
广西长寿饮食模式对人体肠道菌群结构的调节作用
2023-01-12 03:45韩坤宸窦君霞黄国宏于晓涵廖艳婷张钦任宋瑶马金克李全阳
现代食品科技 2022年12期
韩坤宸,窦君霞,黄国宏,于晓涵,廖艳婷,张钦任,宋瑶,马金克,李全阳*
(1.广西大学轻工与食品工程学院,广西南宁 530004)(2.泰山职业技术学院生物技术工程系,山东泰安 271000)(3.广西职业技术学院农业工程学院,广西南宁 530226)
肠道微生物作为“被遗忘的器官”,可参与体内能量平衡和代谢过程,并对改善机体炎症和免疫功能,调节机体健康状态起着重要作用[1]。过去的研究表明,肠道菌群失调与肥胖、心血管疾病、过敏性疾病、炎症性肠病等疾病密切相关[2,3]。因此,稳定肠道菌群的平衡对改善机体健康状态十分重要。饮食是影响肠道菌群组成的决定性因素之一。研究表明,即便短期饮食习惯的改变也可对肠道微生物产生很大影响。Cancello 等[4]的研究表明15 d 的低热量地中海饮食可降低肠道中的促炎菌属丰度,增加与“瘦”相关分类菌属的丰度;Bonder 等[5]发现4 周的无麸质饮食可以改变健康志愿者的肠道微生物组成。已有较多的研究试图通过饮食干预的手段来调节肠道菌群失衡,改变机体的健康状态。地中海饮食可增加代谢综合症患者粪便菌群中参与调控胰岛素水平的菌属,从而有效改善心血管疾病[6];能量限制饮食可有效降低机体C 反应蛋白(C-Reactive Protein,CRP)水平,影响微生物组成和丰度,继而改善机体代谢[7]。然而,受限于不同国家和地区主要作物和饮食习惯差异,高依从性地遵循某种饮食并在全球推广难度极大,且有关报道很少。因此,着眼于本土地区健康饮食开展研究所得成果,对帮助当地人群维持机体健康状态更具实用价值。
本着“问计于百岁老人,求健康长寿”的初心,多年来,本团队聚焦广西地区百岁老人,进行了大量膳食方面的调查和研究,构建了广西长寿饮食模式,又从饮食的角度对百岁老人长寿的成因进行了多方面的探索和研究。在对广西长寿地区百岁老人的饮食及代谢物特征、肠道菌群等方面研究后,得知其饮食模式以膳食纤维多糖为主,具有高膳食纤维、高维生素A、低能量、低脂肪、低蛋白质、低胆固醇的特征[8,9]。之后本团队又分别用D-半乳糖致衰老小鼠和自然衰老小鼠,从小鼠抗氧化损伤、认知能力与代谢组学等方面逐步对构建的广西长寿饮食模式进行了验证优化,发现所构建的创新饮食模式具有明显的降低炎症、延缓衰老的作用[10-12]。本研究在前期工作的基础上,试图从肠道菌群的视角,对既有饮食模式的益生作用进行阐释,并通过分析饮食干预所引起的肠道菌群变化评估科学饮食对机体健康的有益作用,以期为丰富我国膳食营养与抗衰老理论体系提供基础支持。
1 材料与方法
1.1 志愿者招募
筛选志愿者时,纳入标准为:身体状态稳定;未长期服用且干预前一个月未服用影响免疫系统的药物和抗生素;实验前两周内未食用益生元、益生菌或维生素;对所涉及的食材均无过敏现象。经过筛选,共有15 名志愿者参与了研究,并被告知试验中涉及的内容和潜在风险,签署了书面知情同意书。在饮食干预过程中,一名志愿者因依从性差而提前退出,故共有14 名志愿者(年龄50~75 岁)完成了整个研究,其中7 名男性,7 名女性。研究方案获得广西大学伦理委员会批准(No.GXU-2020-136)。
1.2 志愿者膳食调查
在广西长寿饮食干预前,参考王芳等[9]的方法,采用半定量膳食频率调查问卷法(Food Frequency Questionnaire,FFQ)分析了解志愿者的日常膳食摄入状况。调查内容包含23 种食物及食物组合。志愿者的食物摄入量计算方法为:

式中:
S——食物摄入量,g/d;
D——摄入频率,次;
J——每次进食量,g;
Z——进食周期,d。
1.3 仪器与设备
Qubit 荧光定量仪,美国Invitrogen 公司;Agilent 2100 bioanalyzer,美国安捷伦公司;MiSeq 测序仪,美国Illumina 公司。
1.4 方法
1.4.1 广西长寿饮食的优化
本研究在团队前期工作的基础上,结合《中国居民膳食指南2016 版》,《中国老年人膳食指南2010》和《广西居民膳食营养与健康状况报告》中各营养素推荐量,在能量、脂肪、蛋白质、碳水化合物和膳食纤维摄入量上对广西长寿饮食模式进行了设计和优化。其中:能量摄入以膳食指南推荐量为基础,结合团队前期动物实验的优化验证结果计算得出[10]。膳食纤维摄入量参考前期动物实验中的最佳剂量,结合对百岁老人的饮食调查结果设定[8,12]。蛋白质,脂肪,碳水化合物的摄入量均基于百岁老人饮食特点[8],结合《广西居民膳食营养与健康状况报告》和膳食指南推荐量优化设计,分别占能量摄入的15%、25%和60%。
每位志愿者的营养素摄入量要根据前期调查和测量得到的性别、体重、身体活动水平等信息而进行针对性地设计。在设计过程中,以BMI=21.25(18.5~24.0的中位数)为标准,以《中国居民膳食指南2016 版》,《中国老年人膳食指南2010》中推荐的不同性别、年龄、体力活动所需要的能量为基础,根据广西长寿饮食模式的设计要求,确定其总能量的摄入量。蛋白质、脂肪、碳水化合物和膳食纤维摄入量在能量的基础上根据广西长寿饮食模式中所设计的各营养素摄入量计算而得。对于每一位志愿者,在同性别、年龄、身体活动水平所需能量的基础上,设计所摄入的能量以其基础代谢率与标准BMI 下比值作为系数进行计算。同时,对于肥胖志愿者(BMI>28),设定了0.95 的肥胖系数以控制其总能量摄入。据此,14 名志愿者在饮食干预期间的日平均摄入各营养素量如表1 所示。饮食干预期间志愿者所摄入的食物是基于所设定的广西长寿饮食模式中的营养素量,根据百岁老人饮食习惯和广西长寿地区的特色食物,参考《中国食物成分表标准版》凝练细化而来。食材主要选择当地种植的季节性水果、蔬菜、谷物和肉类。所有食材都经过精确计量。
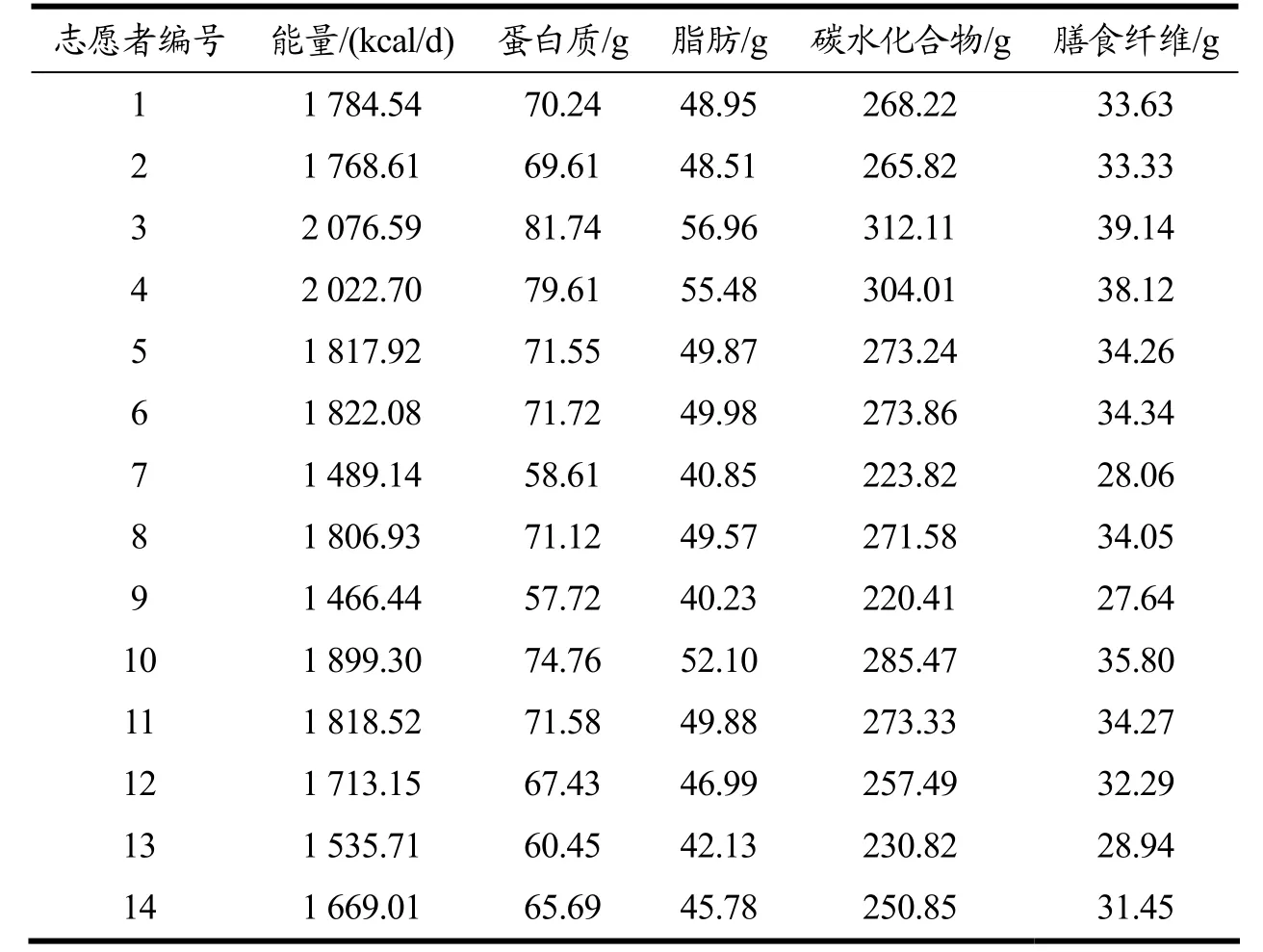
表1 饮食干预期间志愿者平均每日所摄入营养素量Table 1 Average daily nutrient intake of volunteers during dietary intervention
1.4.2 饮食干预方法
志愿者在饮食干预前被要求保持至少两周稳定的习惯性饮食,之后再接受为期2 周高依从性的广西长寿饮食模式干预。在广西长寿饮食模式干预前后分别采集志愿者的粪便样品并进行肠道菌群检测,具体实验流程如图1 所示。在饮食干预期间,志愿者所摄入的所有食物均由本团队提供,包括早餐、中餐、晚餐和水果等。且要求志愿者除自由饮水外,不再食用其他任何食物。此外,所有志愿者都被要求在饮食干预期间保持稳定的身体活动水平。在整个过程中特别注意与志愿者保持沟通,了解其依从情况并帮助其随时解决出现的问题。

图1 样品采集时间流程图Fig.1 Illustration of sample collection time series
1.4.3 粪便样品的采集及DNA 的提取
采集志愿者在广西长寿饮食干预前和干预后的新鲜粪便样本,将其立刻放入保温冰盒并尽快转存在-80 ℃冰柜中。按照说明,使用MagPure Stool DNA KF kit B 试剂盒(Magen)提取粪便中细菌的总DNA,并使用Qubit 荧光仪对DNA 定量,用1%(m/V)琼脂糖凝胶电泳检测所提取DNA 的质量。
1.4.4 16S rDNA 文库构建及测序
采用简并PCR 引物341F(5’-ACTCCTACGGGAG GCAGCAG-3’)和806R(5’-GGACTACHVGGGTWT CTAAT-3’)扩增细菌16S rDNA 基因V3-V4 可变区。PCR 反应条件为:94 ℃ 3min;94 ℃ 30 s → 56 ℃ 45 s→ 72 ℃ 45 s,30 个循环;72 ℃ 10 min。PCR 产物用AmpureXP 磁珠纯化。文库经Agilent 2100 bioanalyzer质检合格后,按照Illumina 的标准流程在Illumina MiSeq 平台上测序,产生2×300 bp 的双端序列。
1.4.5 生物信息学分析
对原始数据进行过滤去杂,采用FLASH(v1.2.11)软件将去杂后的双端序列拼接成高质量的Tags,并使用UCHIME(v4.2.40)检测过滤序列中的嵌合体。使用UPARSE 软件(v7.0.1090)将序列相似度大于97%的tags 聚类为操作分类单元(Operational Taxonomic Unit,OTU)。对比Greengenes 数据库(v121305),利用RDP classifier 对每条序列进行物种分类注释,设置最小置信阈值为0.60。使用MOTHUR(v1.31.2)软件在OTU 水平上计算饮食干预前后肠道菌群的α多样性。肠道菌群的β多样性用于比较饮食干预前后的菌群结构差异,以主坐标分析(Principal Coordinate Analysis Biplots,PCoA)表示。此外,基于分类学信息,在门、属等水平上进行饮食干预前后肠道菌群的结构组成和差异性分析。
1.5 数据统计分析
使用SPSS 25.0 软件进行数据统计分析,结果表示为。对于正态分布的数据采用配对样本t 检验,非正态分布的数据采用配对的Wilcoxon 非参数检验,显著性水平为p<0.05。冗余分析(Redundancy Analysis,RDA)用来探究饮食与肠道菌群之间的相关性。
2 结果与讨论
2.1 志愿者饮食干预前后膳食摄入状况
2.1.1 饮食干预前后志愿者膳食结构分析
为充分体现广西长寿饮食干预前后志愿者饮食结构的整体变化,基于志愿者的饮食情况构建了包含所有志愿者及其饮食信息的Heatmap 图(图2)。干预前志愿者的饮食信息来自对其饮食调查问卷的具体数据,形象展示了志愿者的习惯性饮食状况。干预期间志愿者的饮食是在广西长寿饮食模式的指导下,结合志愿者情况个性化设计而得。图2 中纵坐标表示志愿者饮食中平均每日各食物种类摄入量的对数值,横坐标表示广西长寿饮食干预前后的志愿者样本。彩色坐标轴由蓝色向红色过度,表示志愿者平均每日摄入该类食物的数量逐渐递增。由图2 可知,在饮食干预前,入选的志愿者平均每日摄入食物以粥和稀饭、深色蔬菜、水果类、米饭为主(平均摄入量>200 g/d),浅色蔬菜类、无馅类食物、家畜肉类次之(平均摄入量>60 g/d),而对加工肉类、油炸类食物、糕点类食物等摄入量较低(平均摄入量<10 g/d),还发现个别志愿者表现出对饮料类的高剂量摄入(平均摄入量>400 mL/d)。在饮食干预期间,为志愿者设计的饮食所依据的长寿饮食模式多源自广西长寿地区百岁老人饮食喜好,表现出与其较为一致的粥类、深色蔬菜类为主的特点[9](平均摄入量>500 g/d)。同时,饮食中包含较大量的水果类、薯类食物(平均摄入量>180 g/d),对于粗杂粮类食物也有较多的摄入(平均摄入量>75 g/d),而对乳制品、油炸类食物、加工肉类等食物则基本不涉及。谢琪等[13]对广西长寿家庭进行饮食调查后,亦发现长寿老人日常饮食中富含蔬菜,主食相对粗杂,谷薯类食物摄入较多,且基本不摄入乳制品,本研究中所设计饮食富含的食物种类与其对长寿老人的饮食调研结果较为一致。

图2 饮食干预前后志愿者饮食结构热图Fig.2 Heatmap of dietary pattern of volunteers before and after dietary intervention
2.1.2 饮食干预前后志愿者食物摄入量对比分析
为了进一步对比广西长寿饮食与志愿者习惯性饮食期间摄入食物的差别,本研究分析了志愿者在饮食干预前后各类食物的摄入量,结果如图3 所示。在饮食干预期间,志愿者平均摄入蔬菜类(876.79 g/d)、水果类(387.72 g/d)、粗杂粮类(78.48 g/d)、蛋类食物量(65.74 g/d)较志愿者在习惯性饮食期间551.21 g/d、218.88 g/d、33.83 g/d、38.16 g/d 相比显著升高(p<0.05)。乳制品(0 g/d)、海鲜类(12.28 g/d)食物的平均摄入量则显著低于志愿者习惯性饮食期间的摄入量66.51 g/d、33.29 g/d(p<0.05)。同时,高碳水化合物类食物(971.01 g/d)、肉类(68.20 g/d)、豆制品(35.45 g/d)的平均摄入量相比志愿者习惯性饮食期间的平均摄入量1 130.22 g/d、84.63 g/d、52.45 g/d 也较低,然而这种差异并不显著(p>0.05)。通过对比分析,清晰地呈现了广西长寿饮食模式中包含的主要食物种类及摄入量,突出了与志愿者习惯性饮食相比,广西长寿饮食模式的主要特征。同时,与现存的健康饮食,如地中海饮食[14]比较来看,广西长寿饮食模式立足于当地食材,饮食中富含膳食纤维,具有高水平蔬菜、水果、粗杂粮摄入,且摒弃了橄榄油,红酒等食物的摄入,更符合国人饮食习惯,易于依从并推广。

图3 饮食干预前后志愿者各类食物每日平均摄入量比较Fig.3 Daily intakes of various type foods of volunteers before and after dietary intervention
2.2 广西长寿饮食模式对人体肠道菌群的影响
2.2.1 饮食干预前后肠道菌群的物种丰富度和α多样性分析
收集志愿者饮食干预前后的粪便样品共28 份,经MiSeq 平台测序分析后,根据物种数量所测得稀释曲线如图4a 所示。稀释曲线可用来判定测序数据的合理性,由图4a 可知,在本研究中,当测序深度至10 000 时,所有粪便样品的稀释曲线趋于平坦,这表明测序量合理,所得测序结果可靠[15]。同时,使用丰度等级曲线来表现每个样本中肠道微生物的丰度和均匀性(图4b)。其中,曲线的水平跨度越宽,表明样本所含物种相对丰度越高,而曲线的平滑度则可有效反映物种的均匀性[16]。由图4b 可知,饮食干预前后志愿者肠道微生物丰富度和均匀性整体差异较小。

图4 饮食干预前后肠道菌群的稀释曲线和丰富等级曲线Fig.4 Rarefaction curve and OTU rank-abundance curve of gut microbiota before and after dietary intervention
广西长寿饮食模式对志愿者肠道菌群α多样性的影响如表2 所示。α多样性可有效量化比较肠道菌群的丰度和物种多样性。其中,Coverage 作为测序深度指数,与样本覆盖率相关。本研究中所有样本的Coverage均大于0.99,表明测序深度已基本覆盖样本中所有序列。Chao1 指数和Ace 指数可反映肠道菌群的丰度,Shannon 和Simpson 指数可反映肠道菌群的多样性[17]。由表2 可知,饮食干预后志愿者肠道菌群的Chao1 指数、Ace 指数、Shannon 指数和Simpson 指数均未发生显著变化,表明短期的广西长寿饮食干预未显著改变肠道菌群的丰富度和多样性。与本研究结果一致的是,Kohnert 等[18]的研究发现尽管短期的饮食干预改变了健康志愿者肠道菌群的组成,但未改变其肠道菌群的α多样性;Willis 等[19]通过对15 项饮食干预研究荟萃分析,结果表明对健康志愿者进行5 d 至24 周的全植物性食物的饮食干预亦未对α多样性产生影响。这可能是因为成人肠道菌群整体多样性相对稳定,故短期的饮食干预对其α多样性的影响较小。
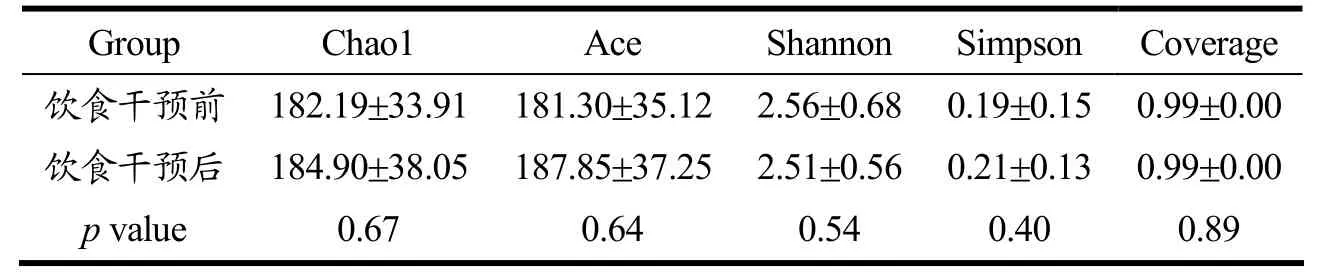
表2 广西长寿饮食干预前后肠道菌群的α多样性分析Table 2 α diversity analysis of gut microbiota before and after dietary intervention
2.2.2 饮食干预前后肠道菌群的β多样性分析
为了进一步区分饮食干预前后志愿者肠道菌群的整体分布状况,对所得数据进行了PCoA 分析。未加权的UniFrac 分析可用于定性测量,通过计算系统发育树中分支长度的比例来比较不同组别的群落距离,所得结果如图5a 所示,两组志愿者的样品在PCoA 图中分布较为一致,未明显分开。加权的UniFrac 分析可在对样品定性测量的同时,考虑每种类型生物的相对丰度,揭示由菌群丰度变化而导致的群落差异。对两组样品再次分析,所得结果如图5b 所示,饮食干预前后志愿者样品在PCoA 图中分布仍较为聚集,β多样性未发生显著改变。这可能是由于饮食干预时间较短所致。毛丙永等[20]用复合益生菌对志愿者进行短期的干预,未改变菌群结构分布;Li 等[21]对志愿者进行三周的蒙古饮食干预后,亦未改变其肠道菌群的PCoA 聚类模式,本研究的结果与其一致。

图5 饮食干预前后志愿者肠道菌群的PCoA 主坐标分析图Fig.5 The PCoA analysis of gut microbiota of volunteers before and after dietary intervention
2.2.3 饮食干预前后肠道菌群门水平差异分析
为了评估广西长寿饮食模式对肠道菌群的具体影响,本研究统计分析了在不同分类水平上肠道微生物优势菌群的相对丰度。志愿者门水平的肠道菌群组成如图6a 所示。其中,厚壁菌门(Firmicutes)、变形菌门(Proteobacteria)、拟杆菌门(Bacteroidetes)和放线菌门(Actinobacteria)为志愿者粪便样品中的主要优势菌群,分别占比56.20%、22.37%、14.70%和6.04%。同时,对比饮食干预前后志愿者肠道菌群门水平结构发现,尽管TM7 在肠道菌群中占比很小,然而,在饮食干预后其相对丰度发生了显著改变(图6b),由0.05%降低至0.01%(p<0.05)。TM7 属于糖化细菌,是一种新发现的候选细菌,在机体内,TM7 已被证明可参与炎症性粘膜疾病的发生,与肠道炎症正相关,且TM7的富集还会导致肠道菌群失调[22]。故饮食干预后TM7丰度的降低可能对志愿者肠道炎症的改善具有积极影响。与本研究一致的是,Choi 等[23]的研究发现短期的水果和蔬菜汁干预可使肠道菌群中TM7 的丰度显著降低,由于广西长寿饮食模式中富含蔬菜和水果,因此,这可能是饮食干预后志愿者肠道菌群中TM7 相对丰度显著降低的原因。
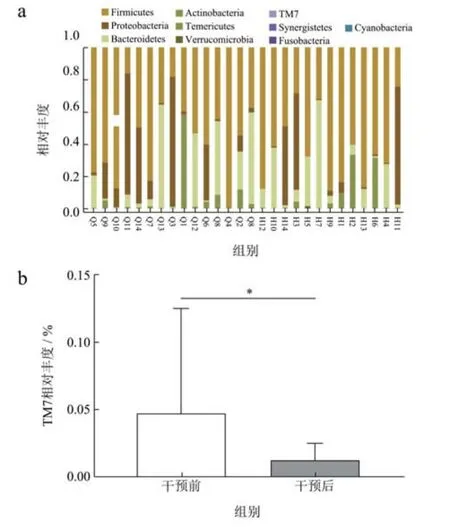
图6 肠道菌群在门水平中物种相对丰度变化及TM7 相对丰度变化Fig.6 Changes of gut microbiota at the phylum level and the relative abundance of TM7
饮食干预后志愿者肠道菌群门水平上的其他菌群组成也发生了变化,如表3 所示。尽管无统计学显著性,相比饮食干预前,14 名志愿者中有8 名志愿者(约占57.14%)粪便样品中变形菌门丰度有所下降。研究表明,变形菌门丰度易受饮食等环境因素影响,其丰度的持续增加会导致肠道菌群的不平衡,且可能与慢性炎症等疾病相关,可作为引发机体疾病的前兆和潜在诊断特征[24]。因此,广西长寿饮食干预后志愿者变形菌门相对丰度在一定程度上的降低,可能对志愿者的肠道菌群结构稳定,机体健康状况的改善有着积极影响。此外,Méndez-Salazar 等[24]的研究表明变形菌门的相对丰度与脂肪摄入量正相关,在广西长寿饮食期间,志愿者肉类摄入较少,且蔬菜、水果摄入量较高,整体脂肪摄入量较习惯性饮食相比较低,故认为变形菌门丰度的降低可能与此有关。De Filippo 等[25]的研究亦表明低脂肪,高膳食纤维的饮食与低丰度的变形菌门水平相关,本研究结果与其一致。此外,由于厚壁菌门和拟杆菌门的丰度在肠道中所占比例很大,因此,厚壁菌门和拟杆菌门丰度的比值(F/B)也可以很好的反映肠道菌群结构在门水平上的变化。尽管在饮食干预后,肠道菌群中F/B 在统计学上无显著差异,然而,由表2 可知,在饮食干预后,14 名志愿者中有9 名(约64.29%)志愿者F/B 值有所降低。与本研究一致的是,Pagliai 等[26]对志愿者进行短期的纯素食干预后,亦未发现饮食对厚壁菌门和拟杆菌门丰度比值造成显著影响,但较饮食干预前相比有所降低。已有报道表明,厚壁菌门与拟杆菌门相对丰度比值的降低与机体能量稳态的改善相关[27],同时,厚壁菌门与拟杆菌门相对丰度比值的增加与机体肥胖,罹患炎症性肠病,便秘等多种疾病有关[28]。因此,饮食干预后,厚壁菌门与拟杆菌门丰度比值的降低在一定程度上表明了广西长寿饮食对肠道菌群结构的调节和对宿主健康状态的改善具有积极作用。此外,Rishi 等[29]的研究表明富含高纤维、高维生素的饮食可以有效降低肠道菌群中厚壁菌门和拟杆菌门丰度的比值,本研究所得结果可能与广西长寿饮食模式中包含此特征相关。

表3 变形菌门、厚壁菌门、拟杆菌门相对丰度及F/BTable 3 Relative abundance of Proteobacteria,Firmicutes,Bacteroidetes and F/B
2.2.4 饮食干预前后肠道菌群属水平差异分析
饮食干预前后志愿者肠道菌群相对丰度较大的前22 种菌属如图7a 所示,箭头表示与饮食干预前相比菌群丰度的变化趋势。其中,18 个属的相对含量大于1%。在饮食干预前,粪便中埃希菌属(Escherichia)所占比例最大(14.00% ),其次为柔嫩梭菌属(Faecalibacterium,7.80%),拟杆菌属(Bacteroides,7.38%),普雷沃菌属(Prevotella,6.78%),芽殖菌属(Gemmiger,6.21%),罗氏菌属(Roseburia,6.07%),瘤胃球菌属(Ruminococcus,5.67%)和双歧杆菌属(Bifidobacterium,5.41%)。
对比饮食干预前后志愿者肠道菌群属水平相对丰度发现,粪球菌属(Coprococcus),瘤胃球菌属,埃希菌属和巨单胞菌属(Megamonas)的相对丰度发生了显著变化(p<0.05)。其中,粪球菌属丰度显著升高(由1.04%增至1.39%),瘤胃球菌属(由5.61%降至2.51%),埃希菌属(由13.41%降至9.02%)和巨单胞菌属(由4.06%降至0.04%)丰度显著降低(图7b~7e)。粪球菌属已被证明对短链脂肪酸的合成有积极作用,且其丰度的增加可抑制体内有害菌的生长,从而缓解肠道炎症,改善肠道菌群失调[30]。研究表明,短期的纯素饮食干预即可增加志愿者粪球菌属水平,而富含肉类的饮食干预则可导致其丰度降低[18]。由此推测,饮食干预后粪球菌属丰度的增高可能与饮食中富含蔬菜相关。瘤胃球菌属丰度已被证实与LDL-c 水平正相关,且其丰度与动脉粥样硬化病变程度显著相关[31]。Gurung 等[32]的研究表明瘤胃球菌属丰度与糖尿病和炎症也呈正相关关系。而高纤维饮食可以通过有效降低肠道菌群中可能的促炎菌属丰度,显示出逆转肠道促炎性生态失调的潜力。此外,瘤胃球菌属丰度的增加也被证明与低水平的水果摄入有关[33]。广西长寿饮食期间,志愿者水果摄入量较高,且饮食中富含膳食纤维,这可能与瘤胃球菌属的降低相关。埃希菌属作为革兰氏阴性菌属,可能会增加血清内毒素水平,从而引起炎症反应。同时,研究表明,细胞壁脂多糖作为内毒素的主要成分,可引起高脂血症和内脏器官中脂肪的积累[34]。因此,饮食干预后埃希菌属丰度的降低可能与机体炎症和血脂水平的改善相关。巨单胞菌属丰度被证明与胆固醇水平正相关[35],在百岁老人肠道菌群中也发现了较低水平的巨单胞菌属丰度[36]。因此,饮食干预后巨单胞菌属丰度的降低可能与改善机体胆固醇水平,降低志愿者心血管疾病风险相关。Pisanu 等[37]的研究发现地中海饮食干预降低了志愿者瘤胃球菌属和巨单胞菌属的丰度,这可能与饮食中纤维和谷物摄入较多,红肉摄入较低有关,本研究与其结果一致。


图7 肠道菌群属水平的变化及差异菌属相对丰度比较Fig.7 Changes of gut microbiota at the genus level and comparison at the relative abundance of difference genus
此外,在饮食干预后,柔嫩梭菌属、拟杆菌属和罗氏菌属丰度也发生了变化,如表4 所示。相比饮食干预前,饮食干预后14名志愿者中有10名(约71.42%)志愿者柔嫩梭菌属丰度有所增加,整体来看,其相对丰度由7.80%增至11.92%,无统计学显著性(p>0.05)。现有研究报道认为,柔嫩梭菌属中包含产生丁酸盐的细菌,且对下调炎症因子、维持结肠健康方面有积极作用[38]。且已有研究表明柔嫩梭菌属丰度与低脂饮食相关[39],这可能是饮食干预后柔嫩梭菌属丰度增加的原因。同时,饮食干预后9 名(约64.29%)志愿者肠道菌群中拟杆菌属有所增加(p>0.05)。研究表明,富含谷物和蔬菜的饮食可通过提供多糖促进拟杆菌属丰度的增加,Koopen 等[40]通过对志愿者进行地中海饮食干预后,亦发现肠道菌群中拟杆菌属丰度有所增加,这与本研究所得结果较为一致。此外,在饮食干预后,14 名志愿者中有8 名志愿者(约57.14%)粪便样品中罗氏菌属丰度有所升高,相比饮食干预前,其平均丰度由6.07%增至7.93%,无统计学显著性(p>0.05)。罗氏菌属可在机体内参与分解糖,产生丁酸,它的增多与高膳食纤维摄入相关[41],Haro 等[42]的研究表明罗氏菌属的丰度升高还与胰岛素敏感性增加相关,可能对预防和治疗糖尿病有一定的作用。综合广西长寿饮食对肠道菌群在属水平上的影响结果分析,表明广西长寿饮食模式对优化肠道菌群结构,降低志愿者心血管疾病风险,调节机体健康状态有重要作用。

表4 柔嫩梭菌属、拟杆菌属、罗氏菌属相对丰度Table 4 Relative abundance ofFaecalibacterium,Bacteroides andRoseburia
2.2.5 肠道菌群多级物种差异性判别分析
为了进一步确定饮食干预对志愿者肠道菌群显著改变的关键菌落,根据线性判别分析效应值(Linear Discriminant Analysis Effect Size,LEfSe)评估了测序信息,设定线性判别分析(Linear Discriminant Analysis,LDA)值为4.0 时,所得结果如图8 所示。与饮食干预后相比,饮食干预前肠球菌科(Enterococcaceae)、肠球菌属(Enterococcus)、埃希菌属、克雷伯氏菌属(Klebsiella)和巨单胞菌属的相对丰度较高(LDA>4.0)。Tian 等[43]的研究表明短期的膳食纤维干预可显著降低肠球菌属的丰度。此外,在纯素食主义的人群中发现了较低水平的埃希菌属[44]。广西长寿饮食中包含较多的水果、蔬菜,较志愿者习惯性饮食相比摄入膳食纤维含量较高,摄入肉类和油脂较少,故饮食中脂肪含量较低,这可能是饮食干预后肠球菌属和埃希菌属降低的原因。克雷伯氏菌属可在肠道环境不佳时大量滋生,其丰度与高碳水化合物的摄入相关,高丰度的克雷伯氏菌属还会引发机体炎症,与细菌性肺炎,脊柱炎密切相关[45]。在广西长寿饮食干预期间,志愿者所食用高碳水化合物类食物较少,故饮食干预后志愿者克雷伯氏菌属丰度的降低应该与此相关,这也表明了广西长寿饮食具有改善机体炎症水平的潜力。Beam 等[46]的研究也证实了高脂肪饮食会导致肠杆菌科,埃希菌属,克雷伯氏菌丰度较高,这与本研究结果一致。巨单胞菌属降低的结论与其在属水平上丰度的显著降低一致,这可能与降低志愿者心血管疾病风险密切相关。

图8 饮食干预前后肠道菌群的LDA 判别图Fig.8 LDA score of gut microbiota before and after dietary intervention
2.2.6 RDA 分析饮食对肠道菌群的影响
为了进一步探究广西长寿饮食对肠道菌群结构的具体影响,选取广西长寿饮食模式中与志愿者习惯性饮食每日平均摄入量具有明显差异的食物种类作为环境变量,应用Envfit 函数来判断食物种类与肠道菌群属水平相对丰度之间的关系(999 次置换循环检验)。所得RDA 结果归结于图9。其中,箭头的长度代表该食物种类的重要性。RDA1 和RDA2 占食物种类与肠道菌群组成之间相关性的49.67%和25.55%。结果表明:肠道菌群的组成变化与蔬菜类食物的每日平均摄入量相关性最高(r2=0.89,p<0.01),其次为水果、牛奶、鸡蛋、粗杂粮(r2>0.60,p<0.01),比较而言,肠道菌群的变化与海产品相关性较差,但仍十分显著(r2=0.59,p<0.01)。结合本研究结果分析,饮食中蔬菜的日摄入量可能对肠道菌群在属水平相对丰度的影响最大。与本研究一致的是,Koponen 等[47]使用同样的方法评估食物成分与肠道菌群关联时,亦发现在所有饮食成分中,富含高纤维的蔬菜是影响肠道菌群最重要的因素。这些结果进一步分析了饮食中对肠道菌群影响密切的食物种类,也为剖析饮食影响肠道菌群的作用机制提供了理论基础。

图9 饮食与肠道菌群关系的RDA 分析Fig.9 RDA analysis of diet and gut microbiota
3 结论
本研究通过分析志愿者在高依从地遵循广西长寿饮食模式干预前后肠道菌群的变化,探究了饮食对肠道微生物的影响及广西长寿饮食模式对人体肠道菌群的调节作用。结果表明:广西长寿饮食模式显著降低了肠道微生物中TM7 的丰度,且促进了粪球菌属的增殖,同时显著抑制了埃希菌属、瘤胃球菌属和巨单胞菌属的生长。LEfSe 多级物种差异判别分析表明,饮食干预后肠球菌科、肠球菌属、埃希菌属、克雷伯氏菌属和巨单胞菌属的相对丰度有所降低。冗余分析表明志愿者每日摄入食物中,蔬菜的日平均摄入量与肠道菌群组成的相关性最高。结合饮食对肠道菌群的影响分析,认为广西长寿饮食干预所引起的肠道菌群变化可能对改善机体炎症、降低心血管疾病风险、维持机体健康状态具有积极作用。这些结果有力地证明了广西长寿饮食模式对人体肠道菌群的有益影响,也为进一步探究通过饮食干预调节肠道菌群,促进机体健康长寿的成效机制提供了理论依据。
猜你喜欢
中国医药科学(2022年14期)2022-08-18
中国典型病例大全(2022年7期)2022-04-22
中老年保健(2021年11期)2021-08-22
昆明医科大学学报(2020年12期)2021-01-26
小学生学习指导(低年级)(2020年9期)2020-11-09
疯狂英语·新读写(2020年1期)2020-04-20
海外星云(2016年23期)2017-01-06
现代妇女(2016年3期)2016-03-31
科学生活(2015年5期)2015-05-19
中国实用医药(2009年13期)2009-07-15
