
黑潮对东海网采浮游植物群落组成与分布的影响*
2023-01-17 05:00孙振皓江志兵朱元励曾江宁陈全震陈建芳
海洋与湖沼 2023年1期
孙振皓 江志兵 詹 琳 朱元励 曾江宁 陈全震 陈建芳
黑潮对东海网采浮游植物群落组成与分布的影响*
孙振皓1, 2, 4江志兵1, 2, 3, 4①詹 琳1朱元励1, 2, 4曾江宁1, 2, 4陈全震1陈建芳1
(1. 自然资源部第二海洋研究所 自然资源部海洋生态系统动力学重力实验室 浙江杭州 310012; 2. 浙江省近海海洋工程环境与生态安全重点实验室 浙江杭州 310012; 3. 浙江省海洋科学院 海洋空间资源管理技术重点实验室 浙江杭州 310012; 4. 自然资源部长三角海洋生态环境科学野外观测研究站 浙江舟山 316021)
黑潮入侵深刻影响东海生态环境, 但对其如何影响浮游植物群落组成与分布仍知之甚少。为此, 于2011年四季对东海(26°~33°N, 121°~128°E)共164个站位进行浮游植物拖网采集和环境因子测定, 分析了浮游植物丰度和优势种组成及其对黑潮入侵的响应。调查共检出浮游植物9门509种(含变种、变型和未定种), 其中硅藻305种、甲藻154种, 蓝藻、定鞭藻、金藻、裸藻、绿藻、隐藻和黄藻种类数较少。秋季浮游植物细胞丰度最高(30 496.91×103cells/m3), 高值区位于黑潮与长江冲淡水交汇形成的锋面处; 夏季次之(28 911.28×103cells/m3), 高值区分布与秋季相似; 春季较少(19 180.76×103cells/m3), 高值区位于舟山群岛东南部; 冬季最低(472.36×103cells/m3), 高值区位于东海南部。冬季受黑潮表层水入侵影响, 主要优势种为铁氏束毛藻(); 春、夏季主要优势种为骨条藻(spp.), 由于黑潮次表层水势力增强, 入侵东海并携带丰富的磷酸盐, 加之长江冲淡水输入大量无机氮, 夏季浮游植物总丰度高于春季; 秋季主要优势种为柔弱伪菱形藻(), 受黑潮表层水势力增强和次表层水影响, 磷酸盐浓度最高, 浮游植物总丰度在四季中最高。种类数表现为秋季>冬季>夏季>春季, 水平分布为由近岸向外海递增, 种类数高值位于黑潮影响区域。相似性分析和多维尺度分析显示, 浮游植物群落组成时空差异明显。由此可见, 黑潮输送的磷酸盐促进了浮游植物的生长, 且其携带的暖水种增加了浮游植物的种类多样性, 使东海浮游植物群落组成表现出明显的季节、空间变化。
东海; 黑潮; 浮游植物; 群落组成; 季节变化
东海生态环境主要受沿岸流系(沿岸流和长江冲淡水)和黑潮流系(黑潮和台湾暖流)影响。其中, 黑潮是北赤道洋流的北支流, 起源于菲律宾海东部, 经台湾岛东北部和琉球群岛西南部流入东海(Qu, 2016)。黑潮每年入侵东海大陆架的总量约1.3~1.4 Sv, 这为东海提供了充足的热量和丰富的磷酸盐(Zhang, 2007; 杨德周等, 2017; Wang, 2018; Liu, 2021)。浮游植物生长一般需按Redfield比值(氮磷比N/P=16)吸收营养盐, 而长江等河流入海输送的氮磷营养盐比值约为100, 导致东海浮游植物的生长受到磷限制(Harrison, 1990; Gong, 1996; Chen, 2004; Li, 2009; Tseng, 2014)。黑潮入侵带来的磷酸盐可能改善东海存在的磷限制问题, 并可能影响该区域浮游植物的丰度与群落构成。此外, 高温、高盐的黑潮有着丰富的热带暖水“种库”(杨清良等, 1999; 刘东艳等, 2000; 孙振皓等, 2021), 也可能显著影响了东海的浮游植物种类组成。黑潮入侵东海陆架的程度与流量大小因季风等变化存在明显季节差异(Tsui, 2012; Wu, 2014)。总体而言, 黑潮表层水入侵东海陆架秋冬季强、春夏季弱, 黑潮次表层水入侵东海陆架夏季最强、冬季最弱(Yang, 2012, 2018)。以往相关研究集中在长江口等近岸海域(顾新根等, 1995; 吴玉霖等, 2004; 栾青杉等, 2007; 王云龙等, 2008; 李俊龙等, 2013; 江志兵等, 2014; Jiang, 2015), 较少覆盖整个东海(Guo, 2014; 周茜茜等, 2015), 而且大都关注了长江冲淡水等沿岸水团对浮游植物群落组成与分布的影响, 鲜有黑潮入侵影响东海浮游植物群落季节、空间变化的系统报道。为此, 本文于2011年冬、春、夏、秋季拖网采集了东海(26°~33°N, 121°~128°E)的浮游植物, 探究其群落组成和分布特征, 并结合黑潮的时空变化, 探讨黑潮对东海浮游植物群落组成与分布的影响。研究结果有助于了解东海浮游植物群落时空分布格局与环流的关系。
1 数据与方法
1.1 调查海域与站位
于2011年冬(12月至翌年1月)、春(3~4月)、夏(7月)、秋(10月)季对东海进行浮游植物拖网调查。春、夏、秋、冬季调查分别设34、39、44、47个站位。除黑潮及其分支(台湾暖流)影响外, 东海还受到长江冲淡水和沿岸流影响。具体站位布设与环流示意见图1。

图1 2011年东海调查站位与环流示意图(a: 冬半年, b: 夏半年)
1.2 样品采集与方法
浮游植物样品采集按照《海洋调查规范》(GB/T 12763.6-2007)进行(中华人民共和国国家质量监督检验检疫总局等, 2008), 用浅水III型浮游生物网(筛绢孔径76 µm)自底层垂直拖曳至表层, 使用流量计(HYDRO-BIOS公司, 德国)记录滤水量。样品在现场用5%中性甲醛固定, 带回实验室静置24 h, 经浓缩或稀释后使用Leica DM6B显微镜进行形态观察、物种鉴定和计数(杨世民等, 2006)。使用CTD(SBE 917 Plus, Seabird, Sea Bird公司, 美国)采水器采集标准层海水测定理化参数, 其中温度、盐度和水深通过CTD传感器测得, 溶解无机氮(dissolved inorganic nitrogen, DIN)、溶解无机磷(dissolved inorganic phosphorus, DIP)和溶解硅酸盐(dissolved silicate, DSi)的采集、测定和数据分析均按照《海洋调查规范》(GB/T 12763.4-2007)进行。
1.3 数据统计与分析
选取受黑潮影响较强的E03、E04、P08和P09站位作为黑潮影响区、受黑潮影响较弱的D01、D02、D03和D04站位作为对照区, 并根据林更铭等(2007)对浮游植物暖水种的划分, 统计各季节黑潮影响区和对照区浮游植物暖水种的种类数、磷酸盐浓度和N/P (附表1)。
将各季节浮游植物细胞丰度占比前10的物种定为该季节的优势种。浮游植物细胞丰度经log(+1)转换后建立Bray-Curtis相似性聚类分析, 基于聚类分析的相似度矩阵对浮游植物群落分布进行多维尺度分析(multidimensional scaling, MDS)与相似性分析(analysis of similarities, ANOSIM)。MDS和ANOSIM均使用PRIMER 6软件(Clarke, 2006)。不同季节环境因子的非参数检验(K-W检验)以及浮游植物优势种与环境因子的斯皮尔曼相关性分析通过SPSS 26.0进行。选取优势种, 使用Canoco 5软件进行典范对应分析(canonical correspondence analysis, CCA), 浮游植物细胞丰度和环境参数均经log(+1)转换。使用Ocean Data View (ODV)软件绘制图件。
2 结果与分析
2.1 理化环境条件
东海表层温度、盐度、DIN、DIP和DSi均存在显著的季节差异(<0.05)。冬季黑潮水入侵导致东海东南部表层海水盐度超过34、温度高于20 °C (图2e), 东海西北部受低温、低盐的沿岸水控制, 使得调查区域出现由西北向南、东南递增的温度、盐度梯度(图2a)。春季东海受黑潮分支影响, 有一条自南向北的暖水舌贯穿调查区域(图2b), 在舟山群岛以东海域衍生一处盐度为31~31.5的低盐区(图2f)。夏季受沿岸上升流影响(吕新刚等, 2007), 长江口南部有一处低温区, 温度在23 °C左右, 调查区域南部受黑潮(和台湾暖流)影响, 温度超过28 °C且向北延伸(图2c); 受指向济州岛方向的长江冲淡水影响, 调查区域北部出现一处低盐区, 盐度为24左右(图2g)。秋季在长江口门外有一低温区, 温度约为20 °C, 调查区域南部温度超过24 °C (图2d); 长江冲淡水相较于夏季势力减弱, 仅在长江口南部有一处盐度低于28的低盐区, 黑潮表层水势力增强, 使盐度为34的等盐线向北扩张(图2h)。
东海四季表层DIN和DSi分布与盐度分布刚好相反, DIN和DSi在低盐的长江口浓度较高、在盐度较高的黑潮控制区浓度较低。其中DIN浓度春(10.71 μmol/L)、夏季(9.41 μmol/L)高于冬(6.20 μmol/L)、秋季(5.41 μmol/L); DSi浓度春季最高(16.64 μmol/L), 冬(9.91 μmol/L)、夏季(9.28 μmol/L)次之, 秋季最低(7.69 μmol/L)。DIP分布与DIN和DSi有较大差异: 冬季DIP高值区分布在东海中部; 春季高值区分布与冬季相似, 但范围明显缩小; 夏季受长江冲淡水和上升流影响, 高值区分布在长江口和舟山群岛附近; 秋季高值区除位于长江口门外, 在东海东南部也有一处高值(图2i~2l)。DIP浓度以春、秋季最高(均为0.44 μmol/L), 冬季次之(0.43 μmol/L), 夏季最低(0.27 μmol/L)。

图2 东海冬(a, e, i)、春(b, f, j)、夏(c, g, k)、秋季(d, h, l)表层温度(°C)、盐度和无机磷(DIP, μmol/L)分布
2.2 浮游植物群落组成与丰度分布
2011年四季调查共检出浮游植物9门509种(含变种、变型和未定种), 其中硅藻305种, 占浮游植物总种类数的59.9%; 甲藻154种, 占浮游植物总种类数的30.3%; 其余门类占9.8%。从种类数组成看, 不同季节均以硅藻和甲藻为主; 从细胞丰度组成看, 硅藻均占据绝对优势(>62.6%), 但蓝藻在秋、冬季占比较高, 分别为8.7%和35.7%。
东海浮游植物丰度分布季节变化明显。冬季浮游植物细胞丰度为(38.50~5 217.69)×103cells/m3, 平均值为472.36×103cells/m3, 冬季丰度高值区处于东海南部(27.5°N, 124.5°E), 最高值出现在E06站位。春季浮游植物细胞丰度为(10.50~599 546.49)×103cells/m3, 平均值为19 180.76×103cells/m3, 春季丰度高值区处于舟山群岛南部(29°N, 122.5°E), 最高值出现在T07站位。夏季浮游植物细胞丰度为(4.80~793 558.08) ×103cells/m3, 平均值为28 911.28×103cells/m3, 夏季丰度高值区位于长江口门外(31.5°N, 123.5°E), 最高值出现在F02站位。秋季浮游植物细胞丰度为(32.50~ 660 854.29)×103cells/m3, 平均值为30 496.91×103cells/m3, 秋季丰度高值区与夏季类似, 位于长江口门外(31.5°N, 123.5°E), 最高值出现在F03站位。四季浮游植物细胞丰度大小为秋>夏>春>冬。在空间分布上, 冬季浮游植物细胞丰度表现为由沿岸至外海递增的趋势, 春、夏、秋季表现为由沿岸至外海递减的趋势(图3a~3d)。硅藻细胞丰度时空分布与浮游植物总丰度分布类似。冬、夏、春季的甲藻细胞丰度分布与浮游植物总丰度分布类似, 春季甲藻丰度高值区位于东海南部, 四季甲藻细胞丰度大小为夏>秋>春>冬(图3i~3l)。
2.3 浮游植物优势种
东海网采浮游植物优势种以硅藻为主, 甲藻仅有三角角藻() 1种, 蓝藻有铁氏束毛藻()和红海束毛藻() 2种。各季节优势种组成有明显差异(表2), 主要优势种细胞丰度分布与浮游植物总细胞丰度分布相似(图4)。冬季主要优势种为铁氏束毛藻, 细胞丰度高值区分布在黑潮区, 次高值区分布在长江口门外和东海南部, 占浮游植物总丰度的31.3%; 骨条藻(spp.)是春、夏季的绝对优势种, 细胞丰度高值区分别在舟山群岛东南部和长江口门外, 分别占浮游植物总丰度的89.3%和80.6%; 秋季主要优势种为柔弱伪菱形藻(), 细胞丰度高值区分布在长江口门外, 占浮游植物总丰度的48.7%。

表1 东海四季浮游植物种类数和丰度

图3 东海冬(a, e, i)、春(b, f, j)、夏(c, g, k)、秋季(d, h, l)浮游植物总丰度、硅藻丰度和甲藻丰度分布

表2 东海四季优势种细胞丰度(×103 cells/m3)与占比(%)

续表
注: —表示该种不是优势种。

图4 东海冬(a)、春(b)、夏(c)、秋季(d)浮游植物主要优势种细胞丰度(×106 cells/m3)分布
2.4 浮游植物种类数
浮游植物种类数的季节差异显著(<0.05)。冬季种类数为45~128, 平均值为74.6, 最高值出现在Z07站位。春季种类数为21~78, 平均值为43.9, 最高值出现在E03站位。夏季种类数为14~112, 平均值为48.1, 最高值出现在P12。秋季种类数为54~117, 平均值为82.4, 最高值出现在Z01站位。种类数秋>冬>夏>春, 水平分布为由近岸向外海递增, 黑潮影响海域种类数最多(图5)。
2.5 浮游植物与环境因子的关系
ANOSIM分析表明, 2011年东海浮游植物群落组成季节差异显著(=0.001,=0.55)。MDS分析表明, 四季浮游植物群落可大致分为三个生态类群为主(图6)。其中, 组1以近岸低盐种(如骨条藻和尖刺伪菱形藻等)占主导, 主要受长江冲淡水和沿岸流影响; 组2以广温广盐种(如具槽帕拉藻和三角角藻等)占主导, 主要受陆架混合水影响; 组3以外海高温高盐种(如铁氏束毛藻)占主导, 主要受黑潮影响。
Spearman相关性显示: 冬、春、秋季浮游植物总丰度与环境因子相关性均较弱, 夏季浮游植物总丰度与DIN和DIP呈正相关, 与盐度呈负相关, 表明长江冲淡水和黑潮次表层水的入侵对东海夏季浮游植物细胞丰度有重要影响。四季浮游植物种类数大多与温度、盐度呈正相关, 与营养盐呈负相关, 表明东海浮游植物种类组成受高温、高盐、寡营养的黑潮水控制。

图5 东海冬(a)、春(b)、夏(c)、秋季(d)种类数(S)分布

图6 基于聚类和多维尺度分析的东海四季浮游植物群落空间分布(1代表沿岸低盐种, 2代表广温广盐种, 3代表高温高盐种)
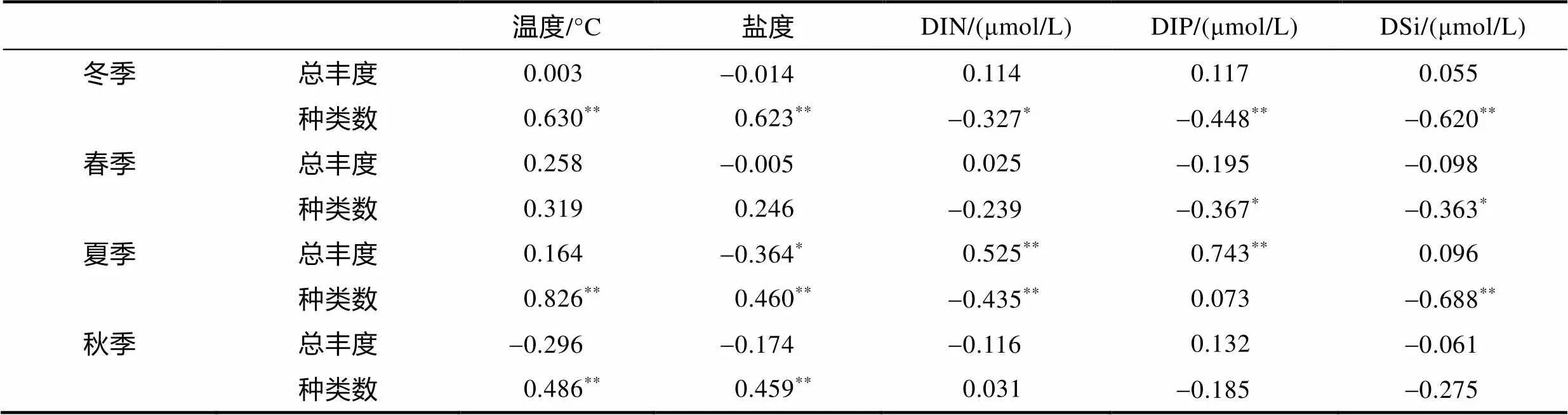
表3 浮游植物细胞丰度和种类数与环境因子的相关系数
注: DIN: 溶解无机氮; DIP: 溶解无机磷; DSi: 溶解硅酸盐;*:<0.05;**:<0.01
蒙特卡罗检验显示, 四季CCA分析中的第一轴和全部轴均为显著(<0.01), 因此排序结果可信。其中, 冬季洛氏角毛藻()和柔弱伪菱形藻等暖水种与温度、盐度呈密切正相关, 而与营养盐负相关, 表明这些物种易于高温、高盐、寡营养的黑潮水和台湾暖流水中占优势; 束毛藻与营养盐呈正相关, 而与温度关系不大, 表明冬季温度和营养盐并非其限制因素。春季骨条藻处于中心位置, 表明其对环境的适应性较好; 柔弱伪菱形藻、洛氏角毛藻和短叉角毛藻()等与温度密切相关, 表明其适宜生长在受黑潮和台湾暖流控制的暖水环境。夏季骨条藻与营养盐密切相关, 与温度和盐度呈负相关, 表明其适宜生长于低盐、富营养的长江冲淡水和沿岸水; 铁氏束毛藻与温度、盐度密切相关, 表明其在高温高盐的黑潮水中占优势。秋季柔弱伪菱形藻与营养盐密切相关, 与温度负相关, 表明其适宜生长在长江冲淡水锋面附近相对低温的环境; 铁氏束毛藻和热带骨条藻()等与温度密切相关, 与营养盐负相关, 表明其适宜生长在相对寡营养的暖水环境中。
图8显示, 冬、秋季长江冲淡水势力较弱、水体混合剧烈, 浮游植物大都分布在N/P=16附近; 春、夏季长江冲淡水势力较强、水体分层现象加剧, 调查区域N/P大都高于16, 磷限制较明显, 且夏季浮游植物丰度高值区N/P高于春季。
3 讨论
3.1 黑潮对浮游植物细胞丰度分布的影响
作为太平洋的西边界流, 黑潮阻隔了东海与太平洋直接进行水体交换, 但黑潮入侵能将大洋的物质和能量输送到东海, 使黑潮表层水和次表层水的季节变化与东海浮游植物细胞丰度分布有密切关系(杨德周等, 2017)。表3和图7显示, 不同季节浮游植物丰度与水团消长及其相关的温度、盐度和营养盐有密切关系。冬季东北季风势力增强, 东海表层海温下降, 且光照减弱, 沿岸浮游植物的生长受到温度、光限制, 导致浮游植物细胞丰度较低, 但黑潮表层水入侵东海南部, 水体透明度较高, 并为浮游植物提供适宜的生长温度和盐度, 适宜铁氏束毛藻等高温高盐种生长, 从而形成浮游植物细胞丰度高值区(图3a)。郭术津等(2012)研究表明, 2009年冬季黑潮占据的东海海域以铁氏束毛藻等为主要优势种。春季黑潮表层水入侵仅限于东海陆架外围, 34等盐线南移, 且东北季风减弱, 水体趋于稳定。沿岸流携带了丰富的营养盐, 但盐度较低, 仅利于骨条藻等沿岸低盐种生长, 导致骨条藻细胞丰度占比高达89.3% (表2)。Yang等(2018)指出黑潮次表层水入侵的近岸分支在春季开始出现并入侵至浙江近岸50 m等深线处。但入侵势力相对较弱(赵瑞祥等, 2014), 难以上升到表层。春季由于黑潮表层水势力减弱, 东海浮游植物细胞丰度高值区主要位于沿岸水和陆架混合水区域。夏季西南季风盛行, 黑潮次表层水与台湾暖流势力增强, 向北入侵加剧, 与长江冲淡水交汇形成的锋面处透明度较高、营养盐丰富, 使得锋面处(盐度较低一侧)有较高的浮游植物细胞丰度(图3c)。这与以往研究一致(Gomes, 2018; Xu, 2019)。秋季长江冲淡水势力减弱, 但黑潮表层水入侵增强, 33.5等盐线北移, 导致骨条藻等沿岸低盐种的细胞丰度占比降低, 而柔弱伪菱形藻等暖水种细胞丰度占比上升(表2); 图2和图3显示秋季浮游植物丰度高值位于黑潮和台湾暖流控制海域。这与以往的调查资料相符(Tang, 2006; Guo, 2014)。罗民波等(2007)指出, 环流是东海浮游植物数量变化和平面分布特征形成的重要因素。

图7 东海四季浮游植物优势种与环境因子间的典范对应分析

图8 东海冬(a)、春(b)、夏(c)、秋季(d)浮游植物丰度在DIN和DIP中的分布
除温度、盐度外, 营养盐等环境因子对浮游植物群落组成和分布也具有重要的调控作用(Jiang, 2015; Wei, 2017)。宋金明等(2019)指出, 黑潮跨陆架输运各种形态的生源要素到达东海, 影响东海碳和营养盐的收支, 使东海生态系统时空变化明显。2011年东海浮游植物总丰度呈现秋>夏>春>冬的趋势, 硅藻细胞丰度占比均最高, 这与Guo等(2014)调查结果类似。冬季黑潮表层水势力较强, 但次表层水势力处于全年最弱, 加之低温和光照等限制, 浮游植物丰度较低, 但黑潮表层水携带的束毛藻在群落中占比较高。春季黑潮表层水势力较弱, 次表层水势力开始增强, 水体混合减弱、水温逐渐升高, 加之冬季未被浮游植物消耗而遗留的丰富营养盐(DIN、DIP和DSi均最高), 浮游植物生长加快, 其总细胞丰度高于冬季, 尤其在近岸发生了硅藻赤潮, 使得硅藻细胞丰度较冬季明显增加。研究表明, 在低透明度、营养盐丰富的条件下, 硅藻因较高的DIN同化率而具有更高的生长速率(Smayda, 1997; Goldman, 2003)。这佐证了我们的观点。夏季, 长江冲淡水势力较强, 向东海输送营养盐的N/P接近100 (Harrison, 1990; Li, 2009), 磷限制较严重, 例如浮游植物优势种与DIP相关性很弱(图7c)。富含DIP的黑潮次表层水(N/P为8.7~16.3)入侵东海陆架后改善了夏季近海的磷限制(赵卫红等, 2006), 浮游植物在次表层水涌升处旺发形成细胞丰度高值区。秋季黑潮表层水势力开始增强而次表层水势力开始减弱, 但次表层水仍能到达28.5°N附近, 黑潮表层水和次表层水输送的DIP与近岸丰富的DIN和DSi为浮游植物生长提供了良好的营养环境, 使秋季浮游植物细胞丰度最高; 此外, 秋季DIN和DSi浓度最低, 极可能与硅藻旺发消耗DIN和DSi有关。张传松等(2008)发现在硅藻赤潮期间硅酸盐浓度呈直线下降的趋势。图7d显示秋季主要优势种链状硅藻柔弱伪菱形藻与营养盐有相关性但不明显, 表明秋季调查时浮游植物已消耗了大量硅酸盐。杨德周等(2017)对东海环流与生态耦合模式进行生态模拟, 结果显示黑潮入侵分支磷酸盐输送的模拟结果与叶绿素的分布结果一致。可见, 黑潮的入侵携带的DIP深刻影响东海浮游植物细胞丰度的分布。
3.2 黑潮对浮游植物群落组成影响
从已有的文献报道看, 东海浮游植物的调查多是单个季节(栾青杉等, 2007; 宋书群等, 2008; 王云龙等, 2008), 且针对浮游植物群落季节变化的研究多集中在长江口及其邻近海域(吴玉霖等, 2004; 何青等, 2009)。东海环流众多, 水文动力条件复杂。受长江冲淡水、黑潮、台湾暖流和黄海沿岸流等影响, 浮游植物物种组成丰富(表1), 主要生态类群有近岸低盐种、广温广盐种和外海高温高盐种等。MDS结果显示, 东海浮游植物群落具有明显的季节变化(图6), 且ANOSIM表明四季浮游植物群落具有显著(<0.05)的时空差异。冬季以蓝藻束毛藻占主导, 其余季节均以硅藻为主。受黑潮表层水和次表层水入侵季节变化影响, 东海浮游植物优势种季节更替现象明显。其中, 冬季优势种以外海高温高盐种铁氏束毛藻为主, 这与以往研究一致(罗民波等, 2007)。春季优势种以骨条藻、细弱海链藻硅藻为主, 这与宋书群等(2008)研究结果一致。夏季近岸低盐种骨条藻在东海占绝对优势, 这与以往研究一致(顾新根等, 1995; 林峰竹等, 2008; 王云龙等, 2008; 王俊等, 2009; Jiang等, 2014)。秋季优势种以柔弱伪菱形藻和铁氏束毛藻等暖水种为主, 这与刘海娇等(2015)调查结果一致。浮游植物的生态特征与其生存的环境具有良好的协调性(Lin, 2020)。四季浮游植物优势种中均有暖水种存在, MDS显示高温高盐种在四季空间分布较广, 且黑潮影响区内检出的高温高盐暖水种如束毛藻属、鸟尾藻属、梨甲藻属和很多角藻种类均可指示黑潮的入侵。黑潮影响区浮游植物暖水种数量显著(<0.05)高于长江口(附表1), 表明黑潮作为浮游植物暖水“种库”, 携带大量浮游植物暖水种到达东海, 对东海浮游植物群落组成产生重要影响。
受黑潮表层水和次表层水入侵影响, 东海四季浮游植物种类数高值区均在高温、高盐的南部或东南部(图5)。表3也证实, 冬、夏、秋季浮游植物种类数与温度和盐度呈显著(<0.01)正相关, 这与江志兵等(2014)研究结果一致。CCA排序较好地反映了浮游植物群落与环境因子间的关系(图7), 结合相关性分析(表3), 冬、春、秋季浮游植物总丰度与环境因子无明显相关性, 但多数优势种与温度、盐度表现为正相关。如冬、春季的洛氏角毛藻和柔弱伪菱形藻等, 秋季的铁氏束毛藻、热带骨条藻和优美旭氏藻矮小变型等, 推测黑潮表层水的入侵为上述优势种提供了适宜的温度和盐度。赵瑞祥等(2015)研究表明, 黑潮表层水在冬、春、秋季入侵东海陆架的程度分别为41.2%、25.0%和31.3%。夏季浮游植物总丰度与盐度呈负相关, 与DIN和DIP呈正相关, 推测浮于上层的高N/P的长江冲淡水和下层的低N/P的黑潮次表层水共同促进了浮游植物的生长, 使夏季浮游植物总细胞丰度与盐度和氮、磷营养盐浓度表现出明显的相关性。宋金明等(2017)指出, 黑潮次表层水在夏半年输送东海的DIN和DIP通量分别为85.1×109和5.79×109mol, 是黑潮表层水的5.16和3.27倍。本次调查对照区(长江口)磷酸盐浓度显著(<0.05)高于黑潮影响区, 原因是本研究仅有表层(3 m)营养盐数据, 黑潮表层水营养盐含量较低, 且黑潮次表层水在100~ 200 m等深线内至多涌升至30 m层(陆赛英等, 1996)。黑潮影响区与对照区N/P无显著差异, 但黑潮影响区N/P均值(20.6)低于对照区(28.3)。图8表明春、夏季节浮游植物丰度高值区N/P在50~80间, 存在明显的磷限制。黑潮次表层水携带大量磷酸盐入侵东海陆架, 缓解了磷限制, 导致嗜氮的骨条藻(洪君超等, 1993)成为春、夏季的绝对优势种(>80.6%)。可见, 黑潮提供磷对东海浮游植物群落组成和藻华影响较大。Jiang等(2014, 2015, 2019)研究指出, 黑潮、台湾暖流、长江冲淡水等环流变化和水团消长对长江口及邻近东海陆架浮游植物群落组成与分布有重要影响。周茜茜等(2015)指出中国东部陆架边缘海的浮游植物群落主要受低盐沿岸水系、黄海中部水系和黑潮暖流水系的影响。综上所述, 东海浮游植物的群落组成和时空分布与黑潮表层水和次表层水的季节、空间变化密切相关。
4 结论
2011年四季东海网采浮游植物群落均以硅藻为主, 但蓝藻丰度也较高。在水平分布上, 营养盐丰富的近岸以骨条藻等硅藻为主, 寡营养的外海以束毛藻等为主。受长江冲淡水和黑潮影响, 浮游植物细胞丰度高值区位于长江口门外、舟山群岛东南部和东海南部。优势种主要有骨条藻、柔弱伪菱形藻和铁氏束毛藻等。浮游植物群落组成季节、空间差异显著, 与黑潮表层水和次表层水的季节、空间变化有较好的耦合性。冬季以束毛藻为代表的外海高温高盐种为主, 春、夏季以嗜氮的骨条藻为代表的沿岸低盐种占优势,秋季以低盐暖水种为主, 且黑潮区暖水种数量显著高于长江口。黑潮携带的DIP等促进了东海浮游植物的生长, 同时也输入了大量暖水种, 深刻影响东海浮游植物的群落组成和分布。
致谢 感谢中国海洋大学江文胜教授提供温度和盐度数据以及东方红2号船在采样过程中的大力协助。
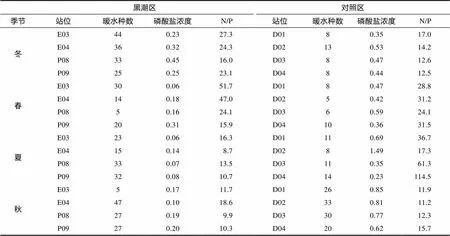
附表1 东海四季黑潮区与对照区暖水种数量、磷酸盐浓度(μmol/L)和氮磷比(N/P)
王云龙, 袁骐, 沈新强, 2008. 长江口及邻近海域夏季浮游植物分布现状与变化趋势[J]. 海洋环境科学, 27(2): 169-172.
王俊, 陈瑞盛, 左涛, 2009. 三峡工程截流后长江口邻近海域的网采浮游植物群落结构特征[J]. 水生态学杂志, 2(2): 80-87.
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008. 海洋调查规范第6部分: 海洋生物调查: GB/T 12763.6-2007[S]. 北京: 中国标准出版社.
吕新刚, 乔方利, 夏长水, 等, 2007. 长江口外及浙江沿岸夏季上升流的潮生机制[J]. 中国科学 D 辑: 地球科学, 37(1): 133-144.
刘东艳, 孙军, 钱树本, 2000. 琉球群岛及其邻近海域的浮游甲藻——1997年夏季的种类组成和丰度分布[C] // 中国海洋学文集——西北太平洋副热带环流研究(二). 北京: 海洋出版社.
刘海娇, 傅文诚, 孙军, 2015. 2009—2011年东海陆架海域网采浮游植物群落的季节变化[J]. 海洋学报, 37(10): 106-122.
江志兵, 曾江宁, 李宏亮, 等, 2014. 长江口及其邻近陆架区夏季网采浮游植物及其影响因素[J]. 海洋学报, 36(6): 112-123.
孙振皓, 盛留洋, 江新琴, 等, 2021. 2017年夏季长江口浮游植物暖水种增多现象分析[J]. 海洋学研究, 39(4): 82-90.
李俊龙, 郑丙辉, 刘录三, 等, 2013. 长江口浮游植物群落特征及其与环境的响应关系[J]. 环境科学研究, 26(4): 403-409.
杨世民, 董树刚, 2006. 中国海域常见浮游硅藻图谱[M]. 青岛:中国海洋大学出版社.
杨清良, 林更铭, 林金美, 1999. 1996年春季副热带环流区浮游植物生态的初步研究[J]. 海洋学报, 21(5): 120-128.
杨德周, 尹宝树, 侯一筠, 等, 2017. 黑潮入侵东海陆架途径及其影响研究进展[J]. 海洋与湖沼, 48(6): 1196-1207.
吴玉霖, 傅月娜, 张永山, 等, 2004. 长江口海域浮游植物分布及其与径流的关系[J]. 海洋与湖沼, 35(3): 246-251.
何青, 孙军, 2009. 长江口及其邻近水域网采浮游植物群落[J]. 生态学报, 29(7): 3928-3938.
宋书群, 孙军, 沈志良, 等, 2008. 长江口冬季和春季浮游植物的粒级生物量[J]. 应用生态学报, 19(3): 658-666.
宋金明, 王启栋, 张润, 等, 2019. 70年来中国化学海洋学研究的主要进展[J]. 海洋学报, 41(10): 65-80.
宋金明, 袁华茂, 2017. 黑潮与邻近东海生源要素的交换及其生态环境效应[J]. 海洋与湖沼, 48(6): 1169-1177.
张传松, 王江涛, 朱德弟, 等, 2008. 2005年春夏季东海赤潮过程中营养盐作用初探[J]. 海洋学报, 30(3): 153-159.
陆赛英, 葛人峰, 刘丽慧, 1996. 东海陆架水域营养盐的季节变化和物理输运的规律[J]. 海洋学报, 18(5): 41-51.
林更铭, 杨清良, 2007. 台湾海峡小型浮游植物的物种多样性和分布特征[J]. 生物多样性, 15(1): 31-45.
林峰竹, 吴玉霖, 于海成, 等, 2008. 2004年长江口浮游植物群落结构特征分析[J]. 海洋与湖沼, 39(4): 401-410.
罗民波, 陆健健, 王云龙, 等, 2007. 东海浮游植物数量分布与优势种[J]. 生态学报, 27(12): 5076-5085.
周茜茜, 陈长平, 梁君荣, 等, 2015. 中国东部陆架边缘海网采浮游植物种类组成和季节变化[J]. 生物多样性, 23(1): 23-32.
赵卫红, 王江涛, 李金涛, 等, 2006. 长江口及邻近海域冬夏季浮游植物营养限制及其比较[J]. 海洋学报, 28(3): 119-126.
赵瑞祥, 刘志亮, 2014. 台湾东北部黑潮次表层水入侵的季节变化规律[J]. 海洋学报, 36(1): 20-27.
赵瑞祥, 刘志亮, 2015. 台湾东北部黑潮表层水入侵东海陆架的季节变化规律[J]. 海洋科学, 39(7): 118-123.
洪君超, 黄秀清, 蒋晓山, 等, 1993. 嵊山水域中肋骨条藻赤潮发生过程主导因子分析[J]. 海洋学报, 15(6): 135-141.
顾新根, 袁骐, 杨蕉文, 等, 1995. 长江口羽状锋海区浮游植物的生态研究[J]. 中国水产科学, 2(1): 1-15.
栾青杉, 孙军, 宋书群, 等, 2007. 长江口夏季浮游植物群落与环境因子的典范对应分析[J]. 植物生态学报, 31(3): 445-450.
郭术津, 孙军, 戴民汉, 等, 2012. 2009年冬季东海浮游植物群集[J]. 生态学报, 32(10): 3266-3278.
Chen Y L L, Chen H Y, Gong G C,, 2004. Phytoplankton production during a summer coastal upwelling in the East China Sea [J]. Continental Shelf Research, 24(12): 1321-1338.
Clarke K R, Gorley R N, 2006. PRIMER v6: User Manual/Tutorial [M]. Plymouth, UK: PRIMER-E Ltd.
Goldman J C, McGillicuddy D J J, 2003. Effect of large marine diatoms growing at low light on episodic new production [J]. Limnology and Oceanography, 48(3): 1176-1182.
Gomes H D R, Xu Q, Ishizaka J,, 2018. The influence of riverine nutrients in Niche Partitioning of Phytoplankton communities–a contrast between the Amazon River Plume and the Changjiang (Yangtze) river diluted water of the East China Sea [J]. Frontiers in Marine Science, 5: 343.
Gong G C, Lee C Y L, Liu K K, 1996. Chemical hydrography and chlorophyll a distribution in the East China Sea in summer: implications in nutrient dynamics [J]. Continental Shelf Research, 16(12): 1561-1590.
Guo S J, Feng Y Y, Wang L,, 2014. Seasonal variation in the phytoplankton community of a continental-shelf sea: the East China Sea [J]. Marine Ecology Progress Series, 516: 103-126.
Harrison P J, Hu M H, Yang Y P,, 1990. Phosphate limitation in estuarine and coastal waters of China [J]. Journal of Experimental Marine Biology and Ecology, 140(1/2): 79-87.
Jiang Z B, Chen J F, Feng Z,, 2015. Controlling factors of summer phytoplankton community in the Changjiang (Yangtze River) Estuary and adjacent East China Sea shelf [J]. Continental Shelf Research, 101: 71-84.
Jiang Z B, Chen J F, Zhai H C,, 2019. Kuroshio shape composition and distribution of Filamentous Diazotrophs in the East China Sea and Southern Yellow Sea [J]. Journal of Geophysical Research: Oceans, 124(11): 7421-7436.
Jiang Z B, Liu J J, Chen J F,, 2014. Responses of summer phytoplankton community to drastic environmental changes in the Changjiang (Yangtze River) estuary during the past 50 years [J]. Water Research, 54: 1-11.
Li J, Glibert P M, Zhou M J,, 2009. Relationships between nitrogen and phosphorus forms and ratios and the development of dinoflagellate blooms in the East China Sea [J]. Marine Ecology Progress Series, 383: 11-26.
Lin G M, Chen Y H, Huang J,, 2020. Regional disparities of phytoplankton in relation to different water masses in the Northwest Pacific Ocean during the spring and summer of 2017 [J]. Acta Oceanologica Sinica, 39(6): 107-118.
Liu Z Q, Gan J P, Hu J Y,, 2021. Progress of studies on circulation dynamics in the East China Sea: the Kuroshio exchanges with the shelf currents [J]. Frontiers in Marine Science, 8: 620910, doi: 10.3389/fmars.2021.620910.
Qu T D, Tozuka T, Kida S,, 2016. Western Pacific and marginal sea processes [J]. World Scientific Series on Asia-Pacific Weather and Climate, 36(7): 151-186.
Smayda T J, 1997. Harmful algal blooms: their ecophysiology and general relevance to phytoplankton blooms in the sea [J]. Limnology and Oceanography, 42(5part2): 1137-1153.
Tang D L, Di B P, Wei G F,, 2006. Spatial, seasonal and species variations of harmful algal blooms in the South Yellow Sea and East China Sea [J]. Hydrobiologia, 568(1): 245-253.
Tseng Y F, Lin J, Dai M,, 2014. Joint effect of freshwater plume and coastal upwelling on phytoplankton growth off the Changjiang River [J]. Biogeosciences, 11(2): 409-423.
Tsui I F, Wu C R, 2012. Variability analysis of Kuroshio intrusion through Luzon Strait using growing hierarchical self-organizing map [J]. Ocean Dynamics, 62(8): 1187-1194.
Wang W T, Yu Z M, Song X X,, 2018. Intrusion Pattern of the Offshore Kuroshio branch current and its effects on nutrient contributions in the East China Sea [J]. Journal of Geophysical Research: Oceans, 123(3): 2116-2128.
Wei Y Q, Liu H J, Zhang X D,, 2017. Physicochemical conditions in affecting the distribution of spring phytoplankton community [J]. Chinese Journal of Oceanology and Limnology, 35(6): 1342-1361.
Wu C R, Hsin Y C, Chiang T L,, 2014. Seasonal and interannual changes of the Kuroshio intrusion onto the East China Sea Shelf [J]. Journal of Geophysical Research: Oceans, 119(8): 5039-5051.
Xu Q, Sukigara C, Goes J I,, 2019. Interannual changes in summer phytoplankton community composition in relation to water mass variability in the East China Sea [J]. Journal of Oceanography, 75(1): 61-79.
Yang D Z, Yin B S, Chai F,, 2018. The onshore intrusion of Kuroshio subsurface water from February to July and a mechanism for the intrusion variation [J]. Progress in Oceanography, 167: 97-115.
Yang D Z, Yin B S, Liu Z L,, 2012. Numerical study on the pattern and origins of Kuroshio branches in the bottom water of southern East China Sea in summer [J]. Journal of Geophysical Research: Oceans, 117(C2): C02014, doi: 10.1029/2011JC007528.
Zhang J, Liu S M, Ren J L,, 2007. Nutrient gradients from the eutrophic Changjiang (Yangtze River) Estuary to the oligotrophic Kuroshio waters and re-evaluation of budgets for the East China Sea Shelf [J]. Progress in Oceanography, 74(4): 449-478.
INFLUENCE OF KUROSHIO INTRUSION ON COMPOSITION AND DISTRIBUTION OF NET-COLLECTED PHYTOPLANKTON COMMUNITY IN THE EAST CHINA SEA
SUN Zhen-Hao1, 2, 4, JIANG Zhi-Bing1, 2, 3, 4, ZHAN Lin1, ZHU Yuan-Li1, 2, 4, ZENG Jiang-Ning1, 2, 4, CHEN Quan-Zhen1, CHEN Jian-Fang1
(1. Key Laboratory of Marine Ecosystem Dynamic, Second Institute of Oceanography, MNR, Hangzhou 310012, China; 2. Key Laboratory of Nearshore Engineering Environment and Ecological Security of Zhejiang Province, Hangzhou 310012, China; 3. Key Laboratory of Ocean Space Resource Management Technology, Ministry of Natural Resource, Marine Academy of Zhejiang Province, Hangzhou 310012, China; 4. Observation and research station of Yangtze River Delta Marine Ecosystems, Marine of Natural Resources, Zhoushan 316021, China)
The intrusion of Kuroshio influences profoundly the ecology and environment of the East China Sea (ECS). However, its influence on phytoplankton community composition and distribution remain unclear. To analyze the phytoplankton abundance and dominant species composition and their responses to the Kuroshio intrusion into the ECS, phytoplankton samples were collected vertically using net trawl method from 164 stations (26°~33°N, 121°~128°E) in the four seasons. Among all the samples collected, 9 phyla and 509 species were identified, including 305 diatom, 154 dinoflagellate, and a few cyanobacteria, haptophyta, chrysophyte, euglenophyte, chlorophyte, cryptophyte, and xanthophyte species. The seasonal pattern of phytoplankton abundance showed an order of autumn (30 496.91×103cells/m3) > summer (28 911.28×103cells/m3) > spring (19 180.76×103cells/m3) > winter (472.36×103cells/m3). High phytoplankton abundance was observed in the front between Kuroshio and the Changjiang (Yangtze River) Diluted Water, the southeast of Zhoushan Islands, and the southeast part of the ECS. A high-temperature and high-salinity species,was dominant in winter under the intrusion of Kuroshio surface water.spp. was dominant in spring and summer. Phytoplankton abundance was higher in summer than in spring because the enhanced Changjiang Diluted Water and intrusion of Kuroshio subsurface water transported abundant nitrogen and phosphorus, respectively. In autumn, phytoplankton abundance and dissolved inorganic phosphorus concentration were the highest due to the influence of Kuroshio surface water and subsurface water; the community was mainly dominated by-. The species number showed an order of autumn > winter > summer > spring. A large number of warm-water phytoplankton species was found in Kuroshio water, resulting in increased species number from coastal to offshore waters. Similarity analysis and multidimensional scaling showed that phytoplankton community composition varied significantly with seasonal and spatial scale. These results demonstrate that the abundant phosphorus and offshore warm-water species transported by Kuroshio promote the growth of phytoplankton and enhance the species diversity, which influence profoundly the seasonal and spatial patterns of phytoplankton community composition in the ECS.
East China Sea; Kuroshio; phytoplankton; community composition; seasonal variation
Q948
10.11693/hyhz20220500142
*浙江省尖兵研发攻关计划, 2022C03044号; 国家重点研发计划课题, 2021YFC3101702号; 国家自然科学基金, 41876198号; 浙江省自然科学基金, LR22D060001号; 长江口-浙江近海-邻近东海多学科长期观测(LORCE)计划, SZ2001号。孙振皓, 硕士研究生, E-mail: sunzhenhao98@163.com
江志兵, 博士生导师, 研究员, E-mail: jzb@sio.org.cn
2022-05-27,
2022-07-16
猜你喜欢
海洋通报(2022年6期)2023-01-07
潍坊学院学报(2020年2期)2021-01-18
流行色(2019年10期)2019-12-06
海洋世界(2017年1期)2017-02-13
环境科技(2016年2期)2016-11-08
海洋科学(2016年1期)2016-07-06
海洋渔业(2016年6期)2016-04-16
海洋渔业(2016年6期)2016-04-16
应用海洋学学报(2015年3期)2015-11-22
华东师范大学学报(自然科学版)(2014年4期)2014-03-11
