
基于转录组分析温度诱导黑斑原鮡(Glyptosternum maculatum)选择性剪接的变化
2023-01-05 04:44王万良周建设扎西拉姆张驰陈美群
水产学杂志 2022年6期
王万良,周建设,2,扎西拉姆,张驰,陈美群
(1.西藏自治区农牧科学院水产科学研究所,西藏 拉萨 850032;2.河南农业大学动物科技学院,河南 郑州 450046)
近些年,全球气候变化[1]导致了一系列水生态问题[2]。西藏是我国重要的生态安全屏障,对全国乃至全球气候的变化影响显著[3],水生生态系统是西藏生态系统的重要组成部分。鱼类是水生低等变温脊椎动物,与环境水体的热交换速率高,容易受到外界水温的影响[4,5]。温度变化可能对它们的行为、生长和繁殖产生负面影响,甚至导致死亡[6]。为了维持这种自我平衡状态,鱼类需要在生理、细胞和分子水平上迅速有效地响应水温的变化。
转录组分析是研究鱼类应对温度变化的有效手段,揭示了多个与氧化应激、蛋白合成、蛋白折叠、细胞凋亡等相关的基因[7,8],尤其是热应激蛋白(Hsps)表达水平在鱼类响应温度变化中显著上调[9]。然而,有关鱼类对温度胁迫后转录后的调控知之甚少,比如转录后的选择性剪接(Alternative Splicing,AS)。选择性剪接在增强真核生物的调节能力和蛋白质机构组成中起关键作用[10],其基本类型包括外显子跳跃(SE)、互斥外显子(MXE)、选择性5’剪接位点(A5SS)、选择性3’剪接位点(A3SS)和内含子保留(RI)[11]。选择性剪接所产生的mRNA 和蛋白质亚型在结构、稳定性、亚细胞定位和功能上可能不同[12]。在发育和对环境压力的反应过程中,选择性剪接是一个被严格调控的过程[13,14],它在应对非生物胁迫,包括热胁迫的过程中发挥着关键作用[7,15]。斑马鱼(Danio rerio)响应热应激后,其热休克蛋白1(hsf1)转录变体的比例发生了明显的变化[16]。

图1 黑斑原鮡背部(a)、胸部(b)及栖息地环境(c)(张驰拍摄)Fig.1 Photos of dorsal(a),chest(b)and habitat of G.maculatum(by Zhang Chi)
1 材料与方法
1.1 实验设计和数据来源
实验中,用三层刺网在夜间布网,凌晨收网采捕多雄藏布支流(N29°27′58.72″,E86°54′36.06″)的黑斑原,用鱼罐车运输到西藏自治区农牧科学院水产科学研究所养殖基地水族箱暂养10 d。箱温度保持在12℃。为了避免性别和年龄对实验结果的影响[21],挑选体长、体质量相近的雌性个体90 尾,分为3 组,每组30 尾。根据黑斑原栖息环境温度的范围[22],试验温度设置3 个组:A 组为对照组,水温保持12℃;B 组以2℃/h 从12℃下降到4℃;C 组以2℃/h 上升,达到24℃时,黑斑原表现张口呼吸,鳃盖猛烈震动,身体颜色变浅,身体出现弓张反应,濒死鱼开始出现。
1.2 样本收集和RNA 提取
选择肝组织作为试验组织[23,24]。C 组在处理过程中,一旦出现濒死鱼,立即取出肝脏,收取3 尾鱼的肝脏样本,然后以2℃/h 降温至12℃保持。同时从A 组和B 组各收集3 尾鱼,用100 mg/L 的MS-222 麻醉致死后取肝脏样本,样本均置于液氮中。总RNA 提取使用TRIzol 试剂(Invitrogen,USA)。采用琼脂糖凝胶电泳和安捷伦2100 生物分析仪(Agilent Technologies,CA,USA)分别测定RNA 浓度和RNA 完整性(RIN)。每个样本3 μg RNA 用于建库,使用NEBNextUltraTMRNA 生成测序文库,使用TruSeq PE Cluster Kit v3-cbo-hs 对样本进行聚类。聚类生成后在Illumina Hiseq 平台进行测序。测序项目由上海派森诺基因科技有限公司服务,RNA-Seq 结果已上传至NCBI 数据库(PRJNA634869)。
1.3 热应激后选择性剪接分析
使用rMATS[25]检测不同样品间的差异剪接基因和样品自身的剪接事件。通过rMATS 统计模型对不同样本进行可变剪接事件的表达定量,然后以likelihood-ratio test 计算P-value 来表示两组样品在Inclusion Level(IncLevel)水平上的差异,lncLevel 并利用Benjamini Hochberg 算法对P-value 进行校正得FDR。FDR<5%为选择性剪接的标准,rMATS 可识别的可变剪接事件有5 种,分别是外显子跳跃(skipped exon,SE)、第一个外显子可变剪接(alternative 5′ splice site,A5SS)、最后一个外显子可变剪接(alternative 3′ splice site,A3SS)、外显子选择性跳跃(mutually exclusive exons,MXE)和内含子滞留(retained intron,RI)。
1.4 功能富集分析
根据GO 注释结果以及官方分类,将差异选择性剪接基因进行功能分类,同时使用R 软件中的phyper 函数进行富集分析。
2 结果与分析
2.1 温度处理后的选择性剪接分析
RNA-Seq 结果,共获得395.87 M的Clean reads,每个样本超过87.29%比对到黑斑原基因组和77.70%比对到外显子区域(表1)。高温组、对照组和低温组选择性剪接事件均只存在MXE 和SE 两种,高温和低温处理后选择性剪接事件均显著减少(图2-a),其中SE 剪接事件在对照组、高温组和低温组分别为4 008、3 336 和3 786 个,MXE 剪接事件在对照组、高温组和低温组分别为207、188 和211个。与控制组相比,高温诱导减少了16.39%选择性剪接事件和16.35%选择性剪接基因,低温诱导减少了5.17%选择性剪接事件和6.21%的选择性剪接基因(图2-b)。

图2 温度胁迫下黑斑原鮡肝脏组织可变剪接事件统计Fig.2 Statistical graph of variable splicing events in the liver tissues of Glyptosternum maculatum exposed to different temperature

表1 RNA-Seq 测序结果Tab.1 Summary of RNA-Seq reads mapping to reference genome
2.2 差异选择性剪接分析
如表2 和图3-a 所示,通过不同处理组间两两比较,对照组与高温组间有3 425 个差异选择性剪接事件和2 400 个差异选择性基因,对照组与低温组间有3 106 个差异选择性剪接事件和2 199 个差异选择性剪接基因,低温组与高温组间有3 679 个差异选择性剪接事件和2 530 个差异选择性剪接基因。高温组有75 个特有基因高表达,低温组有32个特有基因高表达。

表2 差异选择性剪接事件Tab.2 Differential alternative splicing events in the three comparisons
2.3 功能富集分析
为了进一步了解差异剪接事件和基因的功能,对温度胁迫产生的选择性剪接基因进行GO 富集分析。从C vs.H、C vs.L 和L vs.H 的3 个比较组中分别选出309、187 和390 个选择性剪接基因(P<0.01)进行GO 富集分析。其中在C vs.H 组有190 个基因富集到生物学过程(biological process,BP),88 个基因富集到细胞组成(cellular component,CC),318 个基因富集到分子功能(molecular functions,MF);在C vs.L 组,101 个基因富集到生物学过程,50 个基因富集到细胞组成,210 个基因富集到分子功能;在L vs.H 组,富集到生物学功能、细胞组成和分子功能的基因分别是250、109 和406 个(图3-b)。
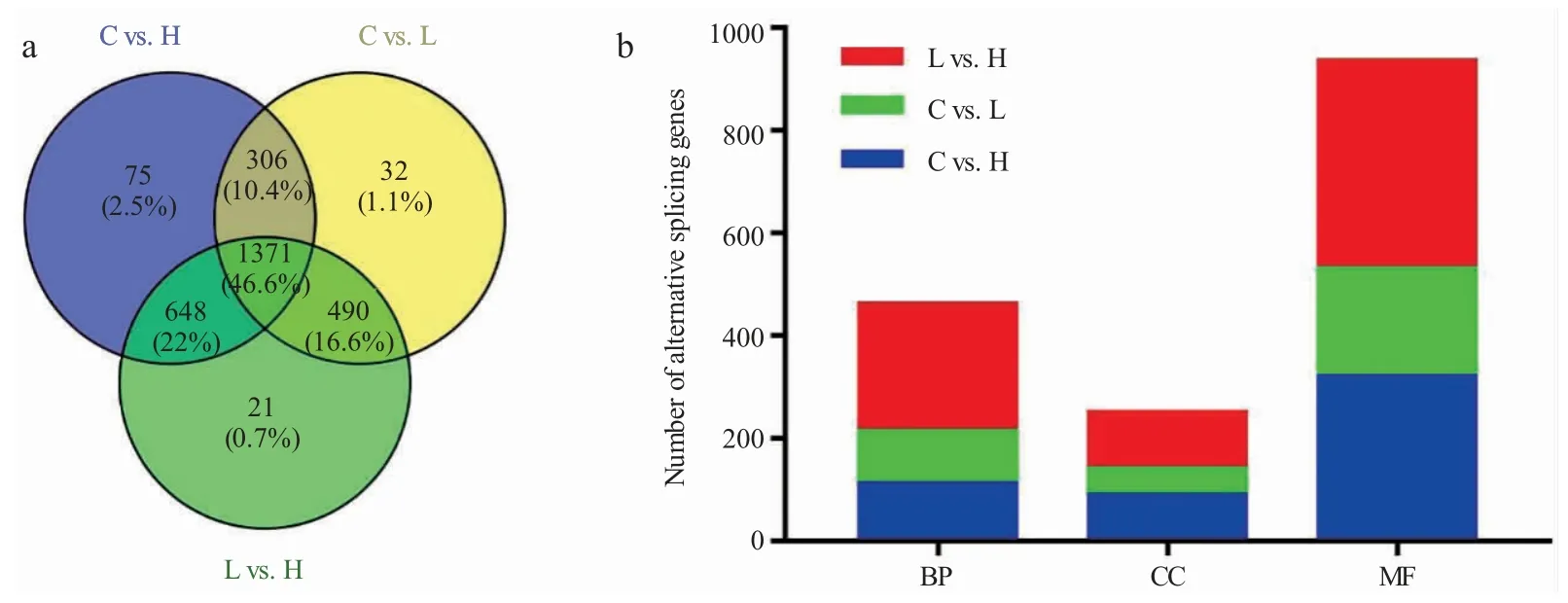
图3 差异选择性剪接基因韦恩图(a)及GO 富集分析(b)Fig.3 The Venn diagram(a)and GO enrichment analysis(b)of differential alternative splicing genes in the three comparisons
KEGG 分析表明,83、9 和88 个通路分别显著富集(P<0.05)在C vs.H 组、C vs.L 组和L vs.H 组,这些通路中,Protein processing in endoplasmic reticulum 和Purine metabolism 在3 个比较组中共有(图4)。2 个共有通路中共有7 个基因(表3),其中热休克蛋白40 同源物亚家族C 成员1(dnajc1)和次黄嘌呤脱氢酶1(impdh1)基因在C vs.H 组差异显著(P<0.05),内质网内降解蛋白2(derl2)、dnajc1、黄嘌呤脱氢酶(xdh)和3′磷酸腺苷5′磷酰硫酸合成酶2(papss2)在C vs.L 组差异显著(P<0.05),xdh、papss2、impdh1、聚合酶(RNA)III(DNA 引导)肽H(polr3h)、dnajc1、derl2 和热休克蛋白40 同源物亚家族C 成员10(dnajc10)在L vs.H 组差异显著(P<0.05)。值得注意的是,dnajc1 在3 个比较组中均差异显著。该基因属hsp40 亚家族成员[26]。

表3 3 个比较组共有通路中基因名称Tab.3 Gene name of co-pathway in the three comparison groups

图4 3 个比较组基因数目前20 个KEGG 通路分析Fig.4 The top 20 KEGG pathways in the three comparison groups
2.4 dnajc1 基因生物信息学分析
人dnajc1 基因选择性剪接转录本如图5-A 所示有12 种,其中选择性剪接体a 为基础转录本,含12 个外显子和11 个内含子,其他转录本均是由a转录本通过外显子跳跃、互斥外显子、选择性5’剪接位点、选择性3’剪接位点和内含子保留的方式衍变而来。本研究中,dnajc1 基因在对照组和低温组鱼肝脏中均产生1 个剪接体。该剪接体在13 号染色体跳过外显子起始和终止位点分别为11110521和11110612、上游外显子起始和终止位点分别为11110338 和11110429、下游外显子起始和终止位点分别为11110761 和11110922。而高温处理后发现了选择性剪接,除了原有剪接体外产生了1 个剪接体。该剪接体在13 号染色体跳过外显子起始和终止位点分别为11109595 和11109642、上游外显子起始和终止位点分别为11109387 和11109489、下游外显子起始和终止位点分别为11109728 和11109894(表4)。生物信息学分析显示,黑斑原肝脏dnajc1 基因选择性剪接转录本如图5-C 所示,与人dnajc1 基因比较,黑斑原选择性剪接体a 为基础转录本,与鲶(Silurus asotus)(图5-B)和人dnajc1基因一样都有12 个外显子,11 个内含子,黑斑原肝脏选择性剪接体b 在本研究的对照组、低温组和高温组出现,而选择性剪接体e 只在高温组出现,也有报道对鲶进行热应激后诱导了热休克蛋白基因hsf1 产生了2 种选择性剪接体[27],进一步说明dnajc1 基因在黑斑原遭遇温度胁迫后肝脏通过产生选择性剪接体来保护细胞的稳态。

表4 不同温度下dnajc1 基因选择性剪接转录体Tab.4 Alternative splicing transcript of dnajc1 gene at different temperatures
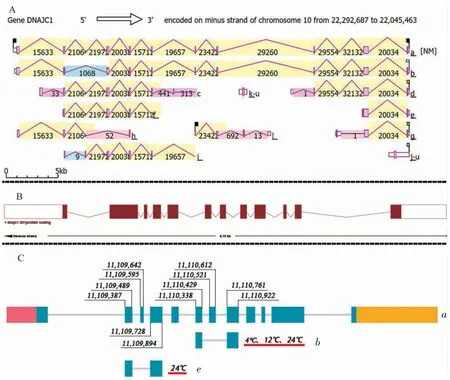
图5 dnajc1 基因选择性剪接示意图Fig.5 Diagram of alternative splicing of dnajc1 gene
3 讨论
3.1 选择性剪接在温度应激中的作用
鱼类是研究环境胁迫下生理和分子反应的良好材料[28]。环境胁迫下生物体通过调控基因的表达模式快速改变转录产物来适应环境[29],环境胁迫因子对基因表达的影响可能是来自于选择性剪接机制的调控[14,30]。然而,选择性剪接事件对鱼类应对环境影响的报道并不多。随着全球气候的逐渐变暖[31],温度胁迫对鱼类的各种影响也在增加。本研究分析了黑斑原在低温和高温处理下肝脏组织的选择性剪接,同时对温度胁迫下选择性剪接基因进行了富集分析,为更好地理解选择性剪接调控机制在应对温度胁迫中的作用提供理论基础。研究表明,选择性剪接有助于动植物更好地适应环境的各种变化[13-15,30]。本研究中,黑斑原经低温和高温处理后,肝脏组织选择性剪接基因和事件数量均减少(图2-b),表明选择性剪接在黑斑原温度胁迫后肝脏组织调解基因表达发挥了作用,这与鲶肝脏[27]热应激后选择性剪接事件和基因均增加相反,选择性剪接在发育和对环境压力的反应过程中受到严格调控[13,14],是增加基因表达复杂性的重要机制,在细胞分化和机体发育中起重要作用[32],温度应激后选择性剪接事件和基因的增加和减少均是对热应激的反应。黑斑原属冷水性鱼类,常年生活在平均海拔4 000 m 以上、水温低于15℃的水域中[19],环境温度的改变致使其减少一些生物功能以维持在高寒环境中生存的需要很有必要。
3.2 热休克蛋白保护机体细胞稳态
Hsp40 蛋白(也称为Dna J 蛋白)是hsp 超级家族中成员最多的亚家族之一,其结构中保守的J 区域为hsp40 与hsp70 结合并调节hsp70 蛋白ATP 酶活性所必需[33]。hsp40 蛋白参与多种细胞功能,包括调节蛋白质折叠、易位和组装,具有多达50 余个同源基因,其中包括dnajc1 基因[26]。本研究中黑斑原经低温和高温处理后,dnajc1 基因在3 个比较组中均差异显著,都富集在内质网内蛋白过程通路中,内质网是一个动态的、复杂的细胞器,在细胞内调节细胞过程,维持细胞内稳态。当这种动态平衡被打破,便出现内质网应激,使内质腔内未折叠的蛋白质不断累积,一种称为未折叠蛋白反应(UPR)信号转导机制启动以恢复内质网的功能和蛋白质平衡[34]。HSP 蛋白在协助内质网内合成蛋白质、保护新生多肽免受各种应激刺激和维持蛋白停滞等方面起着决定性的作用[35]。内质网应激后诱导的选择性剪接可能有重要的生理和病理意义[36]。本研究中,高温处理后黑斑原的dnajc1 基因产生了2 个剪接体,而低温处理后和对照组一样只产生了1 个剪接体(表4)。黑斑原属冷水性鱼类,高温的胁迫作用比低温更明显,因此dnajc1 基因在高温胁迫选择性剪接体发生了变化,以保护多肽免受温度胁迫的应激。大量证据已经证实,Hsp 蛋白的表达对机体的热应激具有保护作用[37,38],但具体保护哪些酶或结构蛋白以及哪些核苷酸的代谢值得研究,Smolenski等[39]通过分析热休克大鼠心脏腺嘌呤、鸟嘌呤、嘧啶和吡啶核苷酸的浓度,证实热应激引起大鼠心脏核苷酸代谢的显著改变,在含氧量正常的条件下,鸟嘌呤和嘧啶核苷酸水平升高,而本研究中温度胁迫下黑斑原肝脏中的4 个基因(表3)均在嘌呤代谢通路中差异显著,进一步推测黑斑原可能在温度胁迫下,dnajc1 基因通过产生选择性的可变剪接来保护嘌呤代谢过程以维持机体细胞的稳态。
猜你喜欢
电子科技大学学报(2022年5期)2022-10-29
当代水产(2022年2期)2022-04-26
中国生殖健康(2020年4期)2021-01-18
当代水产(2020年4期)2020-06-16
当代水产(2020年4期)2020-06-16
当代水产(2020年3期)2020-06-15
第一财经(2019年8期)2019-08-26
中南民族大学学报(自然科学版)(2019年1期)2019-04-04
中国生殖健康(2018年4期)2018-11-06
安徽医科大学学报(2015年9期)2015-12-16
