
二球悬铃木COL5同源基因及其表达模式分析
2023-01-11 06:18孙海莲刘亚红高凤云
北方农业学报 2022年5期
王 洋,谢 宇,孙海莲,石 磊,邱 晓,刘亚红,常 乐,高凤云
(1.内蒙古自治区农牧业科学院 综合试验示范中心,内蒙古 呼和浩特 010031;2.华中农业大学,湖北 武汉 430070;3.内蒙古自治区农牧业科学院特色作物研究所,内蒙古 呼和浩特 010031)
二球悬铃木(Platanus acerifolia)是一球悬铃木(P.occidentalis)和三球悬铃木(P.orientalis)的杂交种,为悬铃木科(Platanaceae)悬铃木属(Plantanus spp.)多年生落叶大乔木,株高可达30 m以上。作为一种观赏植物,二球悬铃木具有生长速度快、分枝能力强、树冠广阔、主干高大等优点,还有吸收硫化氢、二氧化硫、氯气等有毒气体的作用,不仅适合作为行道树,也适合作为厂矿绿化树种[1]。
环境因子能够触发内源信号使其传导,从而调控植物发育阶段的转换。基于一套复杂的分子机制,植物能够准确感应外界环境的变化,从而调整发育策略。对于多年生植物而言,顶端分生组织还受到独特的生长速率调控,会在活跃生长与休眠阶段之间形成周期性地转换,存在“休眠周期”模型[2]。一年生植物中的CO/FT调控模块能够响应日照变化以控制开花时间,同时它也能够控制秋季短日照诱导的休眠和萌芽[3]。
CO是光周期途径中的关键基因,作用是激活下游的FT基因表达,从而诱导植物开花。关于FT基因成花诱导的机制已有研究,但关于FT基因调控休眠的研究还很匮乏。已有相关试验证明,二球悬铃木PaFT基因同时参与了开花和休眠调控,且已经获得了不同休眠时期的转录组数据[4],并结合数据分析选定了CO-Like基因家族中的COL5基因作为目标基因,但是,该基因在不同休眠时期的转录组中存在差异表达,因此有必要明确其对休眠的影响。
FT基因的表达产物被称为成花素。该基因最初克隆自拟南芥,属于小型多基因家族PEBP基因家族,该基因家族的特征在于特定的磷脂酰乙醇胺结合蛋白(phosphatidylethanolamine-binding proteins,PEBPs)结构域,并且该基因家族成员在控制成花转换方面显示出核心作用[5-6]。有研究表明,FT基因的缺失会导致野生型拟南芥无论是在长日照条件下还是在短日照条件下均开花延迟,而FT基因的过表达则导致其不依赖于CO基因或光周期的极端早花,说明FT基因是拟南芥开花时间调控的关键基因。FT基因还具有诱导成花作用并受CO基因的正向调节,是CO基因的下游基因,能够以拮抗方式介导植物的开花信号[7-8]。在合适的光周期调控下,CO将开花信号传导至FT基因从而引起植物叶片中FT表达,进而诱导开花[9-11]。研究表明,FT基因不仅能够在成花转换中起关键作用,植株个体的生理年龄本质可能还与FT基因的表达有关[12],MaFT基因的高表达水平反映出无性繁殖个体成年期发育状态[13]。有研究表明,PaFT4是光周期和热信号的集成器,不仅能够调控植物生长节律,还能够介导芽对光周期和光质量的适应[14]。FT基因除了光周期响应,也能够对温度、光质、热信号等环境因子的变化做出响应[15]。上述研究表明,FT通路可以整合光周期和温度等环境信号从而调控植物的开花及休眠进程。
CONSTANS-Like(COL)基因家族具有保守的B-box和CCT结构域,这已在许多物种中被证明[16]。不同物种的CO及其同源基因作用可能并不同[17-18]。长日照下,拟南芥COL9基因超表达株系出现晚花,COL9突变体出现早花表型,COL3突变体在长日照及短日照下均为早花表型,而COL5基因超表达株系仅在短日照下出现早花表型,说明不同CO-Like基因响应的日照长度不同[19]。CO和COL基因在整个植物生命周期中广泛表达,但并不是所有基因都调控开花或与光周期相关[20]。在拟南芥中,AtCOL基因在调节开花时间、光形态发生、生物钟、根发育、生长素信号传导和花青素积累方面发挥作用。有研究表明,不同物种中的CO或COL同源基因在植物生长和发育中具有不同的功能[21]。因此,研究PaCO-Like基因参与的二球悬铃木休眠调控具有一定的必要性。本试验对COL5基因在单一短日照(24℃,8 h/16 h)、单一低温(8℃,12 h/12 h)和低温短日照(8℃,8 h/16 h)条件下的响应表达模式进行了研究,同时对二球悬铃木成年植株柄下芽不同阶段COL5基因的表达模式进行了分析,旨在明确COL5基因在表达水平上与二球悬铃木休眠调控的相关性。
1 材料和方法
1.1 植物材料
二球悬铃木的柄下芽均采自华中农业大学校园内30~40 a树龄的成熟二球悬铃木,于2020—2021年,分别在休眠诱导期的9月17日、9月29日、10月15日,深度休眠期的11月1日、11月15日、12月15日,次年萌芽期的1月15日、2月15日、3月2日直接采集并置于8 L液氮中保存备用。
二球悬铃木实生小苗群体,分别在单一短日照(24℃,8 h/16 h)、单一低温(8℃,12 h/12 h)和低温短日照(8℃,8 h/16 h)条件下处理,然后分别处理0(对照)、1、4、7、14、21 d后收集叶片并置于液氮中备用。所有样品均进行至少3次独立重复。
1.2 方法
1.2.1 二球悬铃木CO-Like同源基因的筛选
将休眠诱导期(9—10月)、深度休眠期(11—12月)和次年萌芽期(1—3月)的二球悬铃木柄下芽作为样本进行转录组测序,结合基因注释从测序数据库中筛选出表达量有变化的PaCO-Like基因,逐条在NCBI(https://blast.ncbi.nlm.nih.gov/Blast.cgi)上 比对,将与其他物种中COL5同源基因一致性较高的序列作为克隆对象。
1.2.2 COL5同源基因的克隆
根据已知的DNA序列信息,利用Primer Premier 5.0软件设计特异性引物,具体引物见表1,以cDNA为模板进行扩展,克隆得到COL5基因的CDS序列。PCR体系含DNA模板1 μ/L,北京天根有限责任公司生产的10×酶反应缓冲液2 μL、dNTP(10 mmol/L)0.4 μL,北京擎科有限责任公司生产的引物(10 μmol/L)各1 μL,北京天根有限责任公司生产的Taq酶(5 U/L)0.2 μL,加水至20 μL。反应程序为94℃预变性4 min,94℃变性30 s,61℃退火30 s(温度可变),72℃延伸,共35个循环。扩增产物经琼脂糖凝胶电泳分离后,用DNA凝胶回收试剂盒将目的片段回收纯化,连接于克隆载体pMD18-T上,热激转化至大肠杆菌DH5a菌株中。通过菌落PCR鉴定,将阳性克隆基因送上海生工生物技术有限公司测序。

表1 特异性扩增引物
由于转录组数据库中的PaCOL5基因序列不完整,因此本试验采用Tail-PCR(交错式热不对称PCR)进行片段补全。Tail-PCR是根据已知DNA序列,设计3条同向和退火温度较高的特异性引物,兼并退火温度较低的引物(使用MAD1-9和PAD1-2),进行热不对称PCR反应。第1次扩增采用Tail-PCR体系,兼并引物(通引)为MAD1-9。Tail-PCR体系含DNA模板1 μL、10×酶反应缓冲液2 μL、dNTP(10 mmol/L)0.4 μL、兼并引物(通引)2 μL、特异性引物2 μL、Taq酶(5 μ/L)0.2 μL,加水至20 μL。第2次扩增采用Tail-PCR体系,模板为第1次扩增产物,兼并引物(通引)为PAD1。第3次扩增采用Tail-PCR体系,模板为第2次扩增产物稀释20倍,兼并引物(通引)为PAD2。Tail-PCR反应程序及条件见表2。利用琼脂凝胶电泳分离扩增产物,将目的片段用DNA凝胶回收试剂盒回收纯化后,与克隆载体pMD18-T连接,热激转化至大肠杆菌DH5a菌株中,鉴定阳性后送公司测序。

表2 Tail-PCR反应程序及条件
1.2.3 二球悬铃木RNA的提取
二球悬铃木器官总RNA的提取采用蔡芳芳[22]的相关技术体系。
1.2.4 实时荧光定量PCR(quantitative real-time PCR,qRT-PCR)分析
采用实时荧光定量PCR检测COL5同源基因在二球悬铃木中的周期表达模式以及在单一低温、单一短日照和低温短日照条件下的响应表达模式。实时荧光定量PCR反应采用Takara公司生产的荧光定量qRT-PCR试剂盒(SYBR Premix Ex Taq)进行后续试验。利用Invitrogen Primer Premier 5.0软件设计特异性引物,具体引物见表3。提取待测样本的RNA,随后经北京天根有限责任公司生产的DNA水解酶(DNase)处理后反转录成cDNA作为模板,以PaTPI基因为内参基因。实时荧光定量PCR反应体系为10 μL,含cDNA模板1 μL、染料0.2 μL、引物(10 μmol/L)各0.2 μL、酶(5 U/L)5 μL,加水至10 μL。反应过程为95℃2 min;95℃5 s,60℃34 s,40个循环;95℃Melt(溶解曲线)15 s。并用2-ΔΔCt法计算基因相对表达量。

表3 实时荧光定量PCR引物
2 结果与分析
2.1 二球悬铃木COL5同源基因的克隆与序列特征
2.1.1 二球悬铃木CO-Like5同源基因的克隆
根据已知的PaCOL5基因片段分别设计特异性引 物:PaCOL5-1F、PaCOL5-1R和PaCOL5-2F、PaCOL5-2R,以PaCOL5基因表达量较高的二球悬铃木次年萌芽期的DNA为模板,经过测序、拼接和验证,扩增得到2个PaCOL5基因全长,分别命名为PaCOL5-1、PaCOL5-2,再加上前期已经克隆得到的PaCOL5(重新命名为PaCOL5-3)基因作为二球悬铃木COL5同源基因在二球悬铃木中的3个复制(图1)。PaCOL5-1全长为1 444 bp,包括1 134 bp CDS区,推测编码377个氨基酸;PaCOL5-2全长为1 371 bp,包括1 131 bp CDS区,推测编码376个氨基酸;PaCOL5-3全长为1 186 bp,包括1 125 bp CDS区,推测编码374个氨基酸。

图1 二球悬铃木COL5同源基因的PCR扩增结果
2.1.2 二球悬铃木COL5同源基因预测蛋白序列分析
将所克隆的3条PaCOL5基因的预测蛋白序列与其他物种中同源蛋白进行结构分析,结果表明,所有序列中均含有2个B-box和1个CCT结构域,属于典型的COL蛋白(图2)。在以PaFT基因作为组外基因绘制的的系统发育树中,拟南芥COL5基因能够与本试验克隆得到的3个PaCOL5基因同源基因聚在一起(图3)。二球悬铃木COL5同源基因预测蛋白序列与其他植物同源蛋白序列的比对表示该蛋白与荷花(Nelumbo nucifera)、葡萄(Vitis vinifera)中的COL5同源蛋白的亲缘关系很近,暗示它们的功能可能具有保守性。

图2 二球悬铃木COL5同源基因预测蛋白序列与其他植物同源蛋白序列的比对
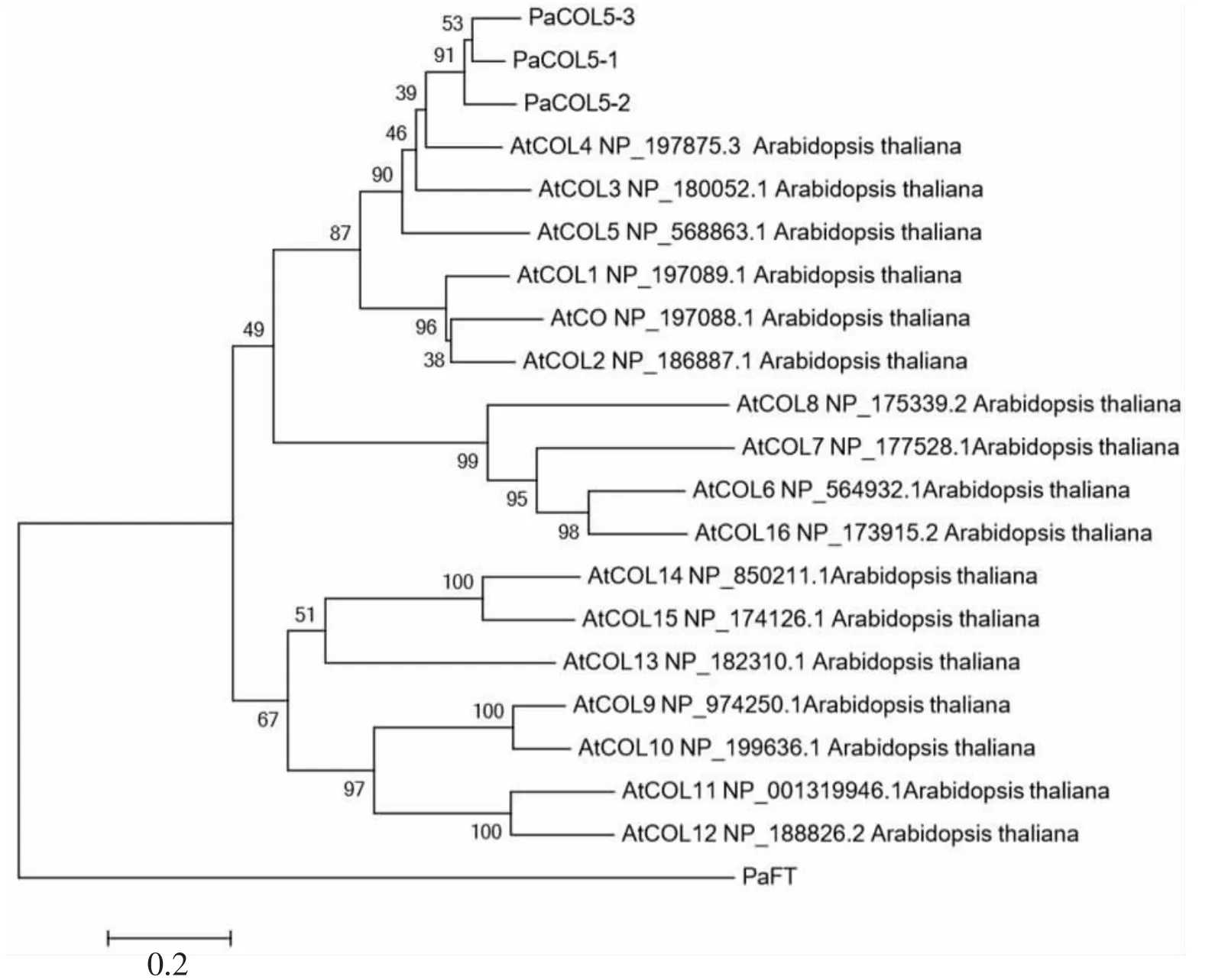
图3 植物COL5同源蛋白的系统发育树
2.2 二球悬铃木COL5同源基因的表达分析
2.2.1 COL5同源基因在二球悬铃木中的周期表达模式
为了研究二球悬铃木COL5同源基因的周期表达模式,在休眠诱导期的9月17日、9月29日、10月15日,深度休眠期的10月31日、11月15日、12月15日,次年萌芽期的1月15日、2月15日、3月2日,采集样品进行了定量实时荧光定量PCR试验。由图4可知,二球悬铃木COL5同源基因PaCOL5-1与PaCOL5-3基因的表达节律较为相近,基因表达前期与PaCOL5-2的趋势相似,都是从休眠诱导期较高的表达量开始下降。但是基因表达中后期有所不同,PaCOL5-1与PaCOL5-3都是从12月开始表达量有所上升,但是PaCOL5-2则是从1月才开始上升。这可能是PaCOL5-1和PaCOL5-3基因与PaCOL5-2基因的功能在基因表达中后期出现了分化所致。

图4 PaCOL5-1(a)、PaCOL5-3(b)和PaCOL5-2(c)基因的周年表达模式分析
2.2.2 PaCOL5在低温和短日照条件下的响应表达模式
由图5可知,在单一低温处理下,仅PaCOL5-1呈现出表达水平下降的趋势;而PaCOL5-2和PaCOL5-3则没有表现出明显的表达规律。此外,由于PaCOL5-3的表达量极低且误差较大,因此不能确定该基因的表达模式。在低温短日照处理下,PaCOL5-1和PaCOL5-3的表达量均呈现先上升后下降的趋势,其表达峰值都出现在处理后7 d,而PaCOL5-2的表达水平则没有发生明显的变化。

图5 PaCOL5-1、PaCOL5-2和PaCOL5-3在单一低温(a)、单一短日照(b)和低温短日照(c)条件下的表达模式分析
3 结论与讨论
序 列 比 对 发 现,PaCOL5-1、PaCOL5-2和PaCOL5-3序列在氨基酸水平上都高度相似。其中,PaCOL5-1和PaCOL5-2序列在氨基酸水平上的相似度为85.1%,PaCOL5-2和PaCOL5-3序列在氨基酸水平上的相似度为84.1%,PaCOL5-1和PaCOL5-3序列在氨基酸水平上的相似度为90.0%。PaCOL5-1、PaCOL5-2和PaCOL5-3拟南芥AtCOL5同源性也较高,达96.1%,但拟南芥COL5基因只有1个单复制,说明二球悬铃木PaCOL5的3个拷贝是在二球悬铃木中独立进化产生的。通过多重序列比对发现,二球悬铃木COL5同源基因与荷花的COL5同源基因序列一致性最高,这与系统进化树分析结果一致。基因序列比对和系统进化的结果均表明本试验3个目标基因属于COL5同源基因(图3)。
实时荧光定量PCR结果发现,PaCOL5呈组成型表达模式,其中次年萌芽期的表达量最高,同时3个基因分别对单一低温、单一短日照和低温短日照产生响应表达。根据周期表达模式分析,PaCOL5-1基因9—10月几乎不表达,10月底到11月表达量开始迅速上升,1月左右达到最高峰,而后开始下降,这与PaCOL5-2基因的周期表达模式刚好相反,所以PaCOL5-2可能是FT基因上游的负调因子。PaCOL5-1和PaCOL5-3基因的表达前期与PaCOL5-2的趋势相似,但是基因表达中后期有所不同,这可能是PaCOL5-1和PaCOL5-3基因与PaCOL5-2基因早期一致,但中后期出现功能分化。PaCOL5-1、PaCOL5-2和PaCOL5-3均可能参与了二球悬铃木休眠调控,但是究竟是否属于FT的直接或间接上游调控因子还需要后期通过蛋白/启动子互作来进一步验证。
本试验中PaCOL5-1和PaCOL5-3均对低温短日照条件有所响应,在处理7 d后会出现1个峰值,因此,可以推测这两个基因可能与休眠环境因子的响应相关。PaCOL5-2对低温短日照条件的响应不明显,分析认为一方面可能是由于处理时间过短,该基因还未产生响应表达;另一方面,也可能是该基因与休眠环境因子响应不相关。从单一低温和单一短日照条件处理中分析得出,单一低温会抑制PaCOL5-1的表达,而单一短日照没有表现出明显的表达规律。综上,PaCOL5-1、PaCOL5-2和PaCOL5-3基因对单一环境因子的响应表达程度不同,可能在二球悬铃木不同的休眠阶段具有协同调控的作用。
猜你喜欢
疯狂英语·新悦读(2022年8期)2022-09-20
基层中医药(2022年4期)2022-07-22
金桥(2021年9期)2021-11-02
金桥(2021年9期)2021-11-02
金桥(2021年9期)2021-11-02
汉字汉语研究(2021年2期)2021-08-30
扬子江(2020年4期)2020-08-04
学苑创造·A版(2020年6期)2020-07-07
汉字汉语研究(2019年2期)2019-08-27
新高考·英语进阶(高二高三)(2018年8期)2018-01-15
