
种子引发剂氯化胆碱和吲哚丁酸钾对盐胁迫下水稻幼苗生长和生理特性的影响
2023-01-14 08:00:10余明龙冯乃杰郑殿峰
核农学报 2023年1期
黄 露 余明龙 冯乃杰 郑殿峰 揭 茵 李 瑶,3
(1广东海洋大学滨海农业学院,广东 湛江 524088;2广东海洋大学深圳研究院,广东 深圳 518108;3黑龙江八一农垦大学农学院,黑龙江 大庆 163319)
由NaCl 引起的高水平盐胁迫是作物生产中主要的环境胁迫因素之一[1-2]。据第二次全国土壤普查数据统计,除海滨滩涂外,我国盐渍土面积高达3 487万公顷,其中仍有近1 333 万公顷的土地具备农业耕种潜力[3]。水稻(Oryza sativaL.)是世界一半以上人口碳水化合物的主要来源。与其他谷类作物相比,水稻更易受到根际盐分的影响,特别是处于营养生长的苗期水稻对盐分胁迫高度敏感[4-5]。盐胁迫通过减少根对水分和养分的吸收,加速叶绿素分解,同时降低光合同化效率进而抑制水稻的生长[6]。此外,Na+长期积累造成的离子毒害以及活性氧(reactive oxygen species,ROS)稳态的失衡可诱导酶活性降低[7]和膜脂过氧化作用加剧[8],严重时会与重要的生物分子如脂质、蛋白质和核酸发生反应,造成细胞死亡[9]。因此,增强水稻幼苗的耐盐性对提高盐碱地利用率和提升水稻产量具有重要意义。
种子引发是由Heydecker 等[10]于1973年提出的一项控制种子缓慢吸水后逐步回干的种子处理技术,能够提高种子的出苗整齐度,并能在植株响应环境胁迫时表现出更快更强的细胞防御反应[11]。氯化胆碱(choline chloride,CC)是一种季胺碱,进入植物体后转化为磷脂酰胆碱,从而对膜系统具有修复作用。前人研究表明,外源施用CC 通过降低膜脂过氧化、抑制叶绿素分解、增加脯氨酸和甜菜碱积累来缓解盐胁迫对植物的损伤[12-13]。吲哚丁酸钾(indole-3-butyric acid potassium salt,IBAK)是一种促生根类植物生长调节剂,能够促进细胞分裂,诱导不定根的生成[14]。近期研究发现IBAK 拌种能够增强不同耐旱性大豆的耐旱能力[15]。然而,有关引发剂CC 和IBAK 对盐胁迫下水稻幼苗生长和生理代谢的影响尚鲜有报道。因此,本研究以杂交稻湘两优900 和常规稻黄华占为试验材料,探讨引发剂CC 和IBAK 对盐胁迫下水稻幼苗表型、ROS 代谢、细胞膜损伤和渗透调节的影响,明确CC 和IBAK 影响水稻苗期耐盐性的生理机制,以期为种子引发技术应用于耐盐水稻生产提供理论参考。
1 材料与方法
1.1 试验材料
供试水稻品种为湘两优900(杂交稻)和黄华占(常规稻),分别由湖南杂交水稻中心和广东省农业科学院提供;供试引发剂为氯化胆碱(CC)和吲哚丁酸钾(IBAK),均由广东海洋大学化控实验室提供。
1.2 试验方法
1.2.1 适宜引发剂浓度筛选试验 试验于2021年5月在广东海洋大学滨海农业学院人工气候室进行,供试水稻品种为黄华占。选取籽粒饱满的水稻种子,经0.5%次氯酸钠消毒10 min 后,用蒸馏水冲洗3 次。随后将种子置于不同浓度CC(0.75、1.5、3.0、6.0 和12.0 mg·L-1)和IBAK(0.25、0.5、1.0、2.0 和4.0 mg·L-1)溶液中,于25 ℃PGX-450D 无光照人工气候培养箱(宁波赛福实验仪器有限公司)中引发24 h,以蒸馏水引发为对照(CK),引发结束后将种子用蒸馏水洗净,再用滤纸吸干水分,随后于25 ℃的JIDI-9240A 鼓风干燥箱(广州吉迪仪器有限公司)内回干至初始含水量后用于后续试验。选取水稻种子点播于上直径12.0 cm、下直径7.5 cm、高5.5 cm 的塑料盆中,每盆均匀播种30 粒,以砖红壤∶沙=3∶1 混合作为培养基质,每盆装土0.3 kg,然后转移至人工气候室中培养(光周期为光照16 h/黑暗8 h,白天温度为25 ℃,夜间温度为18 ℃,湿度为60%),正常水分管理至两叶一心期,取样测定株高、茎基宽、叶面积、地上干重和根干重,根据壮苗指数[16]筛选适宜的引发剂浓度。壮苗指数计算公式如下:

1.2.2 种子引发对NaCl 胁迫下水稻幼苗生长的调控效应试验 试验于2021年7—10月在广东海洋大学日光联动温室进行,供试水稻品种为湘两优900 和黄华占,根据1.2.1 的试验结果,选用3.0 mg·L-1CC 和1.0 mg·L-1IBAK 作为适宜引发浓度。将消毒后的两水稻品种各随机分成三组,分别用蒸馏水、CC 和IBAK各引发24 h,随后的清洗和回干条件同1.2.1。选取水稻种子点播于上直径19.0 cm、下直径14.5 cm、高18.0 cm 的不漏水塑料盆中,每盆均匀播种75粒,每盆装土3.0 kg,土壤成分同1.2.1,随后转移至温室中进行培养,自然光照,正常水分管理至三叶一心期。
于三叶一心期将上述两水稻品种的各三组处理再各随机分成两组,一半仍保持清水浇灌,一半用浓度为0.3%的NaCl 溶液进行浇灌,浇灌时用AR8012A 盐度计(东莞万创电子制品有限公司)控制NaCl 浓度,使NaCl 处理的土壤水层最终含NaCl 量为0.3%,仅在三叶一心期浇灌一次NaCl 溶液,水层深度为2 cm,盆壁标记水位线,此后每隔1 d补充清水至水位线并通过盐度计监测Nacl 浓度。每个品种各设置如下6 个处理:CK(蒸馏水引发+清水浇灌),CC(CC引发+清水浇灌),IBAK(IBAK 引发+清水浇灌),NaCl(蒸馏水引发+0.3% NaCl 浇灌),NaCl+CC(CC 引发+0.3% NaCl 浇灌)和NaCl+IBAK(IBAK 引发+0.3% NaCl 浇灌),每个处理各16盆。试验采用完全随机区组设计,3次重复,生长至四叶一心期对叶片进行取样,样品用液氮速冻,保存于-40 ℃冰箱待测。
1.3 测定项目与方法
1.3.1 形态指标测定 于四叶一心期在各处理随机选取60株水稻幼苗,用去离子水充分洗净,再用滤纸吸干水分,选取其中30株使用直尺测量株高,游标卡尺测量茎基宽,Yaxin-1241叶面积仪(北京博伦经纬科技发展有限公司)扫描测定叶面积。随后将幼苗置于烘箱中105 ℃杀青30 min,75 ℃烘干至恒重后测量地上干重和根干重。另外30 株用于测量根系总长度,先将根系通过台式扫描仪(日本Epson Experssion 公司)进行扫描,再经WinRHIZO 2.3.2 根系分析软件分析得到根系总长度。
1.3.2 叶绿素含量和气体交换参数测定 于四叶一心期采用SPAD-502 手持式叶绿素仪(日本Minolta 公司)测定倒二叶的叶绿素含量(soil and plant analyzer development,SPAD);采用LI-6400XT 便携式光合作用测定系统(美国LI-COR 公司)于上午9:00—11:30 测定倒二叶的气体交换参数,包括净光合速率(net photosynthetic rate,Pn)、气孔导度(stomatal conductance,Gs)、胞间CO2浓度(intercellular CO2concentration,Ci)和蒸腾速率(transpiration rate,Tr),各处理重复测定8次。
1.3.4 生理生化指标测定 于四叶一心期对叶片进行取样,参照《植物生理学实验指导》[18]的方法测定超氧化物歧化酶(superoxide dismutase,SOD)、过氧化物酶(peroxidase,POD)、过氧化氢酶(catalase,CAT)和抗坏血酸过氧化物酶(ascorbate peroxidase,APX)活性;参照Feng 等[2]的方法测定过氧化氢(hydrogen peroxide,H2O2)含量;丙二醛(malondialdehyde,MDA)含量采用硫代巴比妥酸法[19]进行测定;相对电导率(relative electrical conductivity,REC)参照Jia 等[20]的方法进行测定;抗坏血酸(reduced ascorbic acid,AsA)含量采用4,7-二苯基-1,10-菲咯啉显色法[21]进行测定;谷胱甘肽(reduced glutathione,GSH)含量采用5,5-二硫代双-2-硝基苯甲酸显色法[21]进行测定;参照Bradford[22]的方法采用考马斯亮蓝法测定可溶性蛋白含量;脯氨酸含量采用磺基水杨酸法[23]进行测定。
1.4 数据处理与统计分析
使用SPSS 26 软件进行方差分析,采用Duncan’s新复极差法进行差异显著性检验,使用Origin 9.0软件绘图。
2 结果与分析
2.1 不同浓度引发剂CC和IBAK对水稻幼苗生长的影响
由表1可知,与CK相比,3.0 mg·L-1CC和1.0 mg·L-1IBAK 处理均显著降低了黄华占水稻幼苗的株高,而显著增加了茎基宽、根干重和壮苗指数,且壮苗指数分别在3.0 mg·L-1CC 和1.0 mg·L-1IBAK 处理时达到最大值。因此,本研究选取3.0 mg·L-1CC 和1.0 mg·L-1IBAK作为适宜引发浓度。
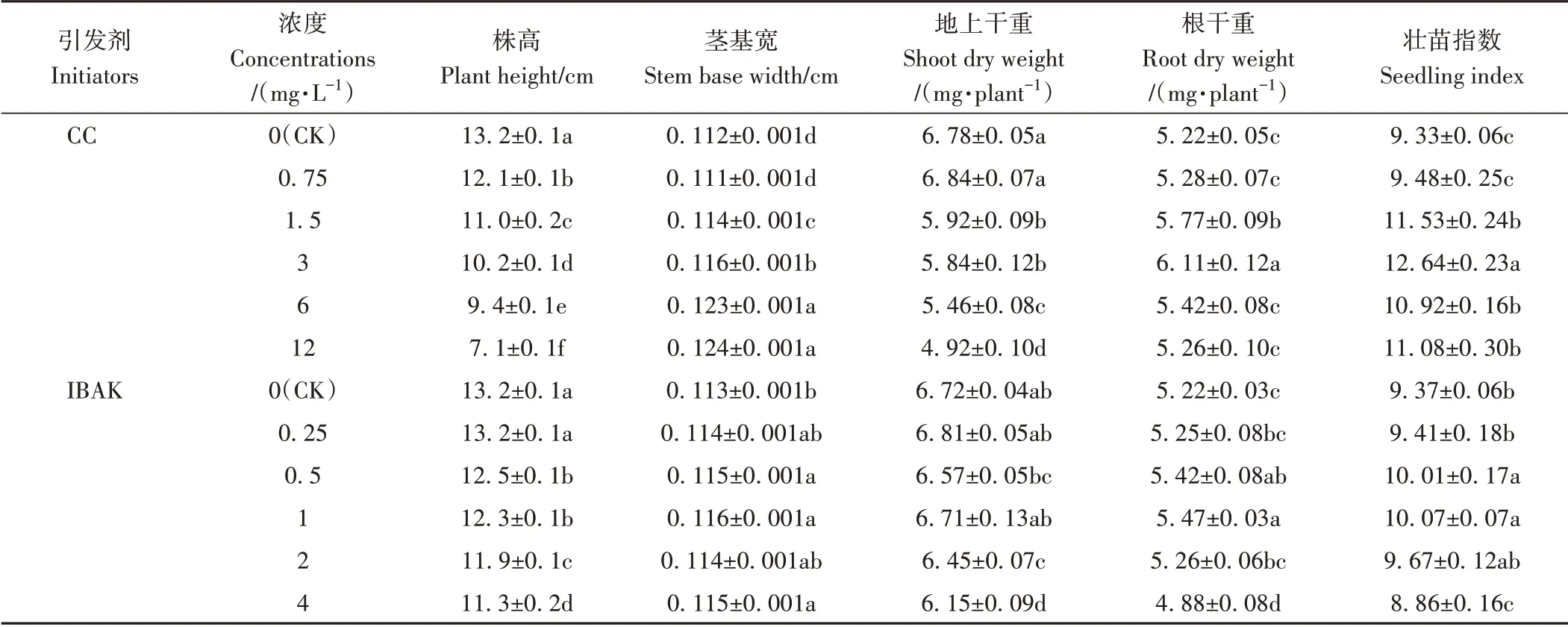
表1 不同浓度引发剂CC和IBAK对黄华占幼苗生长的影响Table 1 Effects of different concentrations of initiators CC and IBAK on the growth of Huanghuazhan seedlings
2.2 引发剂CC 和IBAK 对NaCl 胁迫下水稻幼苗生长的影响
由表2、3 可知,NaCl 胁迫使黄华占的所有生长指标均较CK 显著降低,同时抑制了湘两优900株高和茎基宽的增加。与NaCl 处理相比,NaCl+CC 处理进一步降低了盐胁迫下两品种水稻的株高,但显著提高了湘两优900 的根干重以及黄华占除株高外的所有生长指标;NaCl+IBAK 处理只降低了盐胁迫下湘两优900 的株高,而使黄华占的根系总长度、叶面积、地上干重、根干重和壮苗指数均得到显著提高。非盐胁迫条件下,与CK 相比,两种引发剂处理均能显著降低两品种水稻的株高,CC 处理显著提高了湘两优900 的茎基宽和叶面积以及黄华占的茎基宽、地上干重、根干重和壮苗指数;IBAK 处理对两品种根系总长度均有促进作用,同时显著增加了湘两优900 的叶面积。双因素方差分析结果表明,盐和引发剂的交互作用显著影响了黄华占的株高、叶面积和根干重(P<0.05),极显著影响了壮苗指数(P<0.01)。
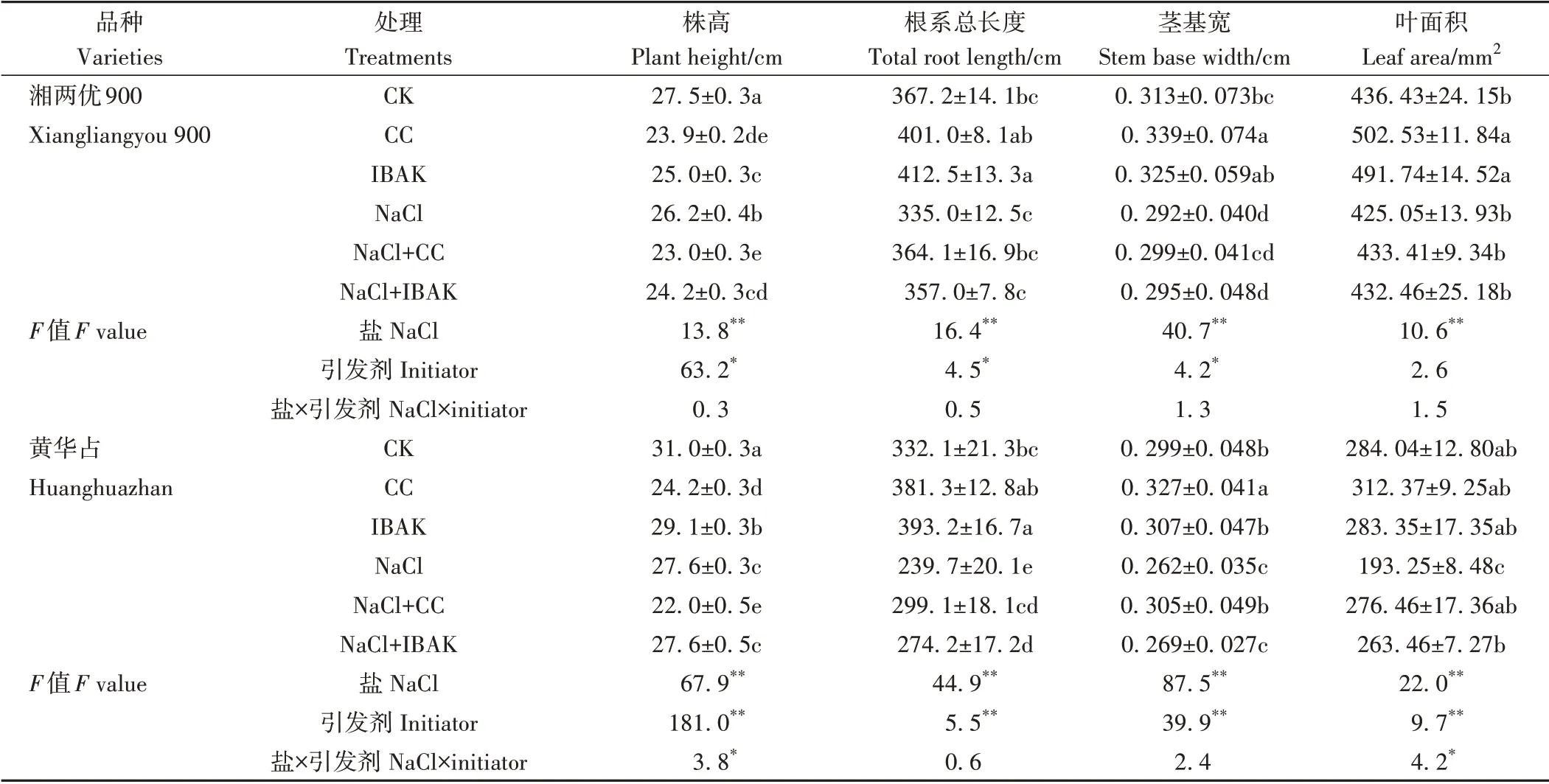
表2 引发剂CC和IBAK对NaCl胁迫下水稻幼苗生长的影响Table 2 Effects of initiators CC and IBAK on the growth of rice seedlings under NaCl stress
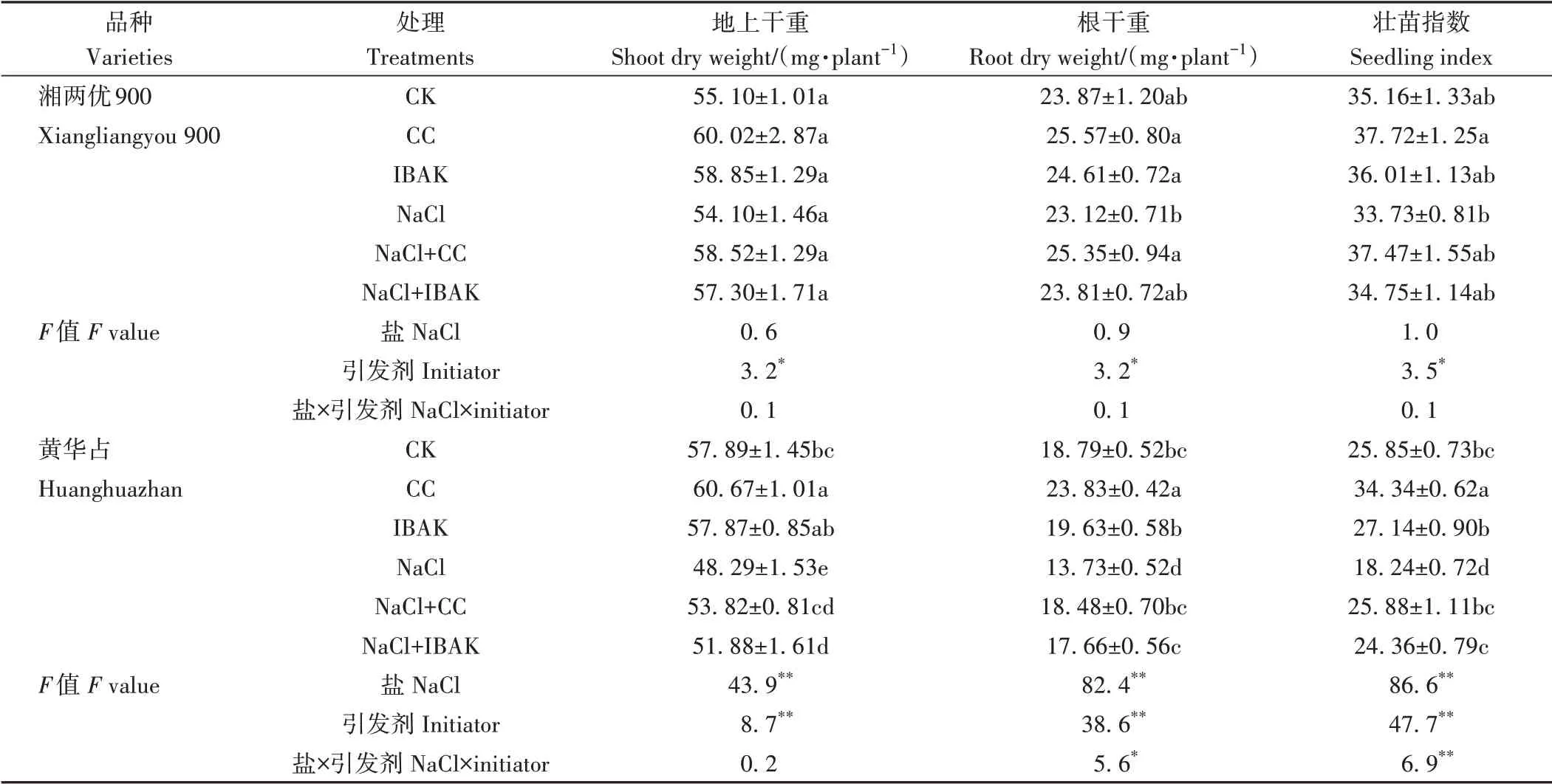
表3 引发剂CC和IBAK对NaCl胁迫下水稻幼苗生长的影响Table 3 Effects of initiators CC and IBAK on the growth of rice seedlings under NaCl stress
2.3 引发剂CC 和IBAK 对NaCl 胁迫下水稻叶片光合特性的影响
由表4可知,与CK 相比,NaCl 处理均能显著降低两品种水稻叶片的Gs、Tr 和SPAD 值,而Pn 和Ci 仅在黄华占中显著降低。与NaCl处理相比,除对Ci均无显著影响外,NaCl+CC 和NaCl+IBAK 处理均能显著提高盐胁迫下湘两优900 和黄华占叶片的Pn、Gs、Tr 和SPAD 值,且引发效果表现为CC优于IBAK。非盐胁迫下,引发剂CC和IBAK处理与CK相比也能显著提高湘两优900 的Pn 和Gs。方差分析结果表明,盐和引发剂的交互作用对湘两优900 的Pn 和SPAD 值存在显著影响(P<0.05),而对黄华占的Pn、Tr和SPAD值存在极显著影响(P<0.01)。

表4 引发剂CC和IBAK对NaCl胁迫下水稻叶片光合特性的影响Table 4 Effects of initiators CC and IBAK on photosynthetic characteristics in rice leaves under NaCl stress
2.4 引发剂CC 和IBAK 对NaCl 胁迫下水稻叶片膜损伤和ROS含量的影响
由图1可知,与CK 相比,NaCl 处理下两品种水稻叶片的REC、MDA和H2O2含量均显著提高,同时组织化学染色显示叶片中染色强度明显增加。与NaCl 处理相比,NaCl+CC处理湘两优900和黄华占叶片的REC分别显著降低7.98 和11.08 个百分点,H2O2含量分别显著降低12.43%和21.67%;NaCl+IBAK 处理湘两优900和黄华占叶片的REC分别显著降低7.44和3.28个百分点,H2O2含量分别显著降低15.56%和17.60%;NaCl+CC和NaCl+IBAK处理下湘两优900的MDA含量均无显著差异,而黄华占的MDA含量分别较NaCl处理显著降低29.68%和20.49%。此外,与NaCl 处理相比,NaCl+CC 和NaCl+IBAK 处理均明显降低了两品种水稻叶片中的染色强度。
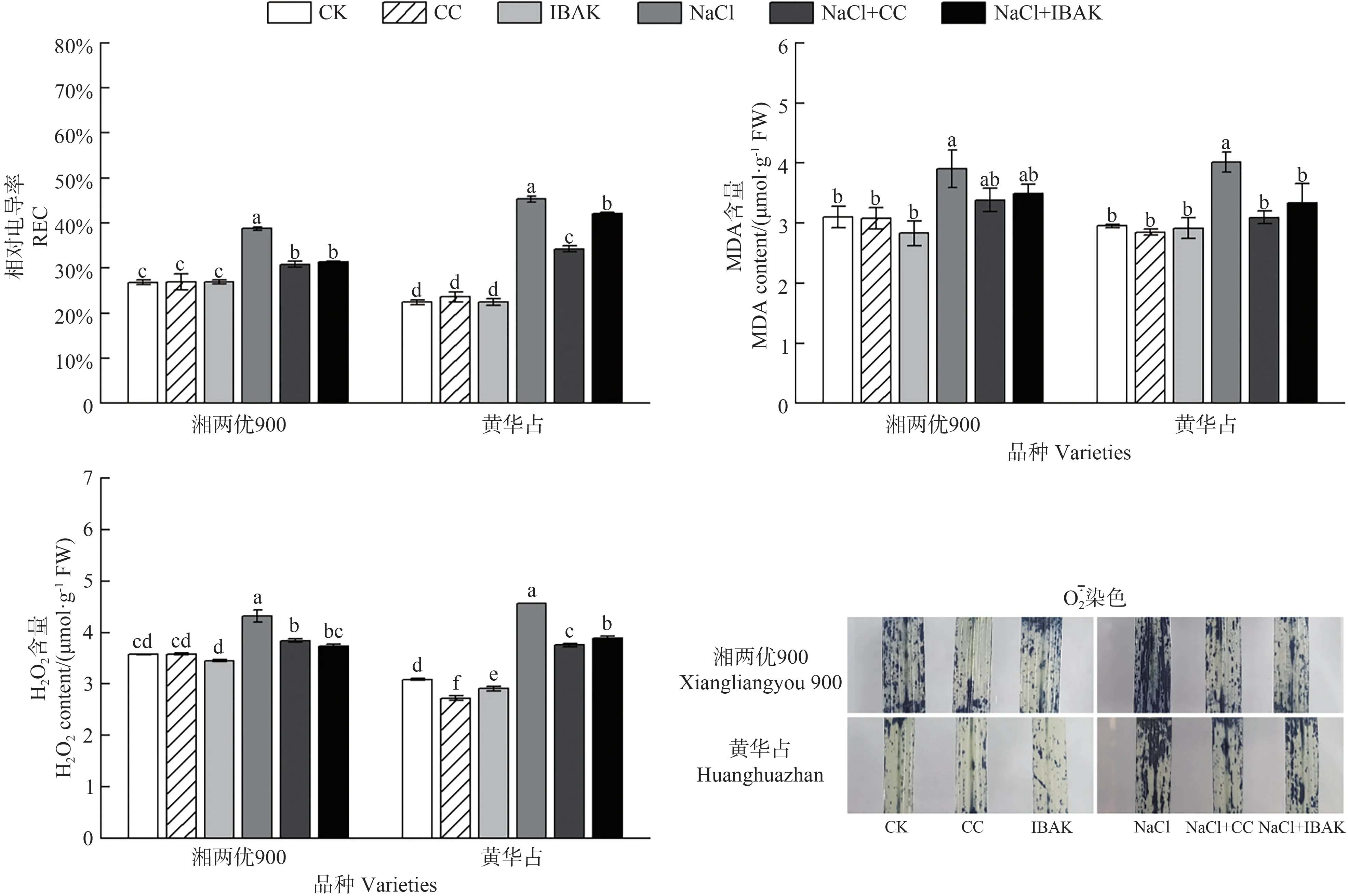
图1 引发剂CC和IBAK对NaCl胁迫下水稻叶片膜损伤和ROS含量的影响Fig.1 Effects of initiators CC and IBAK on membrane damage and ROS content in rice leaves under NaCl stress
2.5 引发剂CC 和IBAK 对NaCl 胁迫下水稻叶片抗氧化酶活性的影响
由图2可知,与CK 相比,NaCl 处理两品种水稻叶片的SOD、CAT 和APX 活性均显著降低;POD 活性在湘两优900 中显著增强,而在黄华占中显著降低。与NaCl 处理相比,盐胁迫下引发剂处理进一步增强了水稻叶片的抗氧化酶活性,表现为NaCl+CC 和NaCl+IBAK 处理黄华占叶片的SOD、POD、CAT 和APX 活性分别显著增加100.03% 和80.61%、29.49% 和26.47%、11.87% 和10.19%、51.10% 和22.97%;同时,NaCl+CC 和NaCl+IBAK 处理湘两优900 叶片的SOD 活性分别显著增加92.67%和82.17%,而盐胁迫下湘两优900叶片的CAT和APX活性仅在NaCl+CC处理后显著提高。此外,非盐胁迫下,与CK 相比,CC 处理显著增强了湘两优900 叶片的POD 活性,IBAK 处理显著增强了湘两优900叶片的POD和CAT活性。

图2 引发剂CC和IBAK对NaCl胁迫下水稻叶片抗氧化酶活性的影响Fig.2 Effects of initiators CC and IBAK on the activities of antioxidant enzymes in rice leaves under NaCl stress
2.6 引发剂CC 和IBAK 对NaCl 胁迫下水稻叶片非抗氧化剂含量的影响
由图3可知,与CK 相比,NaCl 处理湘两优900 叶片的AsA 和GSH 含量无显著差异,而黄华占叶片的AsA 和GSH 含量则分别显著降低36.32%和35.23%。与NaCl 处理相比,NaCl+CC 和NaCl+IBAK 处理黄华占叶片的非抗氧化剂水平进一步提高,AsA 含量分别显著增加52.14%和23.76%,GSH 含量分别显著增加51.64%和39.85%;NaCl+CC 和NaCl+IBAK 处理湘两优900 叶片的AsA 含量分别较NaCl 处理显著提高34.65%和21.75%,而GSH 含量仅在NaCl+CC 处理后显著增加,增幅为32.69%。与CK 相比,非盐胁迫下CC 处理湘两优900 的AsA 含量和黄华占的GSH 含量均显著增加。

图3 引发剂CC和IBAK对NaCl胁迫下水稻叶片非抗氧化剂含量的影响Fig.3 Effects of initiators CC and IBAK on the content of non-antioxidants in rice leaves under NaCl stress
2.7 引发剂CC 和IBAK 对NaCl 胁迫下水稻叶片渗透调节物质含量的影响
由图4可知,与CK 相比,NaCl、NaCl+CC 和NaCl+IBAK 处理湘两优900 叶片的可溶性蛋白含量均无显著差异。NaCl 处理黄华占叶片可溶性蛋白含量较CK显著降低6.24%,而NaCl+CC 和NaCl+IBAK 处理促进了黄华占叶片可溶性蛋白含量的积累,分别较单独盐处理显著增加7.37%和3.44%。与CK 相比,盐胁迫下有无引发剂处理两品种水稻叶片脯氨酸含量均显著增加,但盐胁迫加引发剂处理较单独盐胁迫积累了更高水平的脯氨酸,NaCl+CC 和NaCl+IBAK 处理脯氨酸含量分别较单独盐处理增加了37.14%和55.71%(湘两优900)、47.87%和29.32%(黄华占)。此外,非盐胁迫下,两种引发剂处理与对照相比也显著增加了两水稻品种叶片的脯氨酸含量,而可溶性蛋白含量仅在湘两优900中显著增加。

图4 引发剂CC和IBAK对NaCl胁迫下水稻叶片渗透调节物质含量的影响Fig.4 Effects of initiators CC and IBAK on the content of osmotic regulatory substances in rice leaves under NaCl stress
2.8 引发剂对NaCl胁迫下水稻幼苗生理指标的双因素方差分析
由表5可知,盐或引发剂对两品种水稻的REC、SOD 活性、H2O2、GSH、可溶性蛋白和脯氨酸含量均存在极显著影响(P<0.01),同时盐胁迫极显著影响了两品种的MDA含量(P<0.01),引发剂极显著影响了两品种的POD 活性和AsA 含量(P<0.01);盐和引发剂的交互作用对湘两优900 的REC、SOD、POD 活性、H2O2、GSH 和脯氨酸含量存在极显著影响(P<0.01),对黄华占的REC、SOD 活性、POD 活性、H2O2含量和可溶性蛋白含量存在极显著影响(P<0.01),而对黄华占的APX活性存在显著影响(P<0.05)。

表5 引发剂对NaCl胁迫下水稻幼苗生理指标的双因素方差分析Table 5 Two-way ANOVA analysis of initiator on physiological indexes of rice seedlings under NaCl stress
3 讨论
土壤盐分是限制植株生长的主要因素之一,植株在成苗的早期阶段更容易受到盐胁迫的影响,盐胁迫引起的高渗透压和离子毒害通过诱导养分亏缺以及ROS 过量积累而干扰正常的生理代谢过程,进而抑制植株生长[24]。李瑶等[16]研究发现盐胁迫可显著降低水稻幼苗的株高并抑制生物量的积累。本研究结果表明,盐胁迫降低了湘两优900 的株高和茎基宽,而对黄华占的所有生长参数均有显著的抑制作用,说明盐胁迫对水稻生长具有普遍的不利影响,且对常规稻黄华占的抑制作用更强,杂交稻湘两优900 可能具备更强的抗氧化和渗透调节能力,有利于清除ROS,维持光合活性,这与Huang 等[25]在水稻中的研究结果一致。种子引发是一种经济高效且环境友好的农业生物技术,已被广泛应用于农业生产以改善各种作物的生长和抗逆性,被引发的植物在响应胁迫时会产生更快、更强的防御反应[7]。本试验中,引发剂CC 处理进一步增加了盐胁迫下湘两优900 的根干重,并对除株高外的黄华占生长参数均具有显著的促进效果;引发剂IBAK 处理降低了盐胁迫下湘两优900 的株高,但显著增加了黄华占的根系总长度、叶面积、地上干重、根干重和壮苗指数,而盐和引发剂处理仅在黄华占的形态指标间存在交互作用。说明适宜浓度的CC和IBAK处理能够缓解盐胁迫对水稻幼苗生长的抑制,且对常规稻黄华占的调控效果更为明显。
干物质积累需要光合作用固定CO2,光合作用依赖于叶绿素对光的吸收、传递和转化[26]。盐胁迫下,Na+在叶片组织中的过度积累导致叶绿素分解速率加快或阻碍叶绿素的生物合成,进而导致叶片光合活性降低[27]。本研究结果表明,盐胁迫下两品种水稻SPAD值、Gs和Tr均受到显著抑制,而Pn 和Ci仅在黄华占中显著降低,表明在逆境胁迫下,植物通过关闭气孔,同时降低蒸腾作用以减少水分流失,黄华占叶片Pn显著降低可能是由于其气孔关闭较湘两优900更为明显,进而造成光合碳固定底物供应不足,导致Pn下降[28]。同时,盐胁迫下黄华占的Ci与Gs同步降低,表明盐胁迫下Pn下降的主导因素可能归因于气孔限制,这与Medrano等[29]的研究结果相一致。此外,Abideen等[30]研究发现盐胁迫下更低的Tr 有利于Na+在根部停留,这是植物在逆境胁迫中的一种适应性策略。本研究发现,与NaCl 处理相比,NaCl+CC 和NaCl+IBAK 处理均能显著缓解盐胁迫对两品种水稻叶片气体交换参数的抑制(除Ci 增加不显著外),Ci 和Gs 同时增加进一步表明NaCl+CC和NaCl+IBAK处理增强Pn可能是气孔导度增加促进CO2扩散的结果[2]。前人研究表明,CC 和IBAK处理能提高叶绿体膜结构的稳定性,维持叶片较高的叶绿素含量,从而对盐胁迫下水稻较高的光合速率有一定促进作用[31]。刘美玲等[32]研究发现IBAK 浸种可增加干旱胁迫下大豆叶片的叶绿素含量。周峰等[33]研究发现外源施用CC 能提高低盐胁迫下菠菜叶片的光合活性。本研究同样发现引发剂CC和IBAK处理在水稻耐盐生长中起着积极作用,这可能归功于其对光合参数的改善以及对光合色素的保护作用。
逆境胁迫下过量积累的ROS 会诱导细胞膜脂过氧化作用加强进而造成膜透性增加[34]。引发处理能增强相关抗氧化酶活性,同时增加非抗氧化剂水平以协调清除自由基,促进细胞膜的修复和膜完整性的提高[35]。本研究发现,与CK 相比,NaCl处理显著增加了两品种水稻叶片REC 水平、O-·2染色强度、MDA 和H2O2含量,降低了SOD、CAT 和APX 活性,POD 活性在湘两优900 中显著增加而在黄华占中显著降低,同时非抗氧化剂AsA 和GSH 含量在黄华占中显著降低,说明盐胁迫通过抑制抗氧化系统活性而加剧细胞的氧化损伤,且常规稻黄华占的抗氧化系统对盐胁迫表现更为敏感,杂交稻湘两优900 依赖于自身较强的抗氧化活性更有助于清除ROS,这与前人在大豆[36]和棉花[37]中的研究结果一致。而盐胁迫下NaCl+CC和NaCl+IBAK处理两品种水稻叶片的ROS 含量和膜损伤均有所降低,同时抗氧化酶活性和非抗氧化剂含量均不同程度提高,双因素方差分析中盐胁迫和引发剂的交互作用在两品种REC 及SOD、POD 活性、H2O2和GSH 含量中均存在极显著影响(P<0.01),表明引发处理通过增加抗氧化酶活性直接清除ROS 或抑制ROS 和MDA 的生成途径来缓解膜脂过氧化,进而增强了水稻幼苗在盐胁迫下的防御能力,但其具体调控机理仍需进一步深入研究。盐胁迫下NaCl+CC 和NaCl+IBAK 处理湘两优900 的REC 比黄华占更低,可能是由于引发处理诱导湘两优900 叶片脯氨酸大量积累,进一步增加了ROS清除速率,同时降低了细胞渗透失水。
促进渗透溶质的生物合成是植物抵御环境胁迫的另一个潜在机制,较高的渗透溶质含量有利于根系的水分吸收[5]。本研究结果表明,盐胁迫显著降低了黄华占叶片的可溶性蛋白含量,这与张翯等[38]在玉米中的研究结果相反,可能是由于可溶性蛋白降解有利于氨基酸生物合成,而脯氨酸是参与植物渗透调节的重要氨基酸[39],盐处理后两品种叶片脯氨酸显著积累也从侧面证明了这一点,表明盐胁迫破坏了叶片细胞功能,植物通过主动积累脯氨酸来维持细胞水势[40]。NaCl+CC和NaCl+IBAK 处理进一步增加了盐胁迫下两品种水稻叶片的脯氨酸含量,且在湘两优900中增幅更大,而可溶性蛋白含量仅在黄华占中有所增加,双因素方差分析结果同样表明盐和引发剂的交互作用对湘两优900 的脯氨酸含量以及黄华占的可溶性蛋白含量存在极显著影响(P<0.01),湘两优900中更多的氨基酸参与了脯氨酸的生成,而黄华占中更多的可溶性蛋白含量有利于促进细胞功能恢复。上述结果表明,引发剂处理可通过促进渗透保护剂的积累来恢复叶片功能、维持叶片渗透压、降低膜氧化损伤,促进盐胁迫下水稻幼苗的生长。
4 结论
本研究结果表明,盐胁迫抑制了水稻幼苗的生长,适宜浓度的引发剂CC和IBAK处理可增加盐胁迫下水稻叶片的叶绿素含量,同时改善气体交换参数来维持光合结构活性,从而刺激植株生长。另一方面,引发剂处理通过激活抗氧化酶活性,提高非抗氧化剂水平来清除ROS 并降低膜脂过氧化水平;同时促进渗透保护剂积累来维持叶片的渗透势,进而增强了水稻幼苗的耐盐性。
猜你喜欢
中老年保健(2022年3期)2022-11-21 09:40:36
小学阅读指南·教研版(2022年3期)2022-04-28 22:20:30
今日农业(2021年10期)2021-11-27 09:45:24
科学与财富(2019年15期)2019-10-21 13:35:48
发酵科技通讯(2018年2期)2018-07-06 11:38:46
湖南林业科技(2017年1期)2017-02-06 05:29:00
中国火炬(2015年12期)2015-07-31 17:38:35
湖南农业科学(2014年1期)2014-02-27 14:25:26
中国火炬(2013年3期)2013-07-24 14:15:06
中国火炬(2011年8期)2011-07-25 10:44:25
