后于105 ℃环境下干燥48 h,用粉碎机粉碎、过筛、装袋用于总氮总磷测定。土壤总氮测定采用凯氏法(HJ 717-2014),总磷测定采用钼锑抗比色法(GB 9837-1988)。
3)稻、虾、蟹、野杂鱼及饲料样品的采集与测定。水稻在每块田的每个重复选择1 m2采样框随机取样,清洗后在105 ℃环境下杀青30 min,后降至80 ℃烘干,用粉碎机粉碎、装袋;虾、蟹、野杂鱼于每个试验田随机采样(虾蟹数量大于30只),清洗后在105 ℃环境下烘干,用粉碎机粉碎、装袋;商品饲料、冰鲜鱼、玉米、大豆、青豆等饲料样品采用四分法采样(饲料质量大于200 g),在鼓风干燥机中105 ℃环境下烘干,用粉碎机粉碎、装袋。稻、虾、蟹、野杂鱼及饲料样品总氮的测定采用凯氏定氮法(GB/T 6432-2018),总磷的测定采用矾钼酸铵显色法(GB/T 6437-2018)。
4)数据的收集、计算与分析。日常饲料投喂时,记录饲料的投喂量及种类;虾蟹投放前称量总质量;插秧前记录秧苗总质量;产品收获时,记录稻、虾、蟹、野杂鱼的产量及市场单价;降雨量参考安徽省2021年气候公报[14];大气氮磷沉降数据参考高嵩等[15]、彭秋桐等[16]的报道。
氮磷平衡表示氮磷在环境中的盈余,即整个养殖周期的氮磷输入(饲料、虾苗、蟹苗、稻苗、进水、雨水、大气沉降)与氮磷输出(成虾、成蟹、成稻、野杂鱼、排水)的差值[17];氮磷利用率即氮磷输入与输出的比率[18];根据实际购入成本和收成收入分析经济效益;浮游生物分析采用Shannon-Wiener多样性指数(H')评价不同种养模式下水体污染情况,计算公式如下:
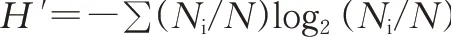
用Microsoft Excel 2016进行数据处理,SPSS Statistics 26进行不同处理间各指标的差异性及相关性的统计分析。
2 结果与分析
2.1 不同种养模式系统氮磷含量变化
相较于试验前,不同种养模式于试验后底泥、土壤的氮磷含量均有不同程度的增加(表1)。在6月,不同种养模式底泥中的总氮含量差异显著(P<0.05),精养蟹模式底泥总氮含量最低,稻虾蟹共作模式最高。相较于6月,9月精养蟹模式底泥的氮、磷含量分别增加了9.69、0.23 g/kg,稻蟹共作模式增加了6.07、0.17 g/kg,稻虾蟹共作模式增加了0.37、0.27 g/kg。相较于试验前,稻蟹共作和稻虾蟹共作模式试验后土壤中总氮分别增加了2.04、3.74 g/kg,总磷分别增加了0.23、0.09 g/kg,种养后期的9月,稻虾蟹共作模式中总氮含量要显著高于稻蟹共作模式(P<0.05)。综合来看,共作模式能有效减少氮磷的沉积,减缓有机质的富集。

表1 不同种养模式下土壤、底泥、水体的氮磷含量Table 1 Nitrogen and phosphorus contents in soil, sediment and water under different culture patterns
从水体氮磷含量来看,精养蟹模式总氮含量种养前后无显著差异(P>0.05),而总磷含量种养后比种养前增加了0.16 mg/L,9月总磷含量显著高于6月(P<0.05);不同共作模式总氮含量相比试验前期有显著性降低(P<0.05),而于9月又显著性升高(P<0.05),均有3.00 mg/L左右的增幅,而试验前后总磷含量则整体呈下降趋势,稻蟹共作模式减少了0.16 mg/L,稻虾蟹共作模式减少了0.08 mg/L,6月与9月水体总磷含量无显著性差异(P>0.05)。水体中总氮、总磷含量变化表现出一定的差异性。
2.2 不同种养模式系统氮平衡及利用
精养模式中,氮输入主要为饲料、虾苗、蟹苗、进水、雨水和大气沉降(图1),3种种养模式下系统氮平衡及利用情况见表2。其氮总输入量为1 234.12 kg/hm2,其中饲料的氮输入量为1 120.97 kg/hm2,占精养模式氮输入总量的90.83%,是输入氮的主要来源,其次为蟹苗,占总输入量的4.01%;氮输出主要为成蟹、野杂鱼、排水,其氮总输出量为203.20 kg/hm2,成蟹、排水的氮输出量的占比分别为84.88%、12.90%。
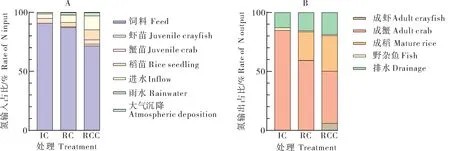
图1 不同种养模式下各指标总氮输入(A)、输出(B)占比Fig. 1 Ratio of TN input(A) and output (B) under different culture patterns
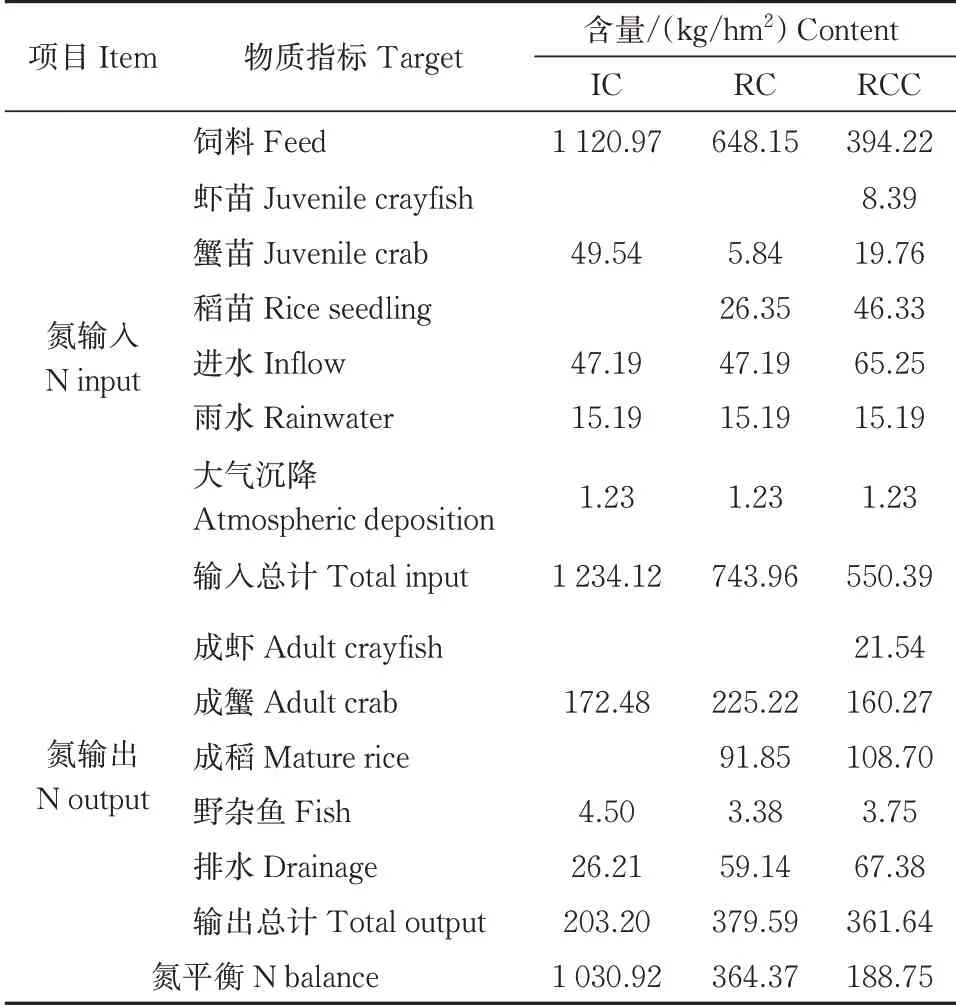
表2 不同种养模式下系统氮平衡及利用Table 2 Nitrogen balance and utilization under different culture patterns
稻蟹共作模式中,氮输入、输出较精养模式增加了水稻的投入和产出。其氮总输入量为743.96 kg/hm2,较精养模式减少了39.72%,其中饲料、进水是输入氮的主要来源,占氮输入总量的87.12%、6.34%;氮总输出量为379.59 kg/hm2,较精养模式增加了86.81%,其中成蟹、成稻的氮输出量合计占氮输出总量的83.53%。
稻虾蟹共作模式中,氮输入输出较精养模式增加了稻苗、虾苗的投入和产出,其氮总输入量为550.39 kg/hm2,较精养模式减少了55.40%,氮输入的主要途径为饲料、进水,分别占氮输入总量的71.63%、11.86%;氮总输出量为361.64 kg/hm2,较精养模式增加了77.97%,成虾、成蟹、成稻等产品合计占输出总量的80.34%。
精养、稻蟹共作、稻虾蟹共作模式的氮平衡分别为1 030.92、364.37、188.75 kg/hm2,稻蟹共作模式和稻虾蟹共作模式相较于精养模式分别减少了64.66%、81.69%。从氮的盈余上看,稻蟹和稻虾蟹共作模式要明显低于精养模式,表明稻蟹共作和稻虾蟹共作减少了系统中氮素的盈余,稻虾蟹共作模式效果更加明显。在氮的利用率上,稻蟹共作和稻虾蟹共作模式分别为48.98%、65.71%,与精养蟹模式的16.47%相比,分别提升了32.51%、49.24%。
2.3 不同种养模式系统磷平衡及利用
3种种养模式系统磷平衡及利用情况如图2所示,3种种养模式的磷输入输出指标与氮的情况类似(表3)。精养模式中,磷总输入量为185.47 kg/hm2,饲料仍是主要输入来源,其磷输入量为182.33 kg/hm2,占精养模式磷输入总量的98.31%,其次为进水,占总输入量的0.60%;磷输出主要为成蟹、野杂鱼、排水,其磷总输出量为14.12 kg/hm2,成蟹、排水的磷输出量的占比为44.19%、36.26%。

图2 不同种养模式下各指标总磷输入(A)、输出(B)占比Fig. 2 Ratio of TP input (A) and output (B) under different culture patterns
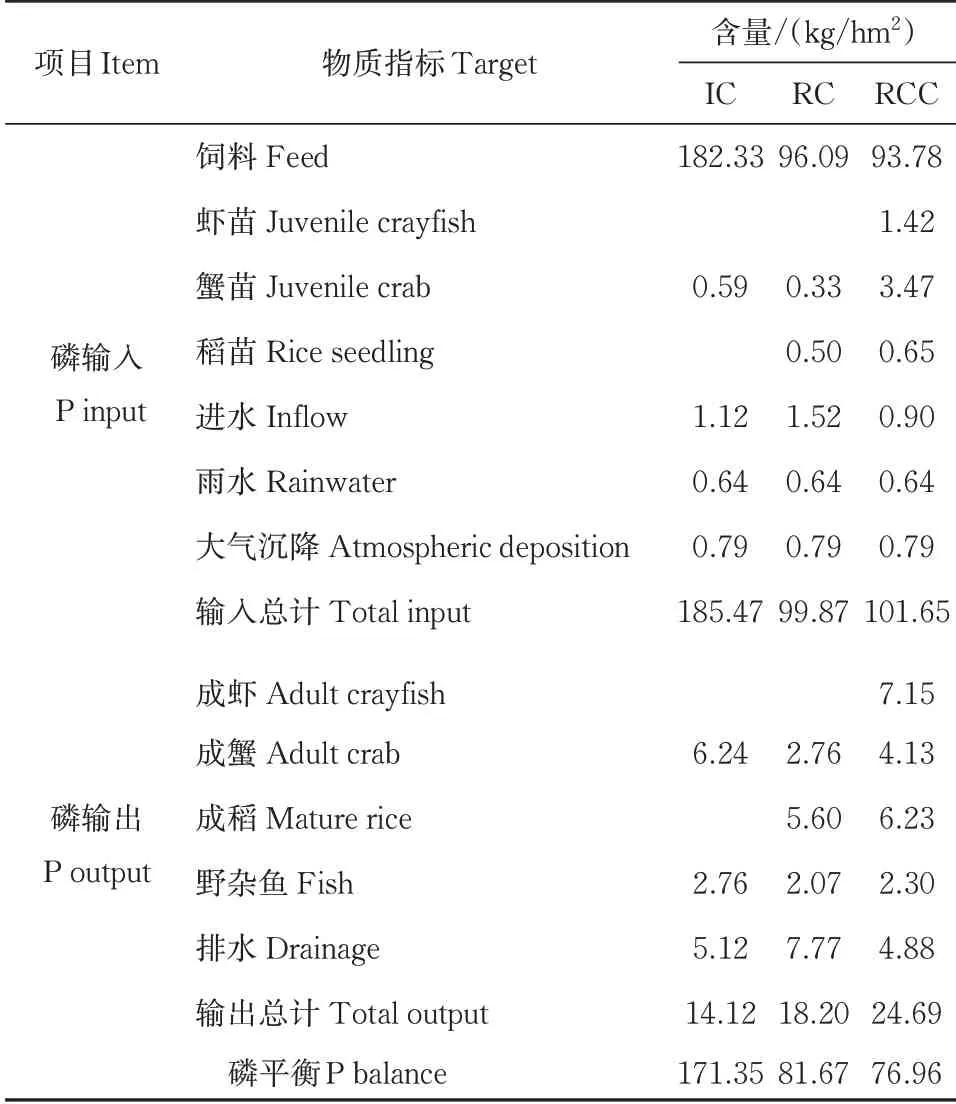
表3 不同种养模式下系统磷平衡及利用Table 3 Phosphorus balance and utilization under different culture patterns
稻蟹共作模式中磷总输入量为99.87 kg/hm2,较精养模式减少了45.23%,其中饲料、进水占磷输入总量的96.22%、1.52%;磷总输出量为18.20 kg/hm2,较精养模式增加了28.90%,其中成蟹、成稻作为主要产品的磷输出量合计为8.36 kg/hm2,占磷输出总量的百分比为45.93%。
稻虾蟹共作模式中,磷输入输出较精养模式增加了秧苗、虾苗的投入和产出,其磷总输入量为101.65 kg/hm2,较精养模式减少了45.19%,与稻蟹共作模式差异不大,磷输入的主要途径为饲料、虾苗、蟹苗,分别占磷输入总量的92.26%、1.40%、3.41%;磷总输出量为24.69 kg/hm2,较精养模式增加了74.86%,成虾、成蟹、成稻的磷输出量合计17.51 kg/hm2,占稻虾蟹共作模式磷输出总量的百分比为70.92%。稻虾蟹共作模式中,磷输入输出较精养模式增加了稻苗、虾苗的投入和产出,其磷总输入量为101.65 kg/hm2,较精养模式减少了45.19%,与稻蟹共作模式差异不大,磷输入的主要途径为饲料、虾苗、蟹苗,分别占磷输入总量的92.26%、1.40%、3.41%;磷总输出量为24.69 kg/hm2,较精养模式增加了74.86%,成虾、成蟹、成稻的磷输出量合计达17.51 kg/hm2,占稻虾蟹池磷输出总量的百分比为70.92%。
3种种养模式的磷平衡分别为精养模式171.35 kg/hm2,稻蟹共作模式81.67 kg/hm2,稻虾蟹共作模式76.96 kg/hm2,稻蟹共作和稻虾蟹共作相较于精养模式分别减少了52.34%、55.09%。从磷的盈余上看,稻蟹和稻虾蟹共作模式要显著低于精养模式,表明稻蟹共作和稻虾蟹共作模式减少了系统中磷素的盈余,稻虾蟹共作模式效果更加明显。在磷的利用率上,稻蟹共作和稻虾蟹共作模式分别为18.22%、24.29%,与精养蟹模式的7.61%相比,分别提升了10.61%、16.68%。
2.4 不同种养模式浮游生物生物量及多样性
在6月,精养蟹模式检出浮游植物8种,浮游动物15种;稻蟹共作模式检出浮游植物16种,浮游动物14种;稻虾蟹共作模式检出浮游植物12种,浮游动物17种。稻蟹共作、稻虾蟹共作浮游植物种类要明显高于精养蟹模式,浮游动物种类则无明显差异。7月,精养蟹、稻蟹共作、稻虾蟹共作模式分别检出浮游植物15、18、27种,浮游动物14、19、15种。相较于6月,浮游植物种类都有所增加,稻虾蟹共作模式尤为突出,浮游动物种类无明显差异。 9月,3种种养模式分别检出浮游植物31、37、28种,浮游动物27、24、37种,相较于7月,除稻虾蟹共作模式浮游植物种类无明显变化外,其余浮游生物种类均有较大提升。总体来看,试验前后3种种养模式浮游生物种类均呈现上升趋势,精养蟹、稻蟹共作、稻虾蟹共作模式分别增加了35、31、36种,稻虾蟹共作模式在浮游生物种类总量和增长量上,均为最多。
除7月份稻虾蟹共作模式中浮游动物生物量显著高于精养模式(P<0.05)外,其余各组浮游生物生物量差异均不显著(P>0.05),但是稻虾蟹共作模式于7月和9月的浮游生物生物量均明显高于精养模式(表4)。精养蟹、稻蟹共作、稻虾蟹共作模式浮游植物在6、7、9月的Shannon-Wiener多样性指数变化范围分别为1.14~2.48、2.00~2.42、1.98~2.42(表4),均值分别为1.92、2.20、2.20;浮游动物在6、7、9月的Shannon-Wiener多样性指数变化范围分别为2.99~3.79、2.48~3.36、2.82~3.98(表4),均值分别为3.27、2.92、3.39。稻虾蟹共作模式的浮游植物、浮游动物的Shannon-Wiener多样性指数均较高。就多样性数值变化趋势看,稻蟹共作、稻虾蟹共作Shan‑non-Wiener多样性指数增长程度更大,且在9月整体高于精养蟹模式,更有利于生物多样性。
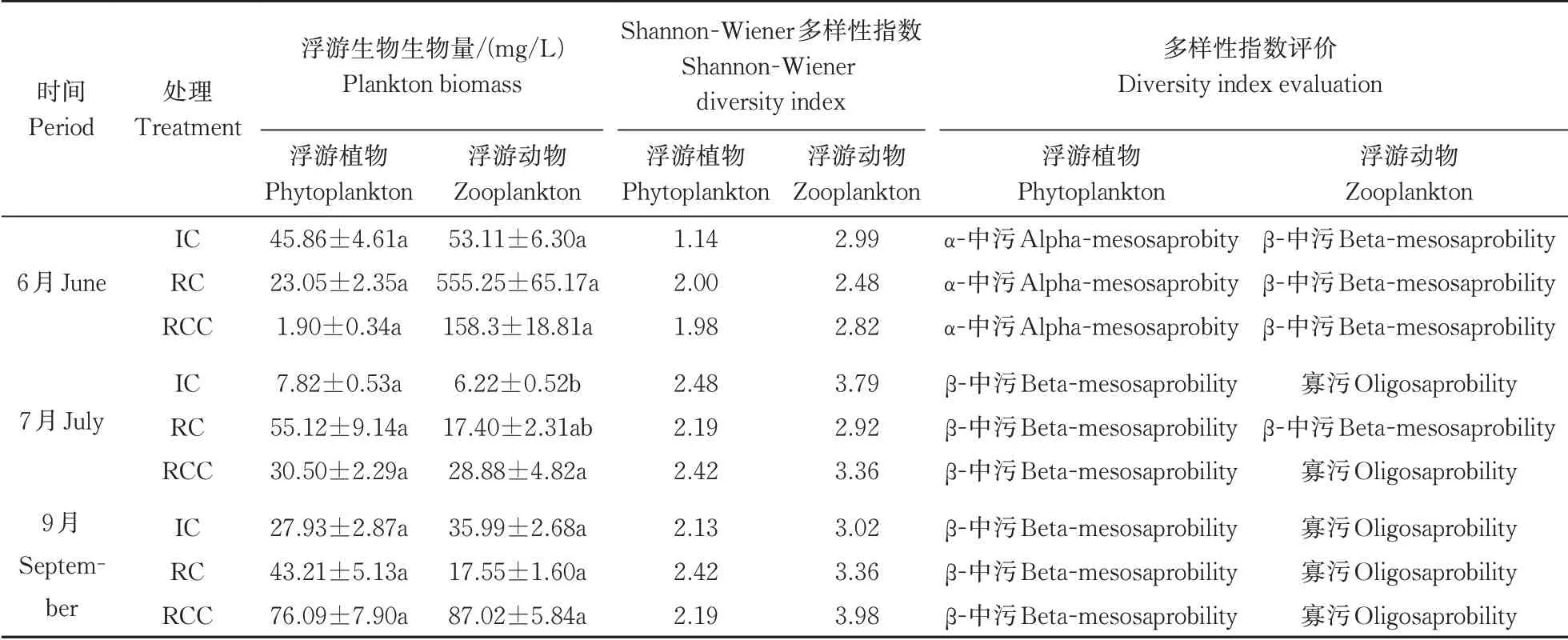
表4 不同种养模式下浮游生物多样性指数Table 4 Plankton diversity index in different culture patterns
根据多样性指数评价标准综合评价,精养蟹、稻蟹共作、稻虾蟹共作模式的浮游植物多样性指数评价结果是6月均为α-中污、7月和9月均为β-中污;浮游动物多样性指数评价结果分别对应β-中污、寡污、寡污(除稻蟹共作模式7月对应β-中污),3种种养模式并无显著差异(P>0.05)。
2.5 不同种养模式的经济效益
不同种养模式的收入(成蟹、成虾、稻米、杂鱼)、支出(虾苗、蟹苗、人工、机械、水电、消毒改底、增氧设施、网具、杂鱼螺蛳、玉米黄豆、地租)情况见图3。稻蟹和稻虾蟹共作模式的支出成本分别为52 790、50 858元/hm2,相较于精养模式分别减少了7.17%、10.57%,减少的费用主要体现在蟹苗、饲料的投入,稻虾蟹共作模式还增加了虾苗的投入。精养蟹模式蟹收益105 150元/hm2,稻蟹共作模式稻、蟹收益合计90 128元/hm2,稻虾蟹共作模式稻、虾、蟹收益合计99 142元/hm2。蟹作为主要的收益产品,其收入分别占精养蟹、稻蟹共作、稻虾蟹共作3种模式总收入的93.51%、78.48%、66.47%,精养蟹模式显著高于后两者。稻蟹共作、稻虾蟹共作的生态米收入分别占其总收入的15.69%、16.51%。稻蟹、稻虾蟹共作模式净收入分别为42 918、53 090元/hm2,相较于精养蟹模式分别减少了22.78%、4.48%。3种种养模式中,精养模式的净收入最高,稻虾蟹共作的净收入次之,其中稻虾蟹共作模式与精养模式差异不大。
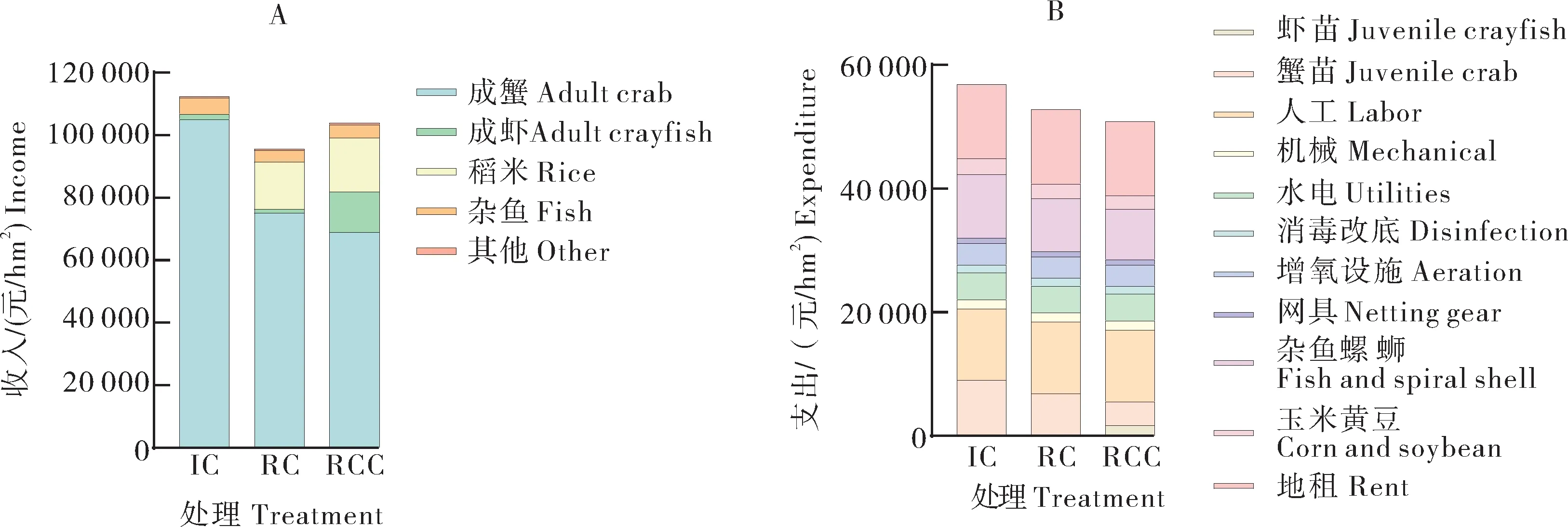
图3 不同种养模式的收入(A)、支出(B)明细Fig. 3 Income (A) and expenditure (B) breakdown of different culture patterns
3 讨论
3.1 氮磷含量、平衡及利用率
邝雪梅等[19]、佀国涵等[20]对稻田生态种养的研究结果表明,共作模式能够改善土壤理化性质,提高土壤通气性,增加总氮、总磷含量。与之相似,本试验期间,精养、稻蟹共作、稻虾蟹共作模式中底泥的总氮、总磷含量均有不同程度升高,但稻蟹、稻虾蟹共作模式的总体沉积量(总输入量-总输出量)要少于精养模式。已有研究多与水稻单作对比,其共作模式的虾蟹活动(与水稻单作相比)更有助于改善土壤结构[19-20]。共作模式下虾蟹的活动范围一般趋向于在沟渠边缘,而不是稻田中间[21],而精养模式中蟹的活动范围是整个区域,因此,共作模式下的土壤氮磷含量虽有增加,但无底泥显著。为了更好地满足蟹的生长,养殖户往往会投入过量的饲料,本试验精养模式饲料投喂量接近共作模式的2倍,而蟹的投放量未达到2倍的关系,因此会造成一定量的未被摄食的饲料残留在底泥中,同时蟹的排泄物也会增加底泥的总氮、总磷含量。虾蟹活动也会进一步影响底泥氮磷含量,精养模式由于更大的养殖密度与更广的活动范围而加剧了底泥氮磷的沉积。此外,精养模式中的水草的生长与腐败亦会影响氮磷的含量。因此,在今后的研究中需要将精养模式下水草对氮磷含量的影响考虑在内。
养殖系统中氮磷含量受饲料投喂、生物投入、生物吸收代谢、换水、降雨、大气沉降等多种因素的影响而呈现出动态波动。张云杰等[22]研究发现共作模式水体氮磷含量出现先增后减的趋势,与本研究不尽相同。本试验中,水体总磷降低可能与水稻生长代谢活动有关,因为植株正常生长时的生理活动会促使根系吸收水体中的氮磷,同时还能利用其聚集和繁殖的微生物来促进氮磷分解净化[23];而水体总氮含量的增加则与张云杰等[22]的研究结果不符,可能与共作模式下浮游植物密度大、生物量多,更易形成有氧环境,促进硝化作用,造成底质氮素向水体释放有关,上述原因还有待进一步研究验证。
氮磷输入与支出间的平衡状况是衡量环境影响、土壤肥力、种养系统养分可持续利用的重要指标[17],体现了氮磷在种养环境系统中的盈余情况。本研究结果表明,饲料是氮磷输入的主要途径,以稻、虾、蟹产品产出为氮磷主要输出方式,其中饲料磷输入超90%,饲料氮输入超70%,产品氮输出超80%,产品磷输出近50%,这与张丁月等[24]、Si等[25]的研究结果相似。精养、稻蟹共作、稻虾蟹共作模式均出现氮磷的盈余,稻蟹共作和稻虾蟹共作在氮磷的盈余量上都要显著低于精养模式,体现出共作模式能缓解养殖系统环境的富营养化的功能,且以稻虾蟹共作模式效果最佳。上述结果可能是以下原因造成:其一,虾蟹可以动植物碎屑、浮游生物为食,稻蟹共作、稻虾蟹共作模式浮游生物量大且多样,可减少饲料输入,因此,这2种模式饲料氮磷的输入量都仅为精养模式的一半或更少;其二,稻蟹共作、稻虾蟹共作引入稻、虾生产环节,有效增加了氮磷输出。可见共作模式通过减少饲料输入、提高产品输出能有效降低养分盈余,减缓环境富营养化。
氮磷的平衡也反映了养分的利用情况。本试验中,稻蟹共作和稻虾蟹共作的氮利用率和磷利用率均要显著高于精养模式。这是因为精养模式中饲料的过量投入使得大部分以饲料途径输入的氮磷直接以残饵形式或间接以排泄物形式沉积在底泥中;共作模式能有效提高水稻对氮磷的吸收[26-27],相较于精养蟹,共作模式增加了水稻培育的生产环节,而水稻生长需利用氮磷元素,这提高了共作模式对氮磷等养分的利用率。磷易与矿质离子形成沉淀[28],且水稻、水产品本身所固定的磷量并不大[29],同时,水稻作为主要的磷输出指标,其产量较低,使得3种种养模式磷利用率普遍低于氮利用率。而对比稻蟹共作、稻虾蟹共作,后者的氮磷利用率均高于前者,表现为更少的氮磷输入、更多的氮磷输出、更佳的氮磷循环利用,更有利于可持续发展。
3.2 浮游生物生物量及多样性变化
浮游生物既会随季节变化发生群落结构改变,也会因环境因子的影响发生变化。本研究中,3种种养模式的浮游生物多样性均随时间推移呈现上升的趋势,浮游植物多样性均低于浮游动物,这与Zhou等[30]的研究结果相符。
精养模式初期浮游植物多样性不及稻蟹、稻虾蟹共作模式,但在试验后期快速上升,并与稻虾蟹共作模式的生物多样性接近。这可能与有机质能促进浮游植物的生长[31]有关。精养模式初期水体培肥需要时间,加上蟹总量较多,可能是造成浮游植物种类相对较少及浮游生物量相对较低的原因;而且在6、7月正值夏季高温,蓝藻门可快速繁殖成为优势种[32],不利于其他浮游植物的生长。水稻生长后期,共作模式中水稻对营养盐的吸收利用促使其氮磷大量消耗,而精养模式对氮磷的累积效果更明显,营养盐含量高,呈现出更佳的浮游植物长势。
温度是影响浮游动物生物量的重要指标[33],枝角类、桡足类在高温时更易生长,水稻种植期间,精养模式无水稻遮荫,温度较高,其枝角类桡足类要明显高于稻蟹共作、稻虾蟹共作。浮游植物与浮游动物之间存在捕食关系,因此,本研究中浮游动物与浮游植物生物多样性呈现相同的变化趋势是合理的。综合分析,稻虾蟹共作模式的浮游生物群落组成较稻蟹共作、精养模式更复杂多样,稳定性更高,能够更好地抵抗不良因素的影响。
3.3 稻、虾、蟹产量及经济效益
稻蟹共作、稻虾蟹共作模式的蟹产量分别为536、494 kg/hm2,相较于精养模式分别减少了215、257 kg/hm2,稻虾蟹共作模式中的虾产量为644 kg/hm2,相较于前人研究中的344 kg/hm2[3]有不小的提升。水稻产量则相对较少,仅为1 502、1 717 kg/hm2,虽显著低于水稻单作的产量,但与厉宝仙等[34]在稻鳖虾共作的研究中水稻产量下降情况相符。未施肥的生态稻米品质有所提升,价格相对普通稻米也更高,对稻蟹、稻虾蟹共作模式有着较大的收入贡献。
本研究中,稻蟹、稻虾蟹共作模式净收入分别为42 918、53 090元/hm2,相较于精养蟹模式分别减少了12 661、2 489元/hm2。蟹作为主要的收益来源,其产量的降低无疑减少了共作模式的收益,但稻虾蟹共作模式中增加的利用养殖时间空档的虾的养殖收成弥补了稻蟹种养模式在收益上的不足,使其与精养模式的经济效益差距减小。
综上可知,相较于精养蟹模式,共作模式能有效降低水体总磷含量、减少氮磷的沉积、改善浮游生物群落结构,同时能够减少饲料投喂,降低氮磷的盈余,提高氮磷的持续循环利用。共作模式中的稻虾蟹共作模式的效应尤为突出,且相比精养模式,在经济效益上也有不错的表现,是一种优质、可持续发展的生态种养模式。
致谢:感谢安徽沱湖宏腾养殖有限公司在试验中给予的大力支持;感谢安徽省农业科学院水产研究所徐斌老师在样品采集中给予的无私帮助。
猜你喜欢
新农民(2023年18期)2023-07-06 08:27:20
作文小学高年级(2022年9期)2022-10-17 07:20:42
天津农林科技(2022年2期)2022-04-19 10:48:16
——百援精养家族培训大会成功举办
汽车维修与保养(2022年9期)2022-03-07 06:50:16
东坡赤壁诗词(2022年1期)2022-02-25 07:28:07
科教创新与实践(2021年38期)2021-11-14 07:09:39
当代水产(2019年10期)2019-12-18 05:28:56
当代水产(2019年1期)2019-05-16 02:41:58
吉林农业(2018年15期)2018-01-18 18:47:18
黑龙江水产(2017年5期)2017-03-02 12:36:45
