
贝莱斯芽孢杆菌(Bacillus velezensis)的研究进展
2023-01-16 05:00:02尉婧王碧香李诗瑶李二峰
天津农学院学报 2022年4期
尉婧,王碧香,李诗瑶,李二峰
(天津农学院 园艺园林学院,天津 300392)
芽孢杆菌(Bacillusspp.)分布范围广,对外界有害因子抵抗力强,属内绝大多数是有益微生物。它包含的部分种越来越多地被应用于抗虫杀菌剂、表面活性剂、生物制剂、增味剂和营养保健剂等方面[1]。贝莱斯芽孢杆菌是芽孢杆菌属的一个新种,最早于 1999年被分离到,但直到 2005年才被首次报道与命名[2]。近年来国内外对贝莱斯芽孢杆菌的报道越来越多,主要集中在贝莱斯芽孢杆菌的分类、应用及抗菌机理等方面。本研究就贝莱斯芽孢杆菌的发现与分类、基因组研究、生防作用及应用几个方面展开综述,以期为后续的研究与开发提供参考。
1 贝莱斯芽孢杆菌的发现与分类
贝莱斯芽孢杆菌最早被分离于西班牙马拉加省一条名为贝莱斯的河流[2],其分类地位的确定比枯草芽孢杆菌(Bacillus subtilis)、解淀粉芽孢杆菌(Bacillusamyloliquefaciens)、苏云金芽孢杆菌(Bacillus thuringiensis)等芽孢杆菌的报道都要晚[3]。由于贝莱斯芽孢杆菌与枯草芽孢杆菌、解淀粉芽孢杆菌、甲基营养型芽孢杆菌(Bacillus methylotrophicus)、暹罗芽孢杆菌(Bacillus siamensis)具有高表型相似性[4],因此只通过表型很难将其区分。通过对DNA亲缘值的分析,WANG等[5]认为贝莱斯芽孢杆菌是解淀粉芽孢杆菌的同种异名。在此基础上,DUNLAP等[6]通过对贝莱斯芽孢杆菌B-41580T菌株与解淀粉芽孢杆菌的基因组分析认为,贝莱斯芽孢杆菌不是解淀粉芽孢杆菌的同种异名,而是甲基营养型芽孢杆菌的同种异名。为了更好地分析这些相似菌株之间的系统遗传学,DUNLAP等[6]还将居水稻芽孢杆菌(Bacillus oryzicola,KACC18228)的全基因组序列与贝莱斯芽孢杆菌NRRL B-41580T、甲基营养型芽孢杆菌KACC 13015T和解淀粉芽孢杆菌植物亚种FZB42T的全基因组序列进行比对分析,结果显示它们的基因组之间仅有微小差异,且菌株间的 DNA杂交值均大于 84%,远远大于种间阈值70%。这些菌株具有相似的表型和基因型,因此提出居水稻芽孢杆菌、甲基营养型芽孢杆菌、解淀粉芽孢杆菌植物亚种是贝莱斯芽孢杆菌的同种异名。2018年,FAN等[7]通过使用TETRA、ANI、AAI和dDDH等技术对相关菌株进行分离鉴定,结果显示菌株FZB42T与贝莱斯芽孢杆菌代表菌株间的TETRA、ANT、AAI、ddH以及16S-rRNA值分别为0.999 1%、98.30%、99.11%、87.10%和99%,它们具有表型与基因型的高度一致性,远高于物种阈值,因此提出了“operational groupB.amyloliquefaciens”的新概念,并将Bacillus amyloliquefacienssubp.plantarumFZB42T更名为Bacillus velezensisFZB42T。除此之外,由于贝莱斯芽孢杆菌的发现早于其他菌株,因此将Bacillus oryzicola、Bacillus methylotrophicus、Bacillus amyloliquefacienssubp.plantarum重新归类并命名为Bacillus velezensis。近年来,有学者通过分析管家基因解旋酶亚基A(gyrA)、解旋酶亚基B(gyrB)构建的系统发育树并结合贝莱斯芽孢杆菌特异性引物PCR验证的方法,研究了其在贝莱斯芽孢杆菌及其近缘物种鉴定中的效果,为贝莱斯芽孢杆菌的快速筛选和分类鉴定奠定了基础[8]。
2 贝莱斯芽孢杆菌基因组的研究
近期,越来越多的研究人员对贝莱斯芽孢杆菌的不同亚种进行了全基因组测序。刘国强等[9]对贝莱斯芽孢杆菌 LS69菌株进行了全基因组测序,发现该菌株有一个约 3.92 Mbp的环形染色体,具有3 643个开放阅读框,确定了大环内酯(Macrolactin)、表面活性素(Surfactin)、泛革素(Fengycin)等10个化合物合成的基因簇,这些化合物均与贝莱斯芽孢杆菌的生防作用有关。此外,还发现贝莱斯芽孢杆菌LS69菌株有植物促生基因dhaS、ysnE和yhcX与促进植物免疫的基因tufA 、fliD、hag和flgK。PETROVA 等[10]对贝莱斯芽孢杆菌5R8菌株进行全基因组测序,发现该菌株含有一个3.91 Mbp的环形染色体,不含质粒,基因组中很大一部分基因用于碳水化合物的代谢,编码大多数参与多糖利用的酶,amyE、malL、sacA、xynA、xynB、xynD、xynC和eglS基因均编码糖苷水解酶。这种丰富的酶谱使纤维素、半纤维素、淀粉和菊粉的转化成为可能,并有望将贝莱斯芽孢杆菌应用于工业原料 2,3-丁二醇的生物生产中。CHEN等[11]通过全基因组测序,在贝莱斯芽孢杆菌157菌株的基因组中发现了编码与纤维素、木聚糖、木质素、淀粉、甘露聚糖、半乳糖苷和阿拉伯聚糖降解有关的酶的基因,表明贝莱斯芽孢杆菌有将农工业废弃材料(小麦秸秆、麦麸、甘蔗渣等)进行固态发酵的潜力。基因组研究使研究对贝莱斯芽孢杆菌的遗传、生物和生理特性等有了更深入的了解,有利于揭示其生物机制,为贝莱斯芽孢杆菌的分子机理研究和开发应用提供信息基础。
3 贝莱斯芽孢杆菌的生防作用
3.1 次生代谢产物及其相关基因
次生代谢产物是芽孢杆菌发挥生防作用的重要手段[12],近年来对贝莱斯芽孢杆菌生防作用的研究热点集中于次生代谢产物及其相关基因。贝莱斯芽孢杆菌产生的具有抗菌活性的次生代谢产物包括脂肽类化合物(Lipopeptides)、聚酮类化合物(Polyketides)、细菌素类(Bacteriocins)和抑菌蛋白(Peptides)等。脂肽类化合物是通过非核
糖体途径合成的,主要包括表面活性素(Surfactin)、伊枯草菌素(Iturin)、泛革素(Fengycin)、杆菌霉素(Bacillomycin)等;聚酮类化合物通过聚酮合酶合成途径合成,包含大环内酯(Macrolactin)、杆菌烯(Bacillaene)、地非西丁(Difficidin)等;细菌素类和抑菌蛋白类是通过核糖体途径合成的。其中细菌素类主要指的是羊毛硫抗生素(Lantibiotics)等,根据核糖体合成之后是否经过修饰可将细菌素分为ClassⅠ和ClassⅡ[13-14];抑菌蛋白包括蛋白酶、几丁质酶、β-葡聚糖酶。以上这些物质对多种病原菌具有良好的抑制作用,是潜在的传统抗生素的替代物。贝莱斯芽孢杆菌具有生防作用的次生代谢产物的合成是由多基因控制的,并且这些基因通常是成簇出现在基因组上,如表面活性素合成相关的基因簇是srfABCD,其中srfAB包含2个模块,srfC含有一个模块,该基因末端连接srfD;溶杆菌素合成相关基因是bacABCDEFGH,其中bacA、bacB、bacC、bacD、bacE和bacF分别编码不同的酶,而基因bacG编码催化蛋白。产生这些具有生防作用次生代谢物的相关基因(簇)如表1[3,15]所示。随着研究的不断深入,将会有更多的具有生防作用的次生代谢产物被发现,更多的合成基因被鉴定,进一步阐释抑菌物质的生物合成途径。
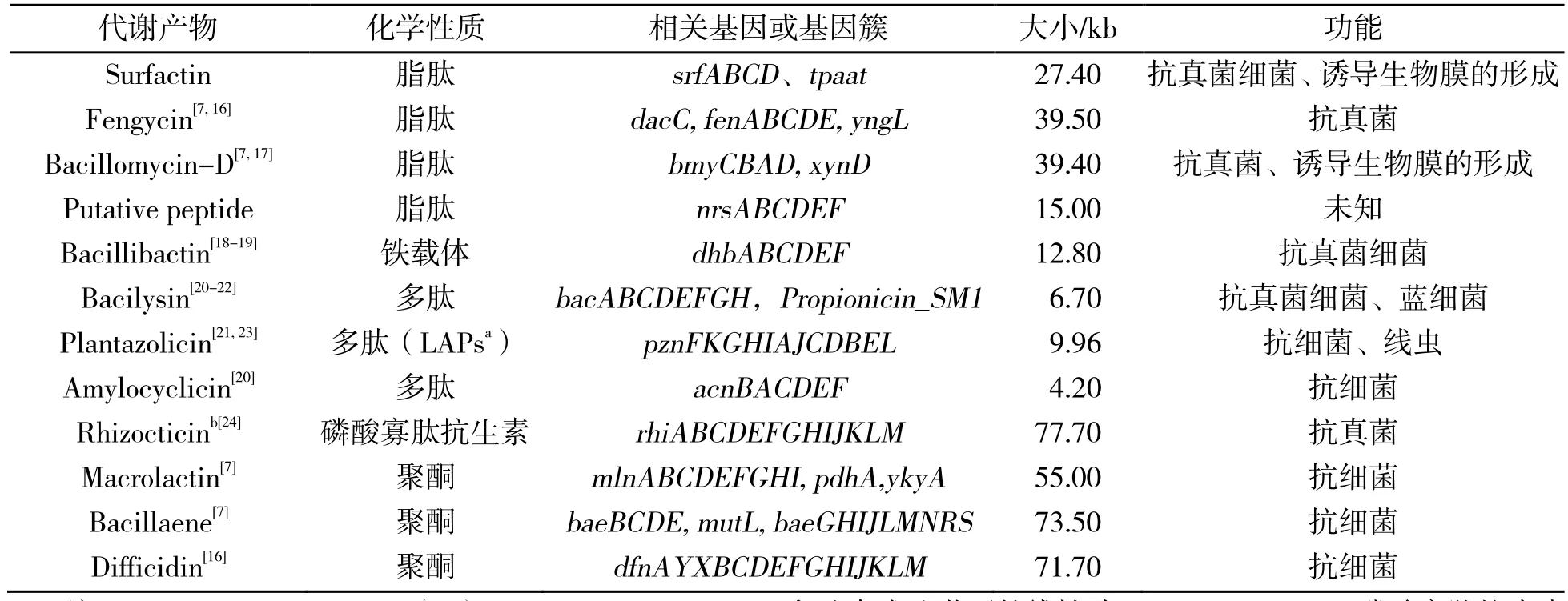
表1 贝莱斯芽孢杆菌合成具有生防作用的次生代谢产物相关的基因簇
3.2 其他生防相关基因的研究
贝莱斯芽孢杆菌除了次生代谢产物以外,还有一些基因也是贝莱斯芽孢杆菌具有生防作用的因素。其中最为典型的是β-1,3-1,4-葡聚糖基因。为了解B.velezensis的β-1,3-1,4-葡聚糖酶对真菌的作用机制,XU 等[25]克隆了该酶的 β-1,3-1,4-葡聚糖酶基因,发现其能在大肠杆菌BL21中高表达,且该重组蛋白可以破坏真菌菌丝的形态而使其致病力降低,表明该酶具有较大的应用潜力。
4 贝莱斯芽孢杆菌的应用
近年来,越来越多贝莱斯芽孢杆菌的作用被发掘,除生防作用以外,贝莱斯芽孢杆菌还具有良好的固氮作用,它能促进养分的吸收和生物膜的形成、促进细胞生长、诱导系统性抗性,对烟嘧磺隆、苯磺隆、吡嘧磺隆、砜嘧磺隆、甲嘧磺隆等磺酰脲类除草剂也有良好的降解效果[26]。因此,贝莱斯芽孢杆菌在农业、工业以及生物医学上的应用也越来越广泛。
4.1 贝莱斯芽孢杆菌在农业上的应用
贝莱斯芽孢杆菌在农业上有着广泛的应用,是常见生防菌之一。从土壤中分离出来的贝莱斯芽孢杆菌 Jnb16菌株能有效的抑制水稻稻瘟病菌的生长,并对水稻具有良好的促生作用[27]。贝莱斯芽孢杆菌 MT310对烟草赤星病菌(Alternaria alternata)、黄连根腐病菌(Fusariumsp.)、石榴干腐病菌(Phomopsissp.)、高粱叶斑病菌(Phyllosticta sorghina)和高粱炭疽病菌(Colletotrichum graminicola)等植物病原真菌有明显抑制效果,并对烟草赤星病菌的抑制效果最好,抑制率达49.96%[28]。
贝莱斯芽孢杆菌的孢子不仅具有耐保存的特性,而且能够耐受极端的外界环境,因此容易被制作成生物制剂。冯江鹏等[29]的研究结果显示,贝莱斯芽孢杆菌JK3菌株含有脂肽类抗菌物质生物合成相关基因srfAA、bymB、fenD、bacA和ituC,提前24 h或接种胶孢炭疽菌孢子的同时喷施JK3发酵上清液对草莓炭疽病的防治效果超过 95%,表明贝莱斯芽孢杆菌有开发为防治胶孢炭疽菌生防制剂的潜力。贝莱斯芽孢杆菌AH2菌株,已经作为植物杀菌剂投入生产应用,可用于防治白粉病、灰霉病、纹枯病、菌核病、晚疫病等[30],并且AH2菌株的生物制剂对烟草赤星病的防治效果与化学防治效果无显著差异。除此之外,贝莱斯芽孢杆菌FZB42菌株能产生具有抑菌作用的次生代谢产物,能够产生生物膜,具有高效的根部定殖能力,可以有效帮助植物抵御病原微生物的侵染,已经作为生物肥料和生物防治试剂在农业领域被广泛应用[31]。截止到2021年3月底,在中国农药信息网(http://www.chinapesticide.org.cn/hysj/index.jhtml)登记的有效成分为甲基营养型芽孢杆菌(现已被归类为贝莱斯芽孢杆菌)有4条记录,暂无有效成分为贝莱斯芽孢杆菌的记录。其中甲基营养型芽孢杆菌LW-6主要剂型为母药和可湿性粉剂,主要防治柑橘溃疡病、黄瓜细菌性条斑病和水稻细菌性条斑病。甲基营养型芽孢杆菌9912的主要剂型也是母药和可湿性粉剂,主要防治草莓根腐病、黄瓜灰霉病和苹果树腐烂病。
目前,贝莱斯芽孢杆菌作为生防菌的应用也存在一些问题。大多数已被分离的菌株的防治机理暂未明确,大田防效暂未得到验证,也并未得到规模化的生产和应用,这些问题还需在后续的研究和应用中解决。生物制剂的剂型、使用方式和使用周期对药效都有影响,因此在应用中还要注意贝莱斯芽孢杆菌生物制剂的生产和使用方式。
4.2 贝莱斯芽孢杆菌在工业上的应用
研究发现,贝莱斯芽孢杆菌含有丰富的蛋白酶和纤维素酶,而这两类酶在工业酶中占有很大的比例。如贝莱斯芽孢杆菌157菌株内含有大量的内切纤维素酶、木质素酶和果胶酶等,其中内切纤维素酶的含量最高,在饲料添加剂、洗涤剂和造纸行业具有良好的应用前景[32]。贝莱斯芽孢杆菌P11菌株具有显著的角蛋白溶解活性、蛋白酶水解活性和纤维素酶活性[33-34],在工业生产上具有应用潜力。甲基营养型芽孢杆菌(现已归属于贝莱斯芽孢杆菌)FBKL1.0190[35]能产生高活性的中性蛋白酶、纤维素酶等,可应用于水产业和纺织业。张德峰等[36]通过试验发现,贝莱斯芽孢杆菌LF01菌株含有大量抗菌合成物质合成基因簇,其抗菌活性物质具有较高的生物安全性,能显著提高尼罗罗非鱼对无乳链球菌的抗病力,该菌株产生的抗菌活性物质可用于水产动物细菌病的防控。贝莱斯芽孢杆菌还可用于降解有毒有害的工业副产品,例如处理屠宰场的废水、降解纺织业垃圾等。此外,贝莱斯芽孢杆菌具有将染料DR28降解为联苯胺和4-氨基联苯的能力,这两种物质都是有效的诱变剂。它们会在短时间内进一步降解,降低染料的毒性和致突变性。因此,贝莱斯芽孢杆菌还可应用于偶氮染料的脱色和解毒[37]。
4.3 贝莱斯芽孢杆菌在生物医学上的应用
芽孢杆菌及其胞外多糖(Exopolysaccharide,EPS)可以调节肠道微生态平衡,改善机体免疫能力,对医疗保健、生物纳米技术等行业具有特殊价值。MOGHANNEM 等[38]使用响应面法对 M7,E1和9I三个菌株进行筛选得出结论:编号为9I的菌株(鉴定为贝莱斯芽孢杆菌KY498625菌株)可生产相对最多的胞外多糖,易于使用二乙氨基乙基纤维素柱(DEAE-Cellulose column)进行提取和纯化,且这种胞外多糖易于被生物降解,方便被应用于健康和生物纳米技术部门、食品和化妆品行业的胶凝剂以及环境部门的生物絮凝剂,被认为是环境可持续方面的重大突破。从贝莱斯芽孢杆菌MHM3菌株中提取出的EPS化合物在极低浓度下就具有极高的抗癌能力,且对健康的宿主细胞没有明显的毒性[39]。此外,从贝莱斯芽孢杆菌 RA5401中提取的五种化合物对乳腺癌细胞有抗增殖活性,其中两种通过抑制细胞内癌蛋白酶发挥作用,其余三种通过抑制癌细胞的G蛋白偶联受体发挥作用[40]。YOO等[41]从某种韩国食品中提取出了贝莱斯芽孢杆菌K68菌株,它可以通过脱氧野尻霉素(Deoxynojirimycin,DNJ)的产生抑制变异链球菌生物膜的形成、粘附和GTF基因的表达来预防龋齿。随着研究的不断深入,贝莱斯芽孢杆菌在生物医学方面更多的用途会被发掘。
5 小结
贝莱斯芽孢杆菌广泛分布于自然界,它作为生防菌是近年来的研究热点。2016年以来,贝莱斯芽孢杆菌引起了国内外学者的广泛关注,发表的研究论文数量呈明显上升态势。其中研究主要集中在分离鉴定、生防作用以及基因功能等方面,对适宜特定环境菌株的筛选、贝莱斯芽孢杆菌的生防机制、定殖能力等方面的报道较少。
在未来的研究中,应该通过大数据解析功能基因的调控网络,更加深入的研究贝莱斯芽孢杆菌与寄主生物互相作用的机制;通过设计不同的环境,进行适宜生存菌株的筛选,使其能更好的在特定的农业生态系统中发挥作用;通过荧光示踪方法分析贝莱斯芽孢杆菌在植物根、叶围等部位以及动物肠道中的定殖能力;挖掘与定殖相关基因(簇),通过分子遗传改良技术增强菌株的定殖能力,提高其应用价值。同时在实际应用的过程中,应更加重视其规模化的生产和使用、生物安全性的合理评定及监管。贝莱斯芽孢杆菌对绿色农业的发展有重大意义,其在工业、生物医学等方面的应用潜力也不容小觑,相信贝莱斯芽孢杆菌很快将在农业、工业、生物医学等行业发挥重要作用。
猜你喜欢
流程工业(2022年11期)2022-11-30 06:51:12
农药科学与管理(2019年6期)2019-11-23 08:17:12
农药科学与管理(2019年6期)2019-11-23 08:17:06
农药科学与管理(2019年8期)2019-11-23 08:04:46
疯狂英语·新策略(2019年8期)2019-10-18 00:59:28
现代园艺(2017年23期)2018-01-18 06:57:48
当代化工研究(2016年7期)2016-03-20 16:21:54
中国酿造(2016年12期)2016-03-01 03:08:15
湖南农业科学(2014年1期)2014-02-27 14:25:50
教育与职业(2014年1期)2014-01-14 02:56:34
