
流动沙丘先锋植物沙米对风沙流和干旱胁迫的光合生理响应
2023-01-04 13:32姜生秀
西北植物学报 2022年11期
姜生秀,赵 鹏
(甘肃省治沙研究所,甘肃省荒漠化与风沙灾害防治国家重点实验室[培育基地],兰州 730070)
沙漠约占全球陆地总面积的20%,是陆地生态系统环境的重要组成部分,沙漠环境中重要的气候特征是风沙流[1]。在中国干旱半干旱地区,风沙流活动频繁,形成了不同形式、不同程度的风沙灾害,如公路、铁路及农田等被风沙掩埋,同时也威胁到这些地区生物资源的可持续利用与发展[2]。中国重要的沙尘源地之一位于甘肃民勤,该地区地处巴丹吉林沙漠东南缘,降雨量小,蒸发量大,气候干燥[3]。春季是沙尘暴频繁发生的季节,约占全年的47%[4],风沙流严重影响到植物的种子萌发和幼苗定居,进而影响到植物的更新,从而出现衰退的现象[5]。水分短缺也是干旱区限制植物分布与生长发育的重要因素[6],有研究表明,风和风沙流可以间接地加剧植物的干旱胁迫,降低植物的光合速率,使其生长发育受到抑制[7]。张雯等的研究发现,紫斑牡丹在土壤干旱和风沙流交互胁迫下,其净光合速率较对照降低[8],而风沙流和干旱胁迫是如何相互作用影响植物光合作用的,这一问题亟待解决。
沙米(Agriophyllumsquarrosum) 属藜科(Chenopodiaceae)沙蓬属(Agriophyllum)一年生沙生植物,在中国西北、华北各省的沙漠中广泛分布。沙米具有抗干旱和盐碱、耐贫瘠、耐风蚀沙埋等特性,适应能力强,是流动沙丘恢复过程中的主要先锋物种[7]。曲浩等的研究发现,沙米在不同吹风强度和和吹风时间处理下,其净光合速率、气孔导度和蒸腾速率均随着风速的增大和吹风时间的加长而显著降低[9];张翠霞等就库布其沙漠沙米光合日变化特征进行了研究,结果表明沙米蒸腾速率日变化曲线有明显的“午休”现象[10]; 黄刚等的研究结果表明,沙米净光合速率在重度干旱胁迫下较对照降低[11]。目前,有关风沙流和干旱复合胁迫对沙米光合生理影响的研究尚不多见。在沙漠地区,风沙流和干旱是影响植物生长发育的主要因素,搞清风沙流和干旱互作对植物的影响对于防风和固沙植物的选择具有重要的理论和现实意义。因此,本研究以沙米为研究对象,设置水分和风沙流处理的双因素实验,对沙米幼苗光合指标进行了测定,预期从光合生理的角度来阐述沙米对风沙流和干旱的适应能力,为改善和发展风沙区的经济生态建设提供理论依据。
1 材料和方法
1.1 研究区概况
试验在甘肃民勤西沙窝进行,该区地理坐标为E103°51′,N38°38′,海拔1 378 m。年平均气温7.6 ℃,极端低温-30.8 ℃,极端高温40.0 ℃,无霜期175 d;降雨量小,蒸发量大,气候干燥,年均降雨量113.2 mm,年均蒸发量2 604.3 mm,干燥度5.1,最高达18.7,相对湿度47%;全年风沙日可达83 d,多集中在2~5月,年均风速2.5 m·s-1,最大风速为23.0 m·s-1。
1.2 试验设置及指标测定
1.2.1 野外试验于2020年4月布置实验,样地选择在沙米生长密集的流动沙丘上,设置4个浇水/挡风墙复合处理,每处理3个重复。4个处理分别以CK(对照,不浇水不设挡风墙)、SP1(样地1,只设挡风墙不浇水)、SP2(样地2,只浇水不设挡风墙)、SP3(样地3,浇水+挡风墙)表示。其中,处理中浇水量按当地降水量的1 倍计算,直接用塑料软管送水至样地,处理期间每隔3 d浇水1次。挡风墙为四周围有250目的一层纱网,长3 m×宽2 m×高1 m,顶部在有沙尘天气来之前用纱网覆盖,当晴天时打开纱网。于2020年7月选晴朗无云的天气,从每个样地中分别选取3株长势一致的植株,采用 Li-6400 XT 便携式光合仪(LI-COR,USA) 对其叶片的光合参数进行测定,测定选用自然光源,用透明叶室,从6:00~18:00每2 h 测定1 次,每次连续测定3 d,每株测定3片叶子,每个叶片重复记录5组数据,取平均值。测定参数包括净光合速率(Pn) 、蒸腾速率(Tr) 、气孔导度(Gs) 和胞间 CO2浓度(Ci) 等,并根据光合参数计算水分利用效率(WUE),WUE=Pn/Tr。
1.2.2 风洞试验风沙流和干旱胁迫试验在甘肃省治沙研究所风洞实验室进行,该风洞为直流下吹式,轴线风速4~30 m/s连续可调,洞体总长38.9 m,试验段长16 m,横截面为1.2 m×1.2 m。在风洞试验段铺设沙源,所用沙子为腾格里沙漠西南缘的风成沙,铺设厚度为10 cm。实验材料为沙米一年生盆栽实生苗,其中一部分正常供水(75%田间持水量),另一部分减少供水量(30%田间持水量)以造成干旱胁迫。分别将两种水分处理条件下培育的沙米幼苗置于风洞中进行试验。其中,风速设置为0、6、8、10和14 m·s-14个水平,依次表示对照、轻度、中度、重度和超重度风沙流胁迫强度,均持续吹袭20 min,重复3次。以8 m·s-1风速分别吹袭0、15、30、45 min来模拟不同的风沙流胁迫时间。采用 Li-6400 XT 便携式光合仪(LI-COR,USA)对每个处理的沙米叶片在吹后即时进行光合参数的测定,每个水平处理3株植物,每株测定3片叶子,每个叶片重复记录5组数据,取平均值。
1.3 数据处理
采用 Excel对所测数据进行整理分析。所有数据采用 Prisim 5.0软件进行分析,采用单因素方差分析(one-way ANOVA)、Tukey法检验和多重比较,试验数据用平均值±标准误差表示。
2 结果与分析
2.1 浇水和挡风处理对野外沙米光合参数日进程和日均值的影响
首先,图1,A显示,野外试验中沙米叶片净光合速率(Pn)日进程在CK(不浇水不设挡风墙)、SP1(只设挡风墙)和SP2(只浇水)处理下均呈“双峰”曲线,均存在明显的“午休”现象。其中,CK、SP1处理日变化曲线均从6:00开始上升,分别在8:00和14:00出现两个峰值,均在10:00出现明显“午休”,第一个峰值均明显高于第2峰值,SP1处理的峰值较CK增大;SP2处理日变化曲线也分别在8:00和14:00出现峰值(分别为3.35 和2.37 μmol·m-2·s-1),但其在12:00出现“午休”,比CK、SP1处理延迟,且其峰值和谷值均明显高于CK、SP1处理。SP3(浇水+挡风墙)处理沙米叶片Pn日进程曲线比较平缓,呈“单峰”曲线,第 2 峰消失;叶片Pn在上午随着光强增强而升高,在10:00达到最大值(4.30 μmol·m-2·s-1),下午随光强的减弱而降低。同时,从表1可知,沙米叶片Pn日均值以SP3处理最高,并与其余处理差异显著(P<0.05),SP2处理次之,且显著高于SP1处理和CK(P<0.05),而SP1处理与CK之间无显著性差异(P>0.05)。由此可见,相比较CK而言,只增加挡风墙处理对沙米Pn日进程和日均值无明显影响,但使峰值增加;单一浇水处理解除干旱胁迫不仅增加了光合速率峰值和日均值,还使得“午休”时间推后;浇水+挡风墙复合处理使得沙米叶片光合“午休”消失,且峰值和日均值明显增加。

CK.对照,不浇水不设挡风墙;SP1.只设挡风墙;SP2.只浇水;SP3.浇水+挡风墙;下同图1 不同样地沙米叶片光合气体交换参数日变化CK. Contrast, no watering and no windshields; SP1. Windshields; SP2. Watering; SP3. Watering + WindshieldsFig.1 The daily process of photosynthetic gas exchange parameters of A. squarrosum in different sample plots
其次,沙米叶片气孔导度(Gs)日变化趋势与Pn表现出一定的相似性(图1,B)。CK、SP1和SP2处理沙米Gs日进程均呈“双峰”曲线,在日出后开始迅速增加,CK和SP1处理在8:00出现第1峰值(最大值),分别达到0.035和0.04 mol·m-2·s-1,随后开始下降,到10:00出现最小值,然后又开始回升,在12:00出现第2峰值,然后降低并趋于平稳;SP2处理日变化曲线分别在10:00和14:00出现两个峰值,且峰值明显高于CK和SP1处理;SP3处理日变化呈“单峰”曲线,在10:00出现最大值(0.08 mol·m-2·s-1),明显高于其余处理峰值。同时,沙米叶片Gs日均值表现为SP3>SP2>SP1>CK,且前两者均与后两者差异显著(P<0.05),但前两者之间、后两者之间均无显著差异(P>0.05)(表1)。
再次,各处理沙米叶片的胞间二氧化碳浓度(Ci)日变化曲线与Pn和Gs明显不同,但各处理Ci低谷出现的时刻与光合速率达到最大的时刻一致(图1,C)。Ci日进程曲线在CK和SP1处理下均呈不规则的“V”形,均在8:00出现最小值,而SP2和SP3处理均呈不规则的“W”型,在6:00时最大,之后随着光强的增强出现先下降后上升再下降的趋势,均在10:00和14:00出现最小值,在12:00出现峰值,但SP3的峰值小于SP2。同时,各处理沙米叶片Ci日均值表现为:SP1>CK>SP2>SP3,但仅在SP1与 SP3处理有显著差异(P<0.05),其余处理间均无显著性差异(P>0.05)(表1)。
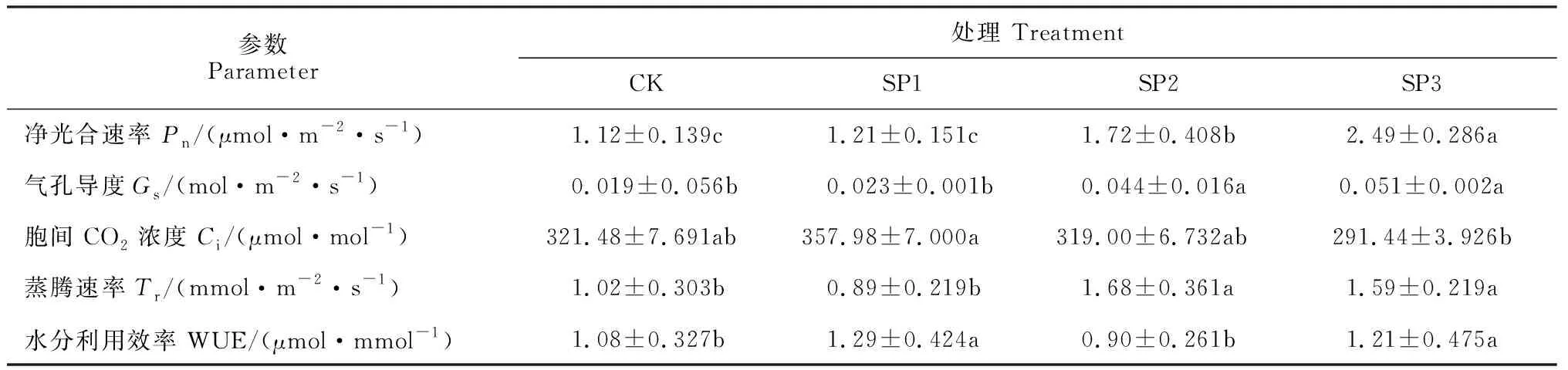
表1 不同处理野外样地沙米叶片光合气体交换参数的日均值比较
此外,各处理沙米叶片蒸腾速率(Tr)日变化曲线呈不规则“抛物线”型(图1,D)。其中,CK和SP1处理最大值出现在8:00,CK峰值大于SP1处理,而SP2和SP3处理最大值出现在10:00,且SP2峰值大于SP3处理;各处理沙米叶片Tr出现最大值的时间也正好是其Gs出现最大值的时间,说明了沙米叶片Gs与Tr的变化有一定的相似性。同时,沙米叶片Tr日均值表现为:SP2>SP3>CK>SP1,且前两者与后两者间差异显著(P<0.05),但前两者之间、后两者之间均无显著差异(P>0.05)(表1)。
水分利用效率(WUE)是植物光合和蒸腾综合作用的结果[11-12]。图1,E显示,CK沙米叶片的WUE日进程为‘双峰’曲线,分别在8:00和12:00出现峰值,在10:00出现最小值;SP1、SP2和SP3处理的WUE日变化趋势与其Pn表现出一致性,SP1和SP2处理WUE在8:00最大,SP3处理WUE则在10:00最大。同时从表1可以看出,沙米叶片WUE日均值表现为:SP1>SP3>CK>SP2,同样表现为前两者与后两者间差异显著(P<0.05),但前两者之间、后两者之间均无显著差异。由此说明,沙米叶片光合性能表现为浇水+挡风墙处理最好,单独浇水处理比设挡风墙处理好,改善水分环境更有利于沙米进行光合作用。
2.2 不同风速的风沙流对沙米叶片光合气体交换参数影响
首先,图2,Ⅰ显示,在浇水条件下,风沙流胁迫的沙米叶片净光合速率(Pn)在6 m·s-1风速下最高(13.51μmol·m-2·s-1),且与无风对照(0 m·s-1)和其余风速处理差异显著(P<0.05),在8 m·s-1风速处理下与无风对照无显著差异(P>0.05),在10和14 m·s-1风速处理下显著低于无风对照(P<0.05)。在干旱条件下,沙米叶片Pn随着风速增加而逐渐降低,但在6 m·s-1风速下与无风对照(10.89 μmol·m-2·s-1)无显著差异(P>0.05),在其余风速处理下均比无风对照显著降低,降幅为43.43%~84.02%。另外,在相同风速处理下,浇水处理沙米的Pn均不同程度地高于相应的干旱胁迫处理。
其次,在浇水和干旱条件下沙米叶片气孔导度(Gs)随风沙流速度的变化趋势与Pn相似(图2,Ⅱ)。但是,各风速处理沙米叶片Gs在浇水条件下均与无风对照无显著差异(P>0.05),在干旱条件下均不同程度低于无风对照,且除6 m·s-1风速处理外降幅均达到显著水平。另外,在相同风速处理下,浇水处理沙米的Gs基本上均不同程度地高于相应的干旱胁迫处理。
再次,沙米叶片胞间二氧化碳浓度(Ci)除浇水条件下14 m·s-1风速处理显著高于无风对照外(P<0.05),浇水条件下其余风速处理和干旱条件下各风速处理均与无风对照无显著差异(P>0.05);在相同风速下,沙米叶片Ci在浇水和干旱环境之间也比较接近(图2,Ⅲ)。
此外,各风速处理沙米叶片蒸腾速率(Tr)在浇水和干旱条件下均不同程度地高于相应无风对照,升幅分别在22.24%~42.37%和10.58%~40.76%,且有随着风速增加而上升的趋势,但其中只有浇水条件下14 和10 m·s-1风速处理的Tr与相应无风对照差异显著(P<0.05);在相同风速条件下,沙米叶片Tr表现为浇水处理均不同程度高于相应干旱胁迫处理(图2,Ⅳ)。

图2 不同风速风沙流胁迫下沙米光合气体交换参数比较Fig.2 The comparison of photosynthetic gas exchange parameters of A. squarrosum under different wind speed treatments
最后,沙米叶片水分利用效率(WUE)在浇水和干旱条件下均随风速的增大而逐渐减小,且除低风速(6 m·s-1)的风沙流胁迫外,其余风速处理均比相应对照显著降低;在相同风速下,沙米叶片WUE在浇水处理和干旱处理间接近(图2,Ⅴ)。
由此可见,在浇水和干旱条件下,沙米叶片的Pn、Gs和WUE均随着风速的增大而逐渐降低,而其Tr则有逐渐升高的趋势,但其Ci则保持相对稳定;浇水沙米叶片的光合作用在轻度风沙流(6 m·s-1)胁迫下受到显著促进,在重度和超重度风沙流(10~14 m·s-1)胁迫下受到显著抑制,而干旱胁迫沙米叶片光合作用在中度至超重度风沙流(8~14 m·s-1)胁迫下受到显著抑制,且受到的抑制程度比浇水处理更重,说明中度以上风沙流胁迫显著抑制了沙米光合作用的进行,而干旱胁迫加重了风沙流胁迫的抑制程度。
2.3 风沙流持续时间对沙米叶片光合气体交换参数的影响
图3显示,浇水沙米叶片的Pn在中度风沙流(8 m·s-1)胁迫15~45 min后与对照之间均无显著性差异(P>0.05),但胁迫45 min处理显著小于胁迫15 min处理(P<0.05);干旱沙米叶片的Pn在中度风沙流胁迫15 min时无显著变化,在胁迫30和45 min后比对照显著降低;在风沙流胁迫30和45 min后干旱处理明显低于相应的浇水处理。浇水和干旱沙米叶片的Gs在中度风沙流胁迫下的变化趋势相似,其叶片Gs均在吹袭15和30 min后与相应对照之间无显著性差异,但均在吹袭45 min后显著低于相应对照;在相同胁迫时间下,干旱处理均不同程度地低于浇水处理,且胁迫时间越长差异越大。浇水沙米叶片Ci在不同风沙流胁迫时间之间均无显著性差异,干旱沙米叶片Ci仅在风沙流胁迫30 min时显著低于对照,在其余胁迫时间也无显著变化。浇水沙米叶片Tr在风沙流胁迫后均明显高于对照,且在胁迫15和45 min后于CK差异显著;干旱胁迫沙米叶片Tr仅在风沙流吹袭45 min时显著高于对照,其余胁迫时间与对照无显著性差异。

图3 不同风沙流持续时间下沙米光合气体交换参数比较Fig.3 The comparison of photosynthetic gas exchange parameters of A. squarrosum under different wind time points
浇水和干旱沙米叶片WUE在风沙流胁迫后均不同程度地低于相应对照,且胁迫时间越长降幅越大,但浇水处理在胁迫15 min降幅已达显著水平,干旱处理却在胁迫30 min时达显著水平。可见,中度风沙流(8 m·s-1)在浇水条件下吹袭45 min才对沙米光合作用产生显著抑制作用,而在干旱胁迫下吹袭30 min时就显著抑制沙米光合作用。说明浇水可以有效延长沙米光合作用对持续风沙流胁迫的耐受时间。
3 讨论与结论
3.1 自然环境下沙米叶片光合气体交换参数对干旱和风沙胁迫的响应特征
3.1.1 参数日进程植物要使其光合机构功能活性达到最大,必须要适应外界光照条件的变化[13]。当外界各种环境因子与植物内部生理调节达到最优时,植物光合日变化就会出现峰值[14]。光合作用是一个易受环境影响的生理过程,由于环境因子的周期性变化,许多植物的光合日变化出现了“午休”现象[15]。据 Farquhar 等的观点,如果植物光合参数Pn和Ci变化方向相同,而Gs降低,则Pn的降低是由气孔因素造成的;如果Pn和Ci变化方向相反,而Gs增大,则Pn的下降主要是由非气孔因素限制,即是由叶肉细胞同化能力降低引起的[16];另外,气孔和非气孔因素在一些情况下可以同时作用[17]。本研究发现,沙米Gs和Pn的日变化在自然条件下均呈“双峰”型。上午随着光强的增加,气孔开放,Gs上升,引起Pn和Tr的迅速上升;之后,随着光照继续增强,气温上升,叶片部分出现短暂的水分亏缺,使得部分气孔关闭,Gs和Pn下降比较明显,Tr也开始下降,在10:00出现“午休”;在12:00 以后,随着光照减弱,气孔开张度增加,Gs和Pn开始升高,Tr也随之上升。这一结果与喀斯特石漠化地区白刺花[18]和库布其沙漠植物[10]的相关研究结果一致。但沙米Ci与Pn、Gs日进程规律相反,经过夜间的富集,Ci在清晨较高,上午随着光照增强,气温和叶温的升高,Pn也逐渐升高,引起Ci降低,在8:00降至最低,而Pn却在这个时段达到最大值。由此说明,自然环境下沙米光合“午休”现象是气孔因素和非气孔因素共同作用的结果。
同时,在本研究中,设挡风墙后(SP1处理)沙米Pn和Gs的“双峰”型日变化规律无变化,但使峰值增加;浇水(SP2处理)解除干旱胁迫不仅增加了Pn,还使得Pn日变化峰值推后;浇水+挡风墙处理(SP3处理)使得“午休”消失,且峰值明显增加。这就说明干旱胁迫会引起光合抑制,沙米为了适应水分亏缺,其叶片气孔选择关闭或暂时关闭,使得光合速率降低,而良好的水分条件则有利于促进光合作用进行。赵哈林等[19]的研究结果表明,频繁净风吹击会增加光合蒸腾“午休”时间,加大“休眠”程度,中午光照强烈、温度过高及湿度过低会对植株造成伤害,植株形成“午休”这种机制来保护自身,在浇水+挡风墙处理后,逆境得以缓解,从而使得“午休”消失。
3.1.2 参数平均值在自然生态系统中,风是一种非常重要的生态因子,影响着植物的生长发育[19]。在本研究中,SP2(浇水)与SP3(浇水+挡风墙)相比,CK与SP1(挡风墙)相比,沙米生长均属于风沙胁迫;SP2处理沙米的日均Pn、Gs和WUE小于SP3处理,而其日均Ci、Tr均大于SP3处理,CK和SP1也表现出相似的趋势。这就说明风吹袭使得沙米叶片气孔关闭,Gs和Pn下降,Ci增大,风吹袭同时促进蒸腾,因为风能将气孔外边的水蒸气吹走,一些相对湿度较低的空气被补充进来,扩散层变薄或消失,外部扩散阻力减小,蒸腾就加快[2]。与此同时,随着风沙流胁迫的加深,致使沙米叶片Pn减小,Tr增大,结果导致WUE减小。说明风和风沙流加重了干旱对植物光合作用的抑制程度。这一结果与于云江[2]及王家强[20]等的相关研究结果一致。
同时,在本研究中,SP1处理与SP3相比,CK与SP2处理相比,沙米生长均受到干旱胁迫,致使SP1处理叶片的日均Pn、Gs和Tr小于SP3处理,而其叶片日均WUE大于SP3处理,CK和SP2处理也表现出相似的变化。这就说明在干旱胁迫条件下,沙米为了适应水分亏缺,其叶片气孔选择关闭或暂时关闭,导致气孔呼吸微弱,Tr和Pn下降。在干旱条件下,水分利用效率越高,则表明植物节水能力越强,耐旱生产力越高说明蒸腾速率对气孔有较强的依赖性,而光合速率除受气孔限制外,非气孔因素也起着相当重要的作用[21]。植物为维持水分平衡,在选择优先光合和优先蒸腾之间达到适度的调节[11]。在干旱条件下,由于水分是最重要的限制因子,选择大幅降低蒸腾速率,而为了提供能量,光合速率仍然维持在适当的水平,因而水分利用效率有所提高。这一结果与于文颖[22]及黄刚[11]等的相关研究结果一致。
另外,在各处理间相比较,沙米叶片日均Pn、Gs以SP3处理最高,SP2次之,CK最小,且SP3处理显著高于其他处理,而SP2、SP1处理与CK之间无显著性差异。这是因为在民勤荒漠区,大风日较多,风力强且持续时间长,在没有人工补水条件下,挡风墙并不能完全阻挡风沙对沙米叶片造成损伤;即使在不设挡风墙时补充了水分,水分也会因强风和日照而迅速蒸发,只有在既浇水又设挡风墙条件下,沙米光合能力才会增强。
3.2 沙米光合气体交换参数对不同风速风沙流和干旱胁迫的响应特征
风速的大小在自然生态系统受风沙流影响强度中起到决定性的作用[23]。已有研究证明,不同风速的风沙流对植物的影响具有两面性,即较低的风速对植物与土壤及大气间物质和能量的交换起到促进作用,进而对植物生长有正向作用,但当风速太大,植物不能承受风沙带来的危害,生长会遭到抑制[24]。本研究发现,低风速(6 m·s-1)风沙流在浇水条件下使沙米叶片的Pn、Gs、Ci和Tr比CK显著升高,但在干旱条件下使得沙米Pn、Gs比CK降低,这就说明,低风速(6 m·s-1)只有在解除干旱胁迫时才促进植物光合作用。之后,随着风速的增大,浇水和干旱胁迫下沙米叶片的Pn、Gs和WUE均逐渐减小,而Ci和Tr均逐渐增大,这是因为无论是浇水还是干旱条件下,不同风速处理相比CK而言,均只是风沙胁迫,因此风吹袭促进了蒸腾作用,使得气孔关闭。植物受到风沙流吹袭时,沙子直接打击到植物体上,植物外部形态和生理结构都会受到损伤,使植物的光合性能降低,无风或轻风不利于植物的蒸腾和光合作用,而和风与劲风则能促进植物的蒸腾和同化作用,但强风的持续吹袭又会对植物造成危害,导致使其光合性能降低,蒸腾作用迅速升高[25]。
3.3 沙米光合气体交换参数对风沙流胁迫持续时间的响应特征
当风吹袭时间较短时,植物通过调节叶片气孔的开闭来调控蒸腾作用,吹袭时间越长,对植物外部形态和生理结构造成的损伤越大,不同植物光合性能对风沙流胁迫的响应方式存在着差异[26]。风速大、吹风间隔长的风沙流使植物光合速率降低幅度最大。已有研究表明,小叶锦鸡儿(Caraganamicrophylla) 等沙生植物在短期风和劲风作用下其蒸腾速率和净光合速率增加[27];梭梭(Haloxylonammodendron)同化枝在风沙流(14 m·s-1)作用 5 min 后气孔导度和蒸腾速率基本不变,而胞间 CO2浓度有所增加[28];油蒿(Artemisiaordosica)等植物幼苗的净光合速率与气孔导度随吹风时间的延长而显著下降[2]。本研究发现,在浇水条件下,沙米叶片净光合速率、气孔导度及胞间 CO2浓度在中风速(8 m·s-1)作用15 min和30 min后与对照之间无显著性差异,而其蒸腾速率显著增加;在中风速作用45 min后,沙米叶片Pn显著小于对照。在干旱胁迫条件下,沙米叶片Pn在中风速作用15min时与对照之间无显著性差异,在作用15 min和30 min后显著小于对照,而其Tr随吹风时间的延长而逐渐增加。由此说明,浇水使得沙米应对风沙流胁迫的能力增强。
猜你喜欢
航空材料学报(2022年4期)2022-08-04
贵州农业科学(2022年4期)2022-05-06
干旱地区农业研究(2022年1期)2022-01-28
当代工人(2020年11期)2020-07-06
汉语世界(The World of Chinese)(2019年5期)2019-11-11
速读·中旬(2019年3期)2019-04-12
科技创新与品牌(2019年12期)2019-02-06
东坡赤壁诗词(2018年6期)2018-12-22
少年文艺·我爱写作文(2017年11期)2017-11-16
作文大王·中高年级(2016年11期)2016-12-09
