
高羊毛氨酸硒对陕北白绒山羊羯羊生长性能、血液指标、瘤胃发酵、营养物质表观消化率及氮代谢的影响
2023-01-06 07:25朱海鲸李陇平
动物营养学报 2022年12期
李 托 屈 雷 朱海鲸 李陇平*
(1.榆林学院,陕西省陕北绒山羊工程技术研究中心,榆林719000;2.榆林学院,生命科学学院,榆林719000)
硒作为动物生长和生产必需的微量元素之一,具有抗氧化、抗衰老、抗辐射等多种功效。硒不仅可以预防动物某些疾病,还可以促进动物生长发育,对维持动物正常生长发育和生产性能有着极其重要的作用[1-2]。陕北白绒山羊是陕北地区(榆林和延安)主要饲养的绒肉兼用型绒山羊,具有产绒量高和肉品质好等特点,仅榆林地区饲养量就达800万只,绒山羊养殖也已发展成为榆林市畜牧业的主导产业和陕北地区乡村振兴的重要途径。研究表明,陕北白绒山羊饲养区土壤中硒及陕北白绒山羊血液中硒的含量分别为0.069 mg/kg和0.040 mg/L,说明陕北白绒山羊饲养区属于严重缺硒地区,陕北白绒山羊体内硒含量也低于国际上公认的0.05 mg/kg硒含量最低阈值,需要通过饲粮添加剂补充硒元素[3]。饲粮中补充硒能够改善反刍动物瘤胃微生物抗氧化状态,从而促进瘤胃微生物生长和瘤胃发酵,提高营养物质消化率,最终促进动物生长[4-6]。Cui等[7]报道,在饲粮中添加酵母硒能够显著提高藏羊瘤胃氨态氮(NH3-N)、总挥发性脂肪酸(TVFA)、乙酸盐、丁酸盐和丙酸盐浓度。Wei等[8]发现,饲粮中添加硒代蛋氨酸羟基类似物提高了奶牛瘤胃发酵和粗蛋白质(CP)、中性洗涤纤维(NDF)和酸性洗涤纤维(ADF)的表观消化率。也有研究表明,饲粮中添加酵母硒可以提高奶牛的产奶量[9]和绵羊干物质(DM)、有机物(OM)、NDF、ADF的表观消化率和氮代谢[10]。然而,关于硒元素在陕北白绒山羊上的研究鲜见报道,只有熊忙利[3]研究了亚硒酸钠对陕北白绒山羊羔羊断奶成活率的影响和马雄等[11]研究了酵母硒对绒山羊生长和组织抗氧化的影响。高羊毛氨酸硒(selenohomolanthionine,Se-HLan)是以甘油为生长介质,产朊假丝酵母(Candidautilis)进行连续发酵获得的一种新型有机硒[12]。Se-HLan与硒代蛋氨酸(SeMet)在体内的代谢途径不同,SeMet除参与硒蛋白合成外,还有相当一部分被当作蛋氨酸参与正常的蛋白质代谢,而Se-HLan主要被用于硒蛋白的合成。研究表明,水溶性Se-HLan在动物体内具有更高的生物利用率和沉积率,如果以畜禽产品沉积量作为评价硒生物利用率的指标,Se-HLan至少比SeMet高出10%[13-14]。因此,本研究以生长期陕北白绒山羊羯羊为试验对象,研究饲粮添加不同水平Se-HLan对生长期陕北白绒山羊羯羊生长性能、血液常规参数、血清生化和抗氧化指标、瘤胃发酵、营养物质表观消化率和氮代谢的影响,以期为Se-HLan在陕北白绒山羊养殖过程中的应用提供数据支撑。
1 材料与方法
1.1 试验动物
选取体重[(26.18±2.71) kg]相近、健康状况良好的6月龄陕北白绒山羊羯羊32只,随机分为4组,每组8只羊。试验羊每天饲喂2次(08:00和17:00),自由饮水。
1.2 试验设计
1.2.1 饲养试验
参照中华人民共和国农业行业标准《肉羊饲养标准》(NY/T 816—2004)[15]中育肥山羊体重25 kg、日增重100 g时的营养需要配制基础饲粮,加工为颗粒料,基础饲粮组成及营养水平见表1。基础饲粮中硒含量为0.016 mg/kg,分别在基础饲粮中添加0、71、146、296 mg/kg的Se-HLan(硒含量为4 000 mg/kg,载体为麦芽糊精),使饲粮硒含量分别为0.016(对照组,Ⅰ组)、0.300(Ⅱ组)、0.600(Ⅲ组)和1.200 mg/kg(Ⅳ组)。饲养试验包括10 d预试期和60 d正试期。饲养试验结束前1 d,所有试验羊晨饲前通过颈静脉采集5 mL新鲜血液。其中一份血液(肝素钠抗凝血)当天测定血常规参数,另外一份不加抗凝剂的血液3 000 r/min离心10 min制备血清,用于后续血清生化和抗氧化指标的测定。另外,于饲养试验结束当天早上晨饲前,用胃管式瘤胃液采样管经口腔采集所有试验羊瘤胃液20 mL,为了避免动物唾液的污染,收集20 mL瘤胃液之前先弃去刚开始经口腔抽出的50 mL瘤胃液。采集的瘤胃液用4层纱布过滤后,迅速用便携式pH计测定瘤胃液pH后,低温运回实验室转移至-80 ℃超低温冰箱保存备用。

表1 基础饲粮组成及营养水平(干物质基础)
1.2.2 消化试验
饲养试验结束后,将所有试验羊放入自制的消化代谢笼,上层为饲养活动笼,下层为粪尿自动分离装置,饲养活动笼的底部为漏缝地板、前方有料槽和饮水槽、后方设置控位门。按照全收粪、尿法进行为期11 d的消化代谢试验,其中预试期7 d,正试期4 d。消化试验正试期内每天记录每只试验羊的给料量和剩料量,准确计算每1只试验羊的采食量;每天08:00和17:00从代谢笼中的粪便收集板收集每只羊的粪便,同时准确称量粪便重量。混匀后按总重量的10%取样,其中一部分按100 g鲜粪加10%的硫酸10 mL固氮,避免粪中氨氮损失,-20 ℃保存,用于测定粪氮代谢;一部分用于风干样品制备及营养成分的测定。同时,每天用盛有100 mL 10%(v/v)硫酸的塑料盆收集每只羊的尿液,记录每只试验羊的尿液容积后,经4层纱布过滤,按10%比例取样,将收集的每只羊4 d尿样混合到一起,放入-20 ℃冰箱冷冻保存,以备测定尿液中氮含量。所有的羊做完消化代谢试验之后放回原圈。
1.3 指标测定
1.3.1 生长性能指标
饲养试验过程中,每天记录每圈羊的给料量和剩料量并计算平均日采食量(ADFI),试验期间每15 d测定1次所有羊的体重,计算平均日增重(ADG)和料重比(F/G)。
ADG=(终末体重-初始体重)/试验天数;ADFI=[(试验期内每组
羊总给料量-试验期内
每组羊总剩料量)/试验天数]/组内试验羊数;F/G=ADFI/ADG。
1.3.2 血常规指标
用肝素钠抗凝管采集的血液样品,在采用后2 h内使用全自动血细胞分析仪(PE-6800,深圳市普康电子有限公司)及配套试剂测定试验羊的血常规参数,包括红细胞参数、白细胞参数和血小板参数等。
1.3.3 血清生化和抗氧化指标
血清中总蛋白(TP)、白蛋白(ALB)和尿素氮(UN)含量测定使用日本富士全自动干式生化分析仪(4000ie);超氧化物歧化酶(SOD,A001-1)、谷胱甘肽过氧化物酶(GSH-Px,A005-1)活性和丙二醛(MDA,A003-1)含量测定采用南京建成生物工程研究所的试剂盒。
1.3.4 营养物质表观消化率
饲粮和粪样中DM含量参照GB/T 6435—2014中的方法,采用105 ℃烘干法测定;粗脂肪(EE)含量参照GB/T 6433—2006中的方法,采用索式抽提法测定;粗灰分(Ash)的含量参照GB/T 6438—2007中的方法,采用550 ℃灼烧法测定;OM含量为DM和Ash之间的差值计算;粗蛋白质(CP)含量和含氮量根据凯氏定氮法,采用全自动凯氏定氮仪(K9860,济南海能仪器有限公司)测定;将饲粮和粪便烘干、粉碎、压块,采用氧弹量热仪(ZDHW-5微机全自动量热仪,鹤壁华诺电子科技有限公司)测定饲粮及粪中的能量;根据Van Soest等[16]提出的方法测定NDF和ADF含量。
饲粮中某营养物质表观消化率(%)=100×
(该营养物质的摄入量-粪中该营养物质的量)/
该营养物质的摄入量;摄入总能(GEI,MJ/d)=日干物质采食量(kg/d)×
饲粮燃烧值(MJ/kg);粪能(FE,MJ/d)=粪便重量(kg/d)×
粪便燃烧值(MJ/kg);消化能(DE,MJ/d)=GE(MJ/d)-FE(MJ/d);能量表观消化率(%)=100×[DE(MJ/d)/
GE(MJ/d)];食入氮(MJ/d)=日干物质采食量×
饲粮DM含氮量;可消化氮(g/d)=摄入氮-粪氮;总排出氮(g/d)=粪氮+尿氮;沉积氮(g/d)=食入氮-粪氮-尿氮;氮消化率(%)=100×可消化氮/食入氮;氮沉积率(%)=100×沉积氮/食入氮。
1.3.5 瘤胃发酵参数
用Staidson(SG2)手持式pH计测定瘤胃液pH;按照团体标准(T/NAIA 005—2020)中的方法,用GC102AF型气相色谱仪测定瘤胃液挥发性脂肪酸浓度;按照团体标准(T/NAIA 0004—2020)用碱性次氯酸-苯酚分光光度法(722型分光光度计)测定瘤胃液NH3-N浓度。
1.4 数据处理分析
所有试验数据以平均值和均值标准误的形式表示,数据分析采用SPSS 20.0统计软件one-way ANOVA程序进行单因素方差分析。若有显著差异则采用LSD法对组间差异进行多重比较,采用正交多项式对比法确定饲粮Se-HLan添加量的线性和二次效应。P<0.05表示差异显著,P<0.01表示差异极显著。
2 结果与分析
2.1 饲粮Se-HLan添加量对陕北白绒山羊羯羊生长性能的影响
由表2可知,随着Se-HLan添加量的增加,试验羊ADFI呈现出先增加后降低的二次变化(P<0.05),Ⅲ组试验羊ADG、ADFI和终末体重均最高,且Ⅲ组ADG显著高于Ⅰ组(P<0.05),其他各组之间差异不显著(P>0.05);Ⅲ组和Ⅳ组ADFI显著高于Ⅰ组和Ⅱ组(P<0.05),Ⅲ组和Ⅳ组之间以及Ⅰ组和Ⅱ组之间差异不显著(P>0.05)。Ⅲ组试验羊F/G最低,显著低于Ⅰ组(P<0.05)。

表2 不同Se-HLan添加量对陕北白绒山羊羯羊生长性能的影响
2.2 饲粮Se-HLan添加量对陕北白绒山羊羯羊血液常规参数的影响
由表3可知,饲粮添加Se-HLan影响陕北白绒山羊羯羊白细胞计数和嗜酸性粒细胞计数,其中Ⅲ组试验羊嗜酸性粒细胞计数显著高于Ⅰ组和Ⅱ组(P<0.05),其他各组之间较差异不显著(P>0.05),Ⅳ组白细胞计数最高,Ⅲ组次之,Ⅰ组最低,且Ⅲ组和Ⅳ组显著高于Ⅰ组和Ⅱ组(P<0.05)。饲粮添加Se-HLan对其余血常规参数均无显著影响(P>0.05)。
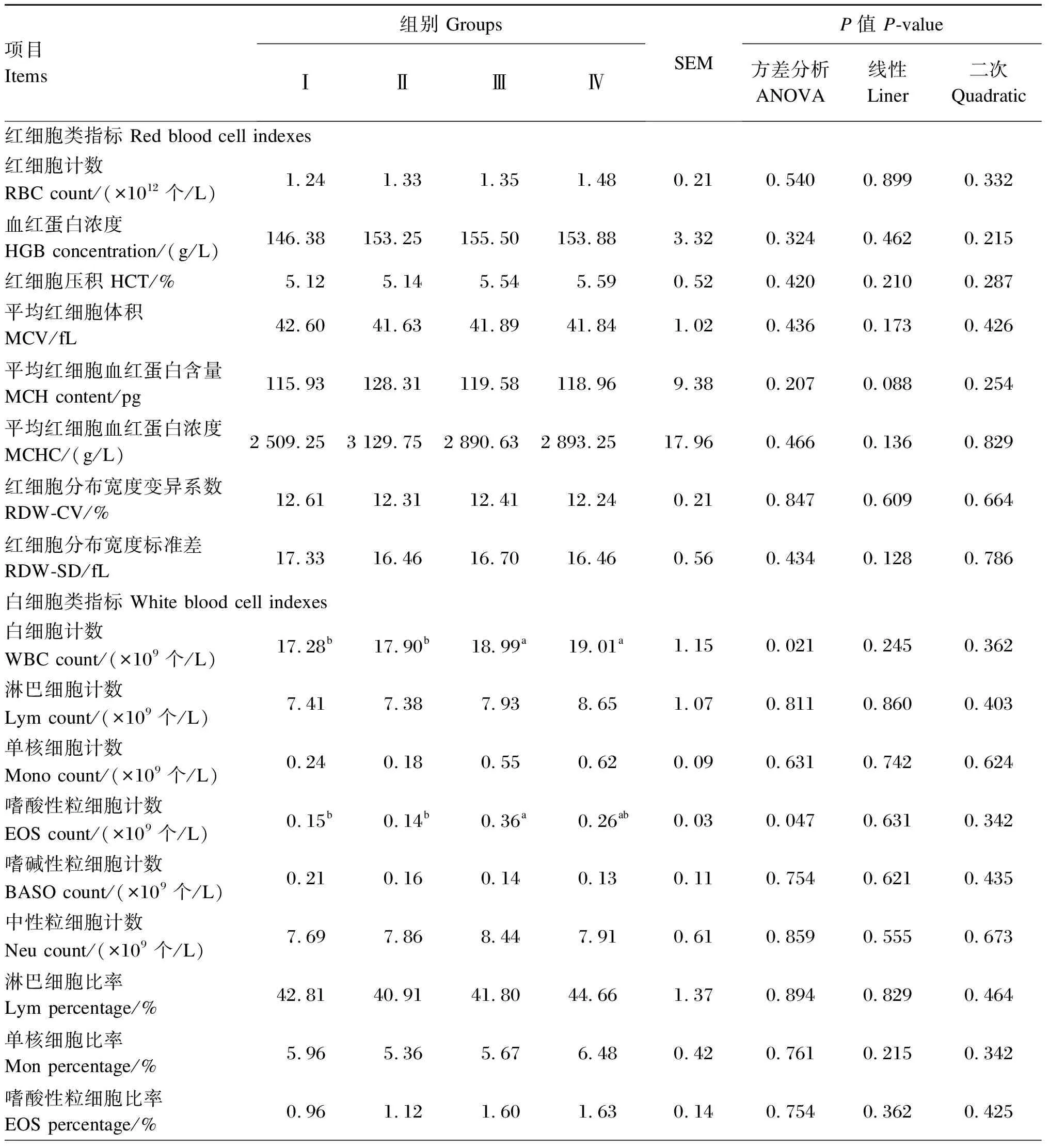
表3 不同Se-HLan添加量对陕北白绒山羊羯羊血液常规参数的影响
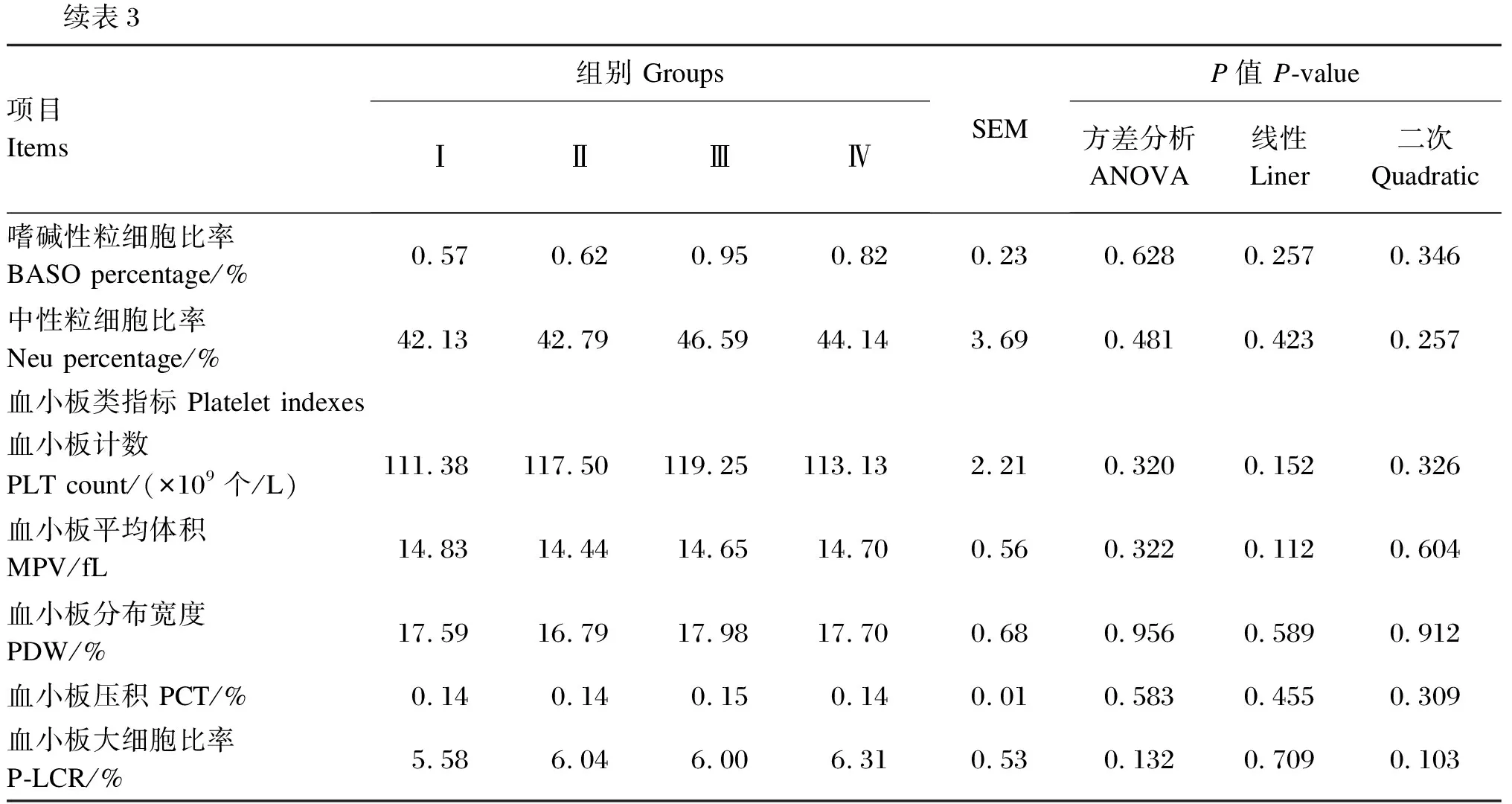
续表3项目 Items组别 GroupsⅠⅡⅢⅣSEMP值 P-value方差分析ANOVA线性Liner二次Quadratic嗜碱性粒细胞比率 BASO percentage/%0.570.620.950.820.230.6280.2570.346中性粒细胞比率Neu percentage/%42.1342.7946.5944.143.690.4810.4230.257血小板类指标 Platelet indexes血小板计数PLT count/(×109个/L)111.38117.50119.25113.132.210.3200.1520.326血小板平均体积MPV/fL14.8314.4414.6514.700.560.3220.1120.604血小板分布宽度PDW/%17.5916.7917.9817.700.680.9560.5890.912血小板压积 PCT/%0.140.140.150.140.010.5830.4550.309血小板大细胞比率P-LCR/%5.586.046.006.310.530.1320.7090.103
2.3 饲粮Se-HLan添加量对陕北白绒山羊羯羊血清生化和抗氧化指标的影响
由表4可知,随着Se-HLan添加量的增加,试验羊血清ALB和TP含量呈现出先升高后降低的二次变化(P<0.01),UN含量则线性降低(P<0.01),Ⅲ组血清ALB含量显著高于I组和Ⅳ组(P<0.05),Ⅱ组和Ⅲ组血清TP含量显著高于Ⅰ组(P<0.05),Ⅲ组和Ⅳ组血清UN含量显著低于Ⅰ组(P<0.05)。
随着Se-HLan添加量的增加,试验羊血清SOD和GSH-Px活性呈现出先升高后降低的二次变化(P<0.01),MDA含量则呈现出先降低后升高的二次变化(P<0.01)。与Ⅰ组相比,Ⅲ组试验羊血清SOD和GSH-Px活性显著升高(P<0.05),其他3组试验羊血清MDA含量显著降低(P<0.05)。Ⅲ组血清SOD活性显著高于其他3组(P<0.05),Ⅲ组血清GSH-Px活性显著高于Ⅳ组和Ⅰ组(P<0.05);Ⅲ组血清MDA含量最低,Ⅰ组最高,且显著高于其他3组(P<0.05)。

表4 不同Se-HLan添加量对陕北白绒山羊羯羊血清生化和抗氧化指标的影响
2.4 饲粮Se-HLan添加量对陕北白绒山羊羯羊瘤胃液发酵参数的影响
由表5可知,添加Se-HLan有增加陕北白绒山羊羯羊瘤胃液pH的趋势,但各组之间差异不显著(P>0.05);随着Se-HLan添加量的增加,瘤胃液NH3-N浓度呈现出先增加后降低的二次变化趋势(P<0.05),Ⅲ组瘤胃液NH3-N浓度最高,极显著高于其他3组(P<0.01);Ⅲ组瘤胃液乙酸浓度最低,显著低于Ⅰ组和Ⅱ组(P<0.05);Ⅲ组瘤胃液丙酸浓度最高,显著高于其他3组(P<0.05);Ⅱ组瘤胃液总挥发性脂肪酸浓度最高,显著高于Ⅰ组和Ⅳ组(P<0.05);瘤胃液丁酸浓度各组之间差异不显著(P>0.05);Ⅲ组和Ⅳ组瘤胃液乙酸/丙酸值显著低于Ⅰ组和Ⅱ组(P<0.05),Ⅰ组和Ⅱ组以及Ⅲ组和Ⅳ组之间差异不显著(P>0.05)。
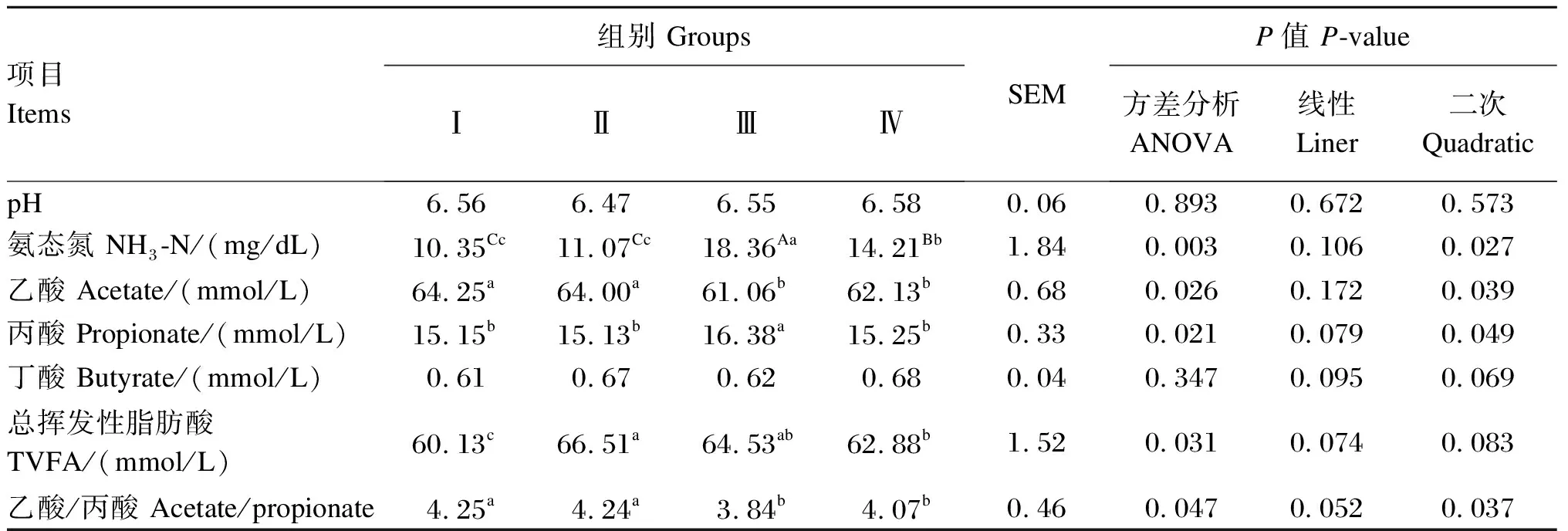
表5 不同Se-HLan添加量对陕北白绒山羊羯羊瘤胃液pH、氨态氮和挥发性脂肪酸浓度的影响
2.5 饲粮Se-HLan添加量对陕北白绒山羊羯羊能量消化的影响
由表6可知,饲粮添加Se-HLan极显著影响陕北白绒山羊羯羊GEI、FE和DE(P<0.01)。随着Se-HLan添加量的增加,试验羊GEI、FE和DE呈现出先升高后降低的二次变化(P<0.01)。其中,Ⅲ组GEI和FE均极显著高于其他3组(P<0.01),其他各组之间差异不显著(P>0.05);Ⅲ组DE极显著高于Ⅰ组和Ⅱ组(P<0.01);Ⅳ组总能表观消化率最高,但各组之间差异不显著(P>0.05)。
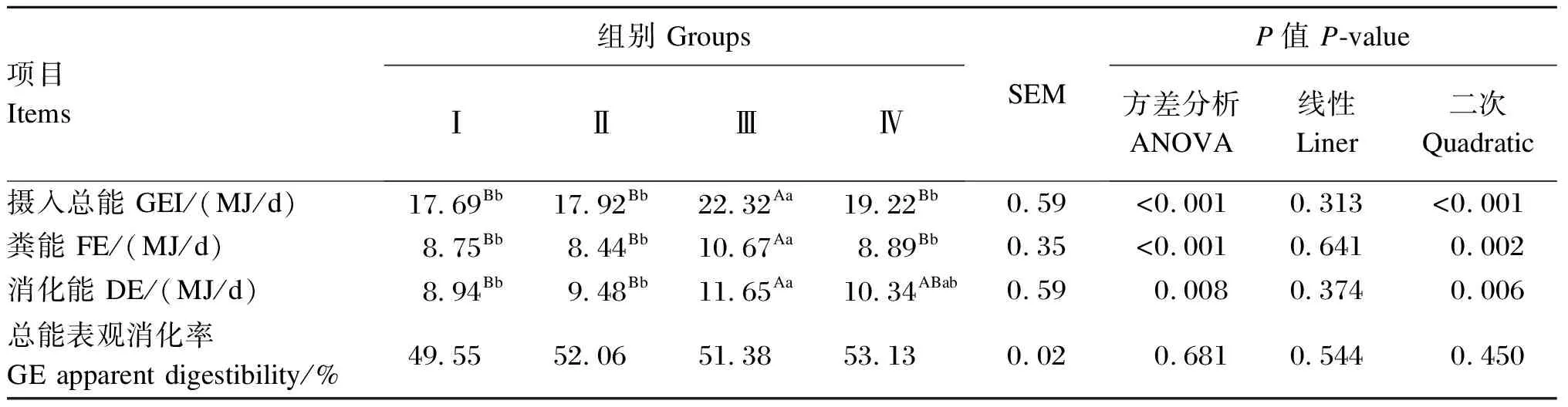
表6 不同Se-HLan添加水平对陕北白绒山羊羯羊能量消化的影响
2.6 饲粮Se-HLan添加量对陕北白绒山羊羯羊营养物质表观消化率的影响
由表7可知,添加Se-HLan显著或极显著影响陕北白绒山羊羯羊DM、OM、NDF、ADF、EE、Ca和P的表观消化率(P<0.05或P<0.01)。随着饲粮Se-HLan添加量增加,DM、OM、NDF、ADF、EE和Ca的表观消化率呈现出逐渐增高的变化趋势(P<0.05或P<0.01);P的表观消化率表现出先增高后降低的二次变化(P<0.05),且Ⅲ组P表观消化率显著高于Ⅰ组(P<0.05)。

表7 不同Se-HLan添加量对陕北白绒山羊羯羊营养物质表观消化率的影响
2.7 饲粮Se-HLan添加量对陕北白绒山羊羯羊氮代谢的影响
由表8可知,Se-HLan添加量极显著影响陕北白绒山羊氮利用及氮利用率各项指标(P<0.01)。随着Se-HLan添加量的增加,食入氮、可消化氮、沉积氮、氮消化率和氮沉积率均呈现出先增高后降低的二次变化(P<0.01)。其中,Ⅲ组食入氮显著高于Ⅰ组(P<0.05),极显著高于Ⅱ组(P<0.01);与Ⅰ组相比,Ⅲ组可消化氮、沉积氮和氮沉积率极显著升高(P<0.01)。

表8 不同Se-HLan添加量对陕北白绒山羊羯羊氮代谢的影响
3 讨 论
3.1 饲粮Se-HLan添加量对陕北白绒山羊羯羊生长性能的影响
关于饲粮添加硒元素对绵羊和山羊生长性能影响的报道中,以前的研究结果不太一致。张永翠等[17]和贾雪婷等[18]的研究发现,饲粮添加酵母硒对杜寒杂交羊(0、0.2、0.4和0.8 mg/kg Se)和滩羊(0、0.25、0.50、1.00和2.00 mg/kg Se)ADG和ADFI均无显著影响;马雄等[11]的研究表明,饲粮添加酵母硒对4~6月龄绒山羊ADG无显著影响。但是,郭孝等[19]研究表明,添加15%高硒苜蓿极显著影响杜泊羊ADG和饲料利用率;郭元晟等[5]的研究表明,添加酵母硒显著提高蒙古羊ADG;张春香等[20]和朱翱翔等[21]研究表明,饲粮中添加亚硒酸钠显著影响山羊生长性能(体重、ADG和饲料转化效率);陈启康等[22]对海门山羊的研究也表明,含硒颗粒饲料极显著影响海门山羊的ADG。本试验结果表明,饲粮中不同Se-HLan添加量显著提高了陕北白绒山羊羯羊ADFI和ADG。本试验研究结果与多数试验结果显示的饲粮添加硒具有提高肉羊生长性能的报道结果一致。硒参与甲状腺激素的代谢,而甲状腺素具有促进生长和发育的功能。饲粮缺硒会导致三碘甲状腺原氨酸(T3)减少和四碘甲状腺原氨酸(T4)增加,降低血液T3/T4的比值,T3减少影响动物生长速率。5-碘甲状腺原氨酸脱碘酶是一种硒依赖性硒蛋白,具有激活T4向T3转化的功能,也是缺硒状态下最后一个活性受到影响的硒蛋白[23]。这表明不同硒蛋白对硒的敏感性存在差异。这在一定程度上解释了补硒并未对动物生长性能(体重、ADG)产生影响的原因。另外,造成硒对动物生长性能研究结果不一致的原因还与饲粮添加不同硒源(亚硒酸钠、酵母硒、蛋氨酸硒和Se-HLan)和不同品种动物(杜寒杂交羊、滩羊和绒山羊)对硒元素的吸收和利用有关[1-2,23]。
3.2 饲粮Se-HLan添加量对陕北白绒山羊羯羊血常规参数和血清生化指标的影响
白细胞参数、红细胞参数和血小板参数构成血常规检测的主要指标,可以反映动物机体的健康状态。白细胞是机体防御卫士,白细胞数量变化是衡量机体免疫力的重要指标[18]。单核细胞、嗜酸性粒细胞和嗜碱性粒细胞参与调节机体局部免疫[24]。本研究结果表明,添加Se-HLan提高了陕北白绒山羊羯羊血液中白细胞计数和嗜酸性粒细胞计数,改善了其免疫状态。血小板计数大于400×109个/L即为血小板增多症,导致皮下出血和血栓病发生[17]。本研究中,试验羊各组间血小板计数无显著差异,表明饲粮添加Se-HLan不会对陕北白绒山羊羯羊机体产生不利影响。血清TP、ALB和UN含量反映了机体蛋白质代谢情况,本试验结果表明,随着饲粮Se-HLan添加量的增加,血清TP和ALB含量均呈现出先增加后降低的二次变化趋势,且Ⅲ组含量显著高于对照组(I组);血清UN含量随着Se-HLan添加量的增加逐渐降低,对照组血清UN含量最高,且显著高于Se-HLan添加组,这些结果与张春香等[20]和鲁玉花等[25]关于山羊的研究结果类似,表明添加Se-HLan降低了陕北白绒山羊羯羊血清UN含量,促进氮素利用,有利于机体蛋白质合成。另外,添加Se-HLan能够显著提高陕北白绒山羊羯羊血清SOD和GSH-Px活性,降低血清MDA含量,张春香等[20]对山羊的研究、黄玉邦等[26]和秦顺义等[27]对羔羊的研究以及石磊等[28]对妊娠母羊的研究也得到了类似的结果,说明添加不同形式硒源均能起到保护动物体内细胞氧化损伤作用,提高山羊和绵羊机体抗氧化能力。本研究中,饲粮Se-HLan的添加量为146 mg/kg (硒含量为0.600 mg/kg),对陕北白绒山羊羯羊血清抗氧化效果最为明显。
3.3 饲粮Se-HLan添加量对陕北白绒山羊羯羊瘤胃健康的影响
瘤胃液pH是反映瘤胃发酵的重要指标,瘤胃pH的高低直接影响瘤胃微生物活力及瘤胃健康状态。最适宜瘤胃微生物生长、繁殖和机体健康的pH为6~7,本研究所有试验羊瘤胃液pH均在此范围里内,表明饲粮添加Se-HLan不会对陕北白绒山羊羯羊瘤胃微生物活动造成不利影响。NH3-N是瘤胃代谢的中间产物,瘤胃液NH3-N浓度变化反映了瘤胃微生物对饲料蛋白质的降解(氮降解)和微生物蛋白合成(氮吸收)之间的平衡状态,是衡量反刍动物对氮利用效率的重要指标。研究表明,瘤胃氨含量过高,瘤胃微生物不能利用的那部分氨就会以NH3-N的形式损失掉;但也有研究表明,瘤胃高水平氨是瘤胃微生物高效利用饲料蛋白质的结果,其和血液蛋白质代谢相关[25]。本试验结果表明,饲粮添加Se-HLan降低了陕北白绒山羊羯羊血清UN含量,增加了血清TP和ALB含量,显著增加了瘤胃液NH3-N浓度,说明添加Se-HLan增强了陕北白绒山羊羯羊瘤胃微生物活力,提高了瘤胃微生物利用NH3-N合成自身蛋白质的能力,增加了体蛋白的沉积,有利用增重。朱翱翔等[21]和Wang等[10]对绵羊的研究,鲁玉花等[25]对山羊研究以及柏建明[29]对湖羊的研究也得出与本试验基本类似的结果。瘤胃微生物对饲料营养物质的消化利用产生大量的挥发性脂肪酸,对于评估瘤胃发酵和瘤胃微生物活力具有重要的参考价值。本研究结果表明,添加Se-HLan提高了陕北白绒山羊羯羊瘤胃液总挥发性脂肪酸和丙酸浓度,降低了瘤胃液乙酸浓度和乙酸/丙酸值,使瘤胃趋于丙酸型发酵类型。反刍动物丙酸型发酵可以产生较多的葡萄糖,可以为机体提供更多的能量物质,有利用育肥,从而提高增重。这与Xun等[30]和Cui等[7]对绵羊的研究得出的结果一致。这说明添加Se-HLan对陕北白绒山羊羯羊瘤胃发酵具有显著的促进作用,也验证了饲粮添加Se-HLan提高了能量表观消化效率。本研究中饲粮Se-HLan的添加量为146 mg/kg (硒含量为0.600 mg/kg),对陕北白绒山羊羯羊瘤胃发酵效果最好。
3.4 饲粮Se-HLan添加量对陕北白绒山羊羯羊营养物质消化率和氮代谢的影响
有关硒对绒山羊饲粮营养物质表观消化率影响的研究较少,但对猪、禽和其他类型的反刍动物的研究较多。研究发现,蛋氨酸硒能够显著或极显著提高猪、禽饲料转化率[31-32]。刘强等[33]对西门塔尔牛的研究表明,饲粮添加蛋氨酸能够显著提高OM、CP、EE、NDF和ADF表观消化率。鲁玉花等[34]的体外试验表明,添加较低水平硒(0.2、0.3和0.5 mg/kg)显著提高纤维素消化率,添加较高水平硒(1.0和2.0 mg/kg)有降低纤维素消化率的作用,而添加高水平硒(5~20 mg/kg)显著降低了纤维素消化率。董升[35]的研究也证实,饲粮添加0.2~0.5 mg/kg的硒能够显著提高纤维降解率。本试验结果发现,添加Se-HLan能够显著或极显莉提高陕北白绒山羊羯羊DM、OM、NDF、ADF、EE、Ca和P的表观消化率。本试验研究还发现,饲粮添加Se-HLan增强了陕北白绒山羊羯羊对饲粮总能的表观消化率。这可能的原因是Se-HLan促进了瘤胃微生物的活性,从而提高了饲粮营养物质的表观消化率。另外,瘤胃绝大多数微生物属于原核生物细菌,细胞膜结构的完整性对于离子交换、电子传递和ATP利用等至关重要,而硒作为GSH-Px重要组分,具有捕获和清除自由基,减少脂质过氧化损伤的作用,还参与辅酶A和辅酶Q生物合成,在三羧酸循环和呼吸链电子传递中发挥重要作用[36]。因此,Se-HLan促进了瘤胃细菌细胞膜结构和功能的完整性,有利于瘤胃发酵、能量代谢等功能的发挥。另外,本试验研究结果表明,添加Se-HLan增加了陕北白绒山羊羯羊食入氮、消化氮和沉积氮,其中0.600 mg/kg Se-HLan组(Ⅲ组)的效果最好。氮是动物生长和育肥期肉羊代谢最重要的营养素,但是关于硒对反刍动物氮代谢的研究报道较少。有报道对植物氮代谢进行研究,结果表明硒可以通过促进硝酸盐还原酶、谷氨酰胺合成酶和谷氨酸合成酶的活性,从而促进氮代谢[37]。Shi等[4]研究了硒对绵羊饲粮消化率和瘤胃发酵的影响,发现硒可以刺激绵羊瘤胃微生物活性和酶活性,进而提高瘤胃发酵和饲料利用率。Wang等[9]对添加酵母硒的藏绵羊进行研究,结果发现添加0.4 mg/kg硒显著提高了NDF、ADF和EE的消化率,并增加了藏绵羊食入氮、消化氮和沉积氮,与本试验结果类似。这些研究表明,硒主要是通过增强瘤胃微生物及其酶活,促进营养物质消化率,进而促进氮的摄入、消化和沉积,但是具体机制有待进一步研究。此外,本研究饲粮营养水平是按照山羊25 kg体重设计,经过饲养试验后开展消化试验的试验羊体重均已达到30 kg,饲粮适宜的营养水平是动物健康生长和生产的重要保障,原有的营养水平在一定程度上可能限制了试验羊对营养物质的表观消化率和氮代谢,试验羊ADG、瘤胃总挥发性脂肪酸浓度均较低。但是,与不加硒的对照组比较,本试验中添加Se-HLan的试验羊均表现出较高的饲粮常规营养物质表观消化率、氮代谢、ADG和瘤胃总挥发性脂肪酸浓度,提示在饲喂营养水平相对较低的饲粮条件下,硒对饲粮营养物质的消化利用和动物健康也发挥了积极的作用,具体机制有待进一步研究和分析。
4 结 论
① 以Se-HLan为补充硒源,饲粮0.600 mg/kg硒含量具有提高生长期陕北白绒山羊羯羊ADFI、ADG、血清抗氧化水平、瘤胃发酵、营养物质表观消化率和氮代谢,促进生长的作用。
② 以Se-HLan为补充硒源,饲粮硒含量在0.300~1.200 mg/kg时不会对生长期陕北白绒山羊羯羊的血液健康和生长产生负面影响。
③ 根据本试验结果,生长期陕北白绒山羊羯羊饲粮Se-HLan的适宜添加量为146 mg/kg (硒含量为0.600 mg/kg)。
猜你喜欢
今日农业(2022年14期)2022-09-15
文萃报·周二版(2022年25期)2022-06-23
北方牧业(2021年16期)2021-11-29
中国畜牧杂志(2021年5期)2021-05-17
国际种业前沿动态(2020年16期)2020-10-14
三农资讯半月报(2020年16期)2020-09-14
读者·校园版(2020年17期)2020-09-02
新农业(2020年5期)2020-06-01
临床检验杂志(电子版)(2020年3期)2020-04-11
湖南饲料(2019年5期)2019-10-15
