
虾青素对猪卵母细胞体外成熟的影响
2023-01-17 09:01:18许建春汪浩鑫自永宏杨小芬陈梦佳石德顺陆凤花
中国兽医学报 2022年12期
许建春,汪浩鑫,自永宏,杨小芬,闫 茜,陈梦佳,石德顺,陆凤花
(广西大学 亚热带农业生物资源保护与利用国家重点实验室,广西 南宁530004)
卵母细胞体外成熟是进行体外胚胎生产的关键环节,对胚胎生物技术的发展起着重要的作用。目前,卵母细胞体外成熟技术存在成熟率低和发育能力差等诸多问题,极大地阻碍了该技术的广泛应用。与体内成熟的卵母细胞相比,体外成熟的卵母细胞往往会出现发育阻滞现象。这可能是因为在体外培养的卵母细胞对外界环境变化敏感,且离体培养的环境与内部环境之间存在一定差异,比如体外环境产生的活性氧、培养液成分、O2浓度、光照、温度和pH值等都会影响卵母细胞正常发育[1-4]。卵母细胞的质量与胚胎发育潜能存在密切联系,高质量的卵母细胞有利于正常受精及胚胎后续发育[5]。在众多影响卵母细胞体外成熟质量的因素中,导致卵母细胞体外发育能力改变的一个重要因素就是活性氧(ROS)的产生。活性氧作为主要的促氧化剂,可以导致DNA损伤、线粒体功能障碍、脂质过氧化等,从而使卵母细胞质量下降,阻碍其后续发育。因此,如何提高卵母细胞成熟质量显得格外重要。虾青素是一种红色的类胡萝卜素,对多种细胞系具有抗氧化和抗炎作用,可以抑制氧化应激诱导的线粒体功能障碍和一些相关疾病的发生。虾青素可以保护线粒体免受内源性氧自由基的侵害,保持其氧化还原能力,从而提高其能量产生效率。虾青素在牛卵母细胞体外成熟中和其他细胞系上的研究有见报道。然而,虾青素对猪卵母细胞体外成熟效果的影响报道较少。本研究主要通过在猪卵母细胞体外成熟培养基中添加一定浓度的虾青素,以探讨其对猪卵母细胞成熟及早期胚胎发育的影响,以期提高卵母细胞的体外成熟效率和囊胚的质量,为猪卵母细胞和胚胎体外培养体系的完善提供依据。
1 材料与方法
1.1 主要试剂及液体配制组织培养液(TCM199)和胎牛血清(FBS)购自美国Gibco公司;虾青素购自成仪睿有限公司;活性氧检测试剂盒(S0033)和总谷胱甘肽检测试剂盒(S0052)购自碧云天生物技术公司;丙二醛检测试剂盒(WLA048)购自万类生物;卵母细胞洗液为CCM液(TCM199+5 mmol/L NaHCO3+5 mmol/L Hepes+3%发情牛血清(OCS));胚胎培养液(4.75 g TCM-199+13.1 mmol/L NaHCO3+30 mg/L 青霉素+50 mg/L 链霉素+2.5 mmol/L Hepes+12.5 mL FBS),搅拌均匀后调pH值为7.2~7.4;猪卵母细胞成熟液为PM液;胚胎培养液为NCSU-23液参照文献[6]配制;离子霉素溶液(胚胎培养液+5 μmol/L 离子霉素)和6-DMAP溶液(胚胎培养液+2 mmol/L 6-DMAP)等均添加60 mg/L青霉素和100 mg/L链霉素,并用0.22 μm微孔滤膜过滤器除菌。微量反转录试剂盒和定量试剂购自ABclonal公司。
1.2 猪卵母细胞的收集猪卵巢取自南宁市屠宰厂,将其置于盛有25~30℃生理盐水的保温瓶中,确保4 h内送达实验室。去除卵巢周围的脂肪组织后,用注射器抽取直径2~6 mm卵泡的卵泡液,将卵泡液置于60 mm玻璃皿中,在体视显微镜下挑出卵丘卵母细胞复合体(COCs),用卵母细胞清洗液清洗干净,只选取卵丘细胞层完整和胞质均匀的卵母细胞进行后续试验。
1.3 卵母细胞的成熟培养挑选含有3层及以上卵丘细胞包被的折光性良好的COCs,采用微滴培养体系(含10 IU/mL hCG+10 IU/mL PMSG)并且含有不同浓度虾青素(0,5,10,20,40 μmol/L)的PM液培养。20~22 h后换成不含激素的培养液,继续培养20~22 h至细胞成熟,统计第一极体排出率。
1.4 孤雌胚胎的激活与培养孤雌胚胎采用化学法激活,成熟的卵母细胞经0.1%透明质酸酶处理,去除周围包裹的卵丘细胞。挑选排出第一极体和胞质均匀的卵母细胞,将裸卵置于5 μmol/L的离子霉素溶液中培养5 min,转移卵母细胞至2 mmol/L 6-DMAP 溶液中继续培养3.5 h,然后将卵母细胞转移至胚胎培养盘中微滴培养,48 h后统计卵裂率,6~8 d统计囊胚率。
1.5 活性氧染色、总谷胱甘肽含量和丙二醛含量测定根据统计的第一极体排出率、卵裂率和囊胚率,选出最适处理浓度进行以下试验。成熟培养后的COCs去除卵丘细胞后,PBS清洗3遍。一部分细胞转移到含10 μmol/L DCFH-DA的PBS中,37℃避光孵育30 min。孵育后将细胞转移至CCM液滴中,荧光显微镜下拍照。谷胱甘肽含量的测定参考文献[7]操作方法进行。丙二醛含量的测定参考试剂盒说明书进行操作。
1.6 囊胚Hoechst33342染色收集第6~7天的囊胚,在不含Ca2+、Mg2+的PBS中清洗2~4遍,4%多聚甲醛(PFA)固定1 h,然后将囊胚转移至10 g/L Hoechst33342染色工作液中,室温条件下避光孵育10~20 min,孵育结束后用不含Ca2+、Mg2+的PBS清洗2~4遍,将染料清洗干净。取载玻片,并在中央滴加约1 μL的抗猝灭剂,放入1枚囊胚,在盖玻片四周涂上1圈凡士林,压片,荧光显微镜下观察染色情况,并统计细胞数。
1.7 实时荧光定量PCR收集对照组和处理组不同时期的早期胚胎,将其分别移入装有细胞裂解液的EP管中,于-80℃冰箱保存。微量反转录试验按照试剂盒说明书进行操作,以反转录得到的cDNA为模板,采用SYBR Green PCR试剂盒进行qRT-PCR检测,反应体系:2×SYBR GreenⅠMaster 10 μL,上、下游引物(10 μmol/L)各1 μL,cDNA 1 μL,ddH2O补至20 μL。PCR反应条件:95℃10 min;95℃15 s,60℃60 s,72℃30 s,共40个循环。引物是根据GenBank公布的猪胚胎抗氧化酶相关基因GPX4和SOD1的mRNA序列设计,引物信息见表1。最终使用2-△△Ct法计算目的基因的相对表达量。
1.8 统计分析用Image J软件对细胞活性氧和Hoechst染色的结果进行量化统计,采用SPASS 20.0软件对数据进行差异显著性分析,P<0.05表示差异显著,所有试验至少重复5次。
2 结果
2.1 不同浓度虾青素对卵母细胞体外成熟的影响卵母细胞在添加不同浓度(0,5,10,20,40 μmol/L)虾青素的成熟培养液中培养40~44 h后发现,与对照组相比,虾青素处理组的第一极体排出率均有显著提高(P<0.05),其中10 μmol/L处理组的第一极体排出率显著高于其他浓度处理组(P<0.05)(表2)。
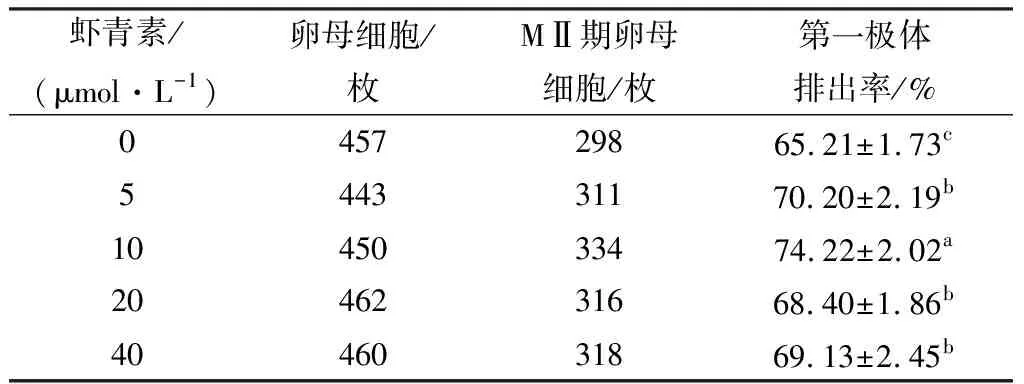
表2 虾青素对猪卵母细胞第一极体排出率的影响
2.2 成熟培养液中添加不同浓度虾青素对孤雌胚胎发育效果的影响对不同浓度虾青素处理的MⅡ期卵母细胞进行孤雌激活处理,培养160~168 h后统计卵裂率和囊胚率(表3)。结果发现,与对照组(0 μmol/L)相比,5,10,20 μmol/L处理组的卵裂率显著升高(P<0.05),40 μmol/L处理组的卵裂率有提高但无显著性差异(P>0.05);10,20 μmol/L处理组的囊胚率显著高于对照组(P<0.05),5,40 μmol/L 处理组的囊胚率有升高,但差异不显著(P<0.05);其中10 μmol/L处理组的第一极体排出率和囊胚率显著高于其他处理组,综合考虑,我们选择10 μmol/L的浓度进行后续试验。
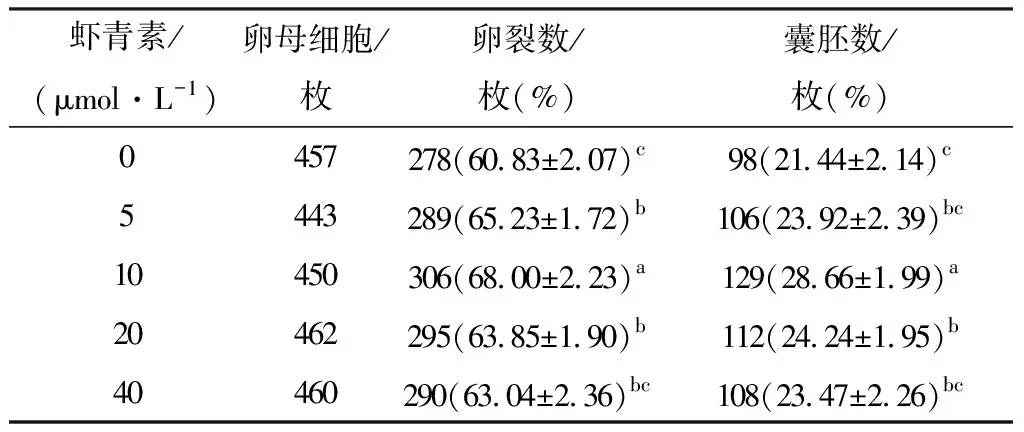
表3 虾青素对猪孤雌激活胚胎发育的影响
2.3 成熟液中添加10 μmol/L虾青素对MⅡ期卵母细胞抗氧化能力的影响为了解虾青素对卵母细胞抗氧化能力的影响,我们对MⅡ期的卵母细胞进行了活性氧染色、丙二醛和谷胱甘肽检测(图1)。结果发现,与对照组相比,10 μmol/L处理组卵母细胞的活性氧水平和丙二醛含量极显著降低(P<0.01),谷胱甘肽含量显著升高(P<0.05)。
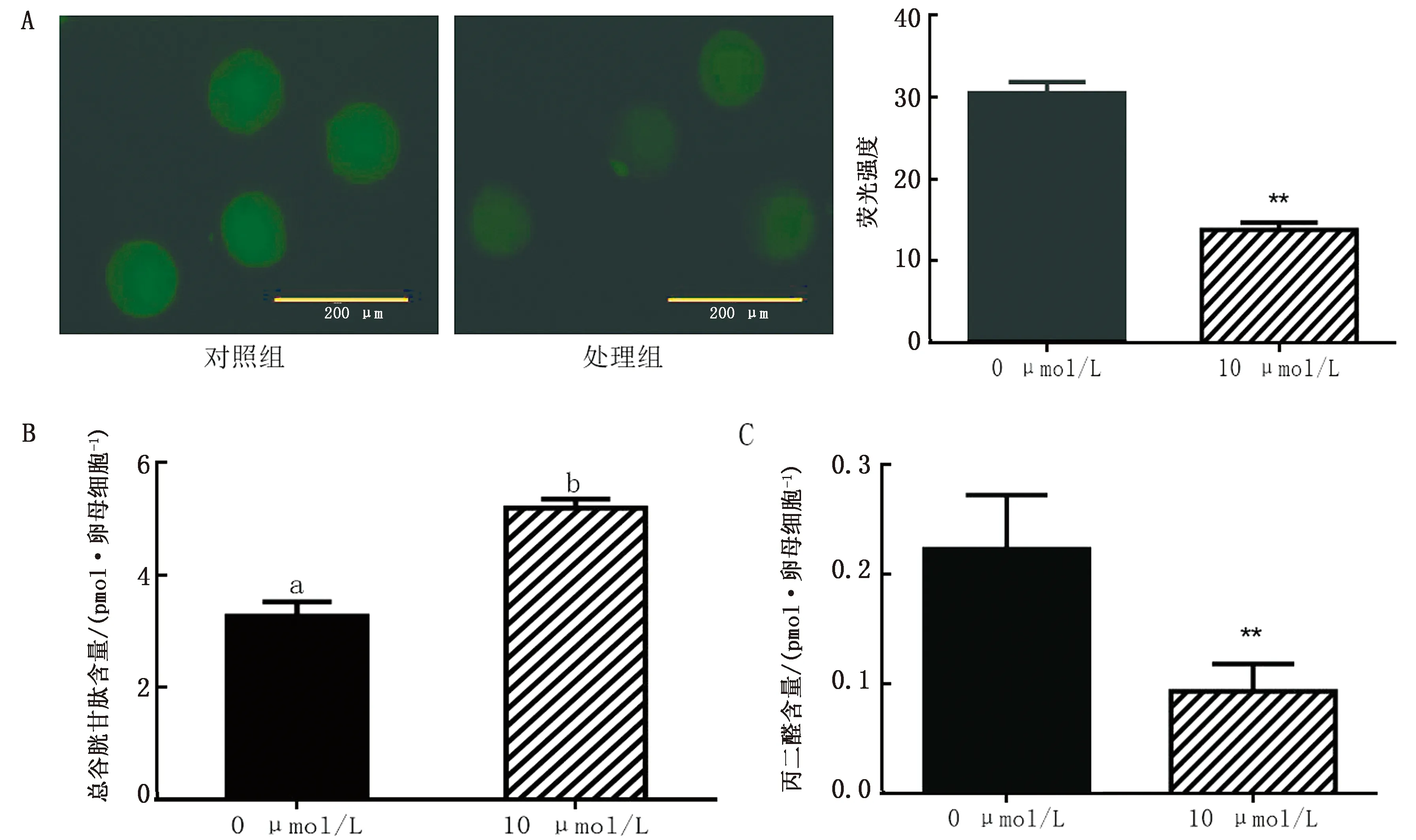
A.活性氧染色(标尺:200 μm);B.总谷胱甘肽含量检测;C.丙二醛含量检测。**表示差异极显著(P<0.01)。下同
2.4 成熟培养液中添加10 μmol/L虾青素对孤雌囊胚细胞数的影响使用10 μmol/L虾青素处理卵母细胞40~44 h后,对成熟的卵母细胞进行孤雌激活,使用Hochest33342染色法统计囊胚细胞数。结果如图2所示,10 μmol/L处理组的囊胚细胞数极显著高于对照组(P<0.01)。
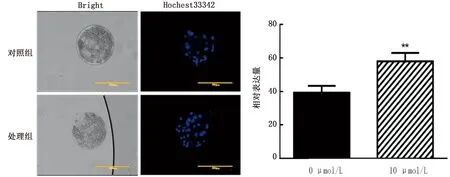
图2 虾青素处理对囊胚细胞数的影响(标尺:200 μm)
2.5 成熟培养液中添加10 μmol/L虾青素对胚胎发育过程中抗氧化酶相关基因表达的影响实时荧光定量PCR结果显示(图3),与对照组相比,GPX4基因在10 μmol/L处理组中MⅡ期卵母细胞中的表达显著升高(P<0.05),在囊胚期的表达也有升高。SOD1基因在10 μmol/L处理组中MⅡ期卵母细胞、4-细胞和囊胚期的表达极显著高于对照组(P<0.01)。
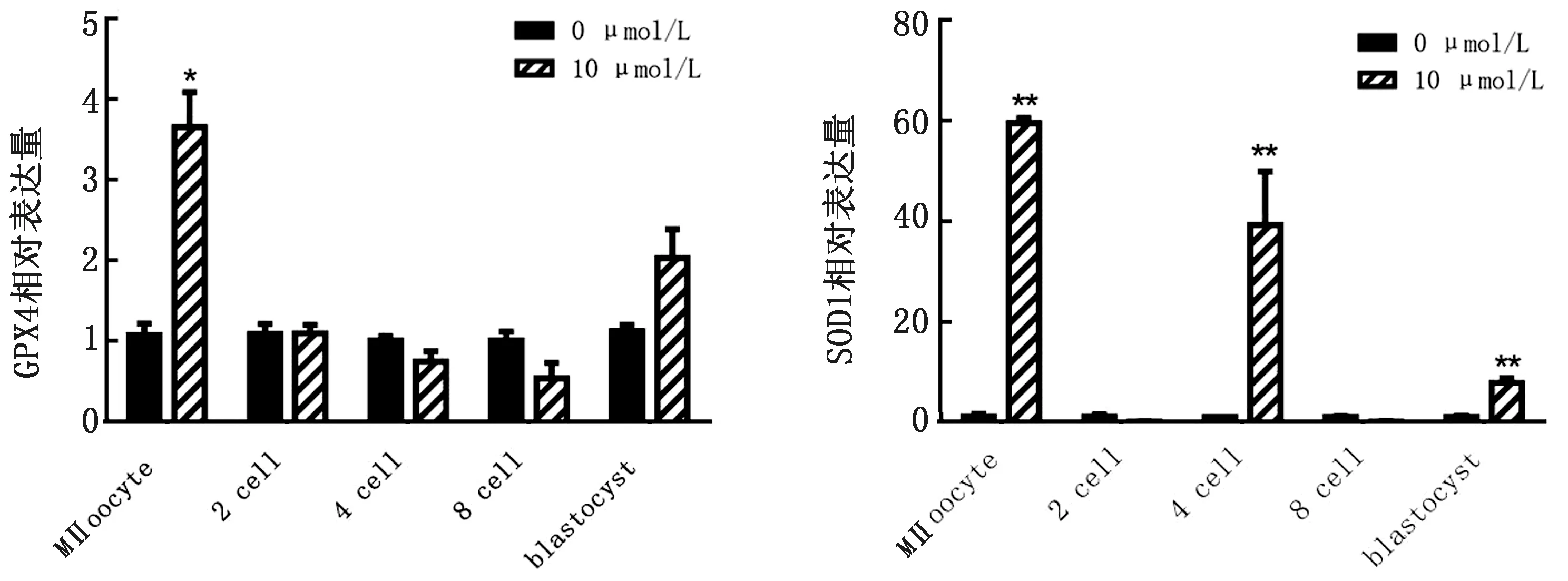
*表示差异显著(P<0.05)
3 讨论
体外培养系统的气相环境对卵母细胞成熟和早期胚胎发育起着关键的作用。氧气作为体外培养系统气相环境的因素之一,对胚胎新陈代谢和发育很重要。在体外20%氧气浓度远远高于体内的低氧环境,过高的氧气浓度会产生高浓度的活性氧和其他自由基,对卵母细胞造成氧化应激[8]。ROS是细胞内活性含氧化合物的总称,作为细胞内信号转导的重要信使,参与了多种增殖和分化等细胞活动,但当ROS产生超过细胞的抗氧化能力时则会造成氧化应激,氧化应激诱导线粒体损伤,可导致酶类失活、脂质过氧化反应、ATP消耗和线粒体功能紊乱。研究表明,氧化应激与卵母细胞质量下降、体外受精率降低和妊娠率降低有着密切联系[9-12]。针对这一问题,科研人员通过在培养基中添加酶类或非酶类的抗氧化剂,能够有效降低卵母细胞氧化应激损伤,提高卵母细胞的成熟质量和后续的胚胎发育[13-15]。虾青素是一种类胡萝卜素,主要来源于藻类和微生物,具有很强的抗氧化活性。此外虾青素还具有抗炎、抗肿瘤和神经保护作用[16-17]。然而,虾青素是否影响卵母细胞的体外成熟和胚胎发育却鲜有报道。本试验通过在成熟液中添加不同浓度的虾青素,处理44 h后发现其可以提高卵母细胞的第一极体排出率、分裂率和囊胚率,其中以10 μmol/L虾青素处理组效果显著,说明适当浓度的虾青素有利于猪卵母细胞的体外成熟和后续的胚胎发育。
研究发现,虾青素处理可降低组织中脂质氧化产物,提高超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GSH-Px)等抗氧化酶活性[18]。与其他的抗氧化剂相比,虾青素有更强的去除自由基能力,同时还可有效降低过氧化氢刺激引起的细胞凋亡[19-20]。本试验结果表明,10 μmol/L 虾青素可以极显著降低卵母细胞内的活性氧水平和丙二醛含量,显著提高总谷胱甘肽的含量,进一步说明了虾青素能够降低卵母细胞内活性氧水平,改善卵母细胞的成熟质量。
总之,在猪卵母细胞体外成熟液中添加适量的虾青素能有效降低卵母细胞内活性氧水平和丙二醛含量,提高总谷胱甘肽含量并促进抗氧化酶相关基因的表达,进而提高卵母细胞成熟的质量及其后续孤雌胚胎的发育能力。
猜你喜欢
当代水产(2021年6期)2021-08-13 08:17:10
落叶果树(2021年6期)2021-02-12 01:29:04
生殖医学杂志(2020年12期)2020-12-22 03:26:18
浙江医学(2020年17期)2020-09-21 09:06:30
河南畜牧兽医(2017年8期)2017-11-24 03:21:44
上海农业学报(2017年3期)2017-04-10 12:39:26
中国医学科学院学报(2015年5期)2015-03-01 04:03:36
郑州大学学报(医学版)(2015年2期)2015-02-27 14:50:51
生殖医学杂志(2013年5期)2013-03-11 18:43:47
食品科学(2013年17期)2013-03-11 18:27:10
