
稀子蕨胞芽发育及营养器官的形态与解剖学研究
2023-01-04 13:32王雨,罗睿
西北植物学报 2022年11期
王 雨,罗 睿
(1 贵州大学 生命科学学院,贵阳 550025;2 贵州大学 生命科学学院 山地植物资源保护与种质创新教育部重点实验室,贵阳 550025)
珠芽(bulbil)是半夏(Pinelliaternata)、卷丹百合(Liliumlancifolium)等被子植物的无性繁殖器官,呈球状、卵圆状等形状,常着生于叶柄基部、叶腋或花序上,一般由特定部位的分化体细胞通过脱分化形成起始细胞、原基形成、器官分化、生长和成熟等发育过程形成。珠芽生殖是植物无性克隆繁殖的一种方式,是植物生态适应的一种重要策略[1]。被子植物珠芽生殖的相关研究较多集中于生殖过程的形态、解剖学特征探究,如薯蓣科的薯蓣(Dioscoreaopposita),天南星科的弥勒魔芋(Amorphophallusmuelleri)、半夏和滴水珠(P.cordata),苦苣苔科植物,以及百合科的淡黄花百合(L.sulphureum)和卷丹百合(L.lancifolium)等的珠芽发育形态解剖学特征[2-8]。顶芽狗脊(Woodwardiaunigemmata)、芽胞蹄盖蕨(Athyriumclarkei)和稀子蕨(Monachosorumhenryi)等蕨类植物在植株羽叶叶轴上形成的胞芽(gemmae),是类似于被子植物珠芽的无性繁殖器官。但是,与被子植物的珠芽生殖相比,蕨类植物胞芽生殖的发育生物学研究还非常少见,目前仅见长柄石杉(Huperziajavanica)的胞芽形态结构观察[9]与顶芽狗脊的胞芽形态发育过程和解剖学研究[10-11]。
稀子蕨是中国分布的碗蕨科(Dennstaedtiaceae)稀子蕨属3种植物之一,零星分布于贵州、云南、广西、广东、湖南等地,在贵州已发现分布于都匀市斗篷山、铜仁市梵净山,国外在日本、印度尼西亚、马来群岛、印度及缅甸也有分布[12]。与尾叶稀子蕨(M.flagellare)和穴子蕨(M.maximowiczii)相比,前两个物种常在羽叶顶端接触地表时有顶芽(buds at apex)发生并发育成新的植株,而稀子蕨常在叶轴中部发育一枚或多枚胞芽生长于腋间,胞芽在成熟后从母体植株脱落并在新的生长季节萌发形成新的植株。虽然稀子蕨含稀子蕨素、稀子蕨内脂等化学成分,具有祛风活血作用,可用于风湿骨痛的治疗[13],但是稀子蕨相关的研究目前仅限于分类学的探讨[14]。稀子蕨生殖生物学相关研究的缺乏限制了对其进一步的了解和开发。
鉴于目前尚缺乏对稀子蕨胞芽无性生殖的了解,本研究拟通过形态学、解剖学方法探究其胞芽生殖的发育过程,揭示其形态、解剖学特征。阐明稀子蕨胞芽发育的形态、解剖学特征可为进一步揭示不同蕨类植物胞芽生殖在发育生物学上的异同、分析胞芽发育的分子机制并探讨蕨类植物胞芽发育与被子植物珠芽发育的异同等研究奠定基础。
1 材料和方法
1.1 材 料
稀子蕨(Monachosorumhenryi)植株及相关部位材料采集自贵州省都匀市斗篷山(N 26°21′11″, E 107°23′56″, 图版Ⅰ,A)。解剖学研究材料(具有不同发育时期胞芽的叶轴对应部位和不同位置的营养器官)在野外现场进行取材、FAA固定液固定后保存在实验室备用;活体材料(完整植株、植株上成熟胞芽及地面已萌发胞芽等)种植于实验室(贵州省贵阳市花溪区贵州大学校园内)用于后期形态观察。
1.2 方 法
1.2.1 形态发育特征观察选择种植个体观察植株生长和胞芽发育的形态变化,观察时期从胞芽恢复活动开始,采用肉眼直接观察或Olympus SZ51体式显微镜观察,内容包括胞芽发生时间、着生位置等。
1.2.2 石蜡切片采用常规石蜡切片方法[15]进行结构特征观察,使用Leica RM2016切片机切片,番红固绿法染色,用Olympus BX53生物显微镜进行观察并拍照,测量羽叶上、下表皮细胞的长和宽,叶肉细胞的长和宽;羽叶叶脉木质部的直径,测量结果为20个视野中测得数值的平均值±标准差,使用Excel进行统计分析。
1.2.3 电镜扫描对不同发育时期的稀子蕨胞芽进行取材并清理干净,切为5 cm ×5 cm大小左右的含胞芽的小段,材料采用修改双固定法[16]进行固定(材料仅用2.5%戊二醛于4 ℃固定14 h),之后用0.1 mol/L磷酸缓冲液(PBS)清洗,使用不同浓度梯度的叔丁醇/酒精混合液脱水,最后经冷冻干燥后镀金,使用S-3400N扫描电子显微镜观察并拍照。
2 结果与分析
2.1 稀子蕨胞芽发育的形态学变化
稀子蕨大多生长于海拔500~1 600 m喜阴多湿的密林下环境中(图版Ⅰ,A)。贵阳地区(海拔1 084~1 216 m)遮荫条件下移栽的稀子蕨植株从5~6月份开始恢复活动,地上羽叶发育可划分为5个时期:萌动期、卷拳期、展叶期、成熟期及衰老期,在展叶期伴随着胞芽发育(图版Ⅰ)。萌动期从5月底至6月初开始,以胞芽中的休眠叶原基恢复生长,以外围顶端卷曲叶原基从向内紧密包裹转为向外伸展为标志(图版Ⅰ,B)。叶原基顶端在萌动期不断伸长生长,以6月上旬顶端分化形成向心卷曲成拳状的小叶复合体为标志显示进入卷拳期(图版Ⅰ,C)。卷拳期叶柄不断生长而伸长,在6月中旬开始稀子蕨逐渐进入展叶期,即顶端的小叶复合体的叶片慢慢地开始展开(从基部叶开始到顶端),展叶过程中叶轴逐渐从向心弯曲转变为离心弯曲伸长(图版Ⅰ,D),随着叶轴展开5对二回羽叶时,三回羽叶的小羽叶展开2~4片,叶轴完全展开后2~4天,小羽叶完全展开。从8月开始稀子蕨基本步入成熟期,即稀子蕨各回羽叶展叶完毕,其叶片常具有5~20个二回羽叶,随着展叶完成,成熟期植株高度不再改变。成熟期持续3~4个月后,11月底稀子蕨进入衰老期,即从稀子蕨顶端的小羽叶边缘黄化开始,顶端慢慢衰老倒伏。
胞芽发育从羽叶的展叶期开始,包括原基发育期、胞芽分化期、胞芽膨大期和胞芽生长成熟期。(1)胞芽原基发育期:胞芽原基是位于第2~11二回羽叶轴与初级叶轴分叉处出现的无结构分化的凸起结构,胞芽原基在7月初展叶前期开始发生,逐渐长大至肉眼可见,呈黄绿色(图版Ⅰ,E、F)。(2)胞芽分化期:胞芽原基继续生长,从7月中旬开始原基开始分化出现幼嫩叶轴;至7月底,胞芽原基分化出多个叶轴使整个胞芽呈现出指形“锚”状结构、颜色呈绿色(图版Ⅰ,G、H、I)。(3)胞芽膨大期:8月初以后,锚状胞芽具有的叶轴数量不再增加(约为5~9个叶轴),但在9月前胞芽的叶轴长度与直径持续增加、颜色保持为绿色(图版Ⅰ,F、H、I)。(4)胞芽成熟期:在9月后,“锚”状胞芽生长不明显,其表面腺毛颜色从绿色转变为棕黄色,在11月或更晚时期其基部形成离层,在有风或触碰时可自然掉落(图版Ⅰ,J)。胞芽发育期持续时间较长,一般为4~5个月。成熟胞芽可在母体上直接发育长出新叶形成新植株(图版 Ⅰ,K),或掉落地面并在环境条件适宜条件下生长为新的植株。在胞芽发育过程中(10月份左右),成熟叶片上形成孢子囊群产生大量孢子,10月中、下旬孢子成熟。

图版Ⅰ 稀子蕨胞芽发育形态变化A. 稀子蕨野外生长生境;B. 稀子蕨胞芽萌动期;C. 卷拳期;D. 展叶期;E~J. 胞芽发育期: E. 胞芽原基发育期;F. 胞芽原基;G和H. 含两个叶原基的胞芽;I. 胞芽分化多个叶原基;J. 胞芽成熟期;K. 成熟胞芽母体上发育新叶。箭头所指为胞芽Plate I Morphological variation on the gemmae development of M. henryiFig.A. Wild habitat of M. henryi; Fig.B. The active recovery stage of M. henryi gemmae; Fig.C. Rolling fist stage; Fig.D. Leaf unfolding stage; Fig.E-J. Gemmae development: Fig.E. Gemmae primordial development stage; Fig.F. Gemmae primodium; Fig.G & H. Gemmae differentiation with two leaf primordia; Fig.I. Gemmae with multiple leaf primorda; Fig.J. Mature gemmae; Fig.K. Leaf development of gemmae on the mother plant. The arrow refers to the gemmae
2.2 稀子蕨胞芽发育的解剖学研究及扫描电镜观察
胞芽原基发育前,叶轴分叉处表面平坦、有较密集分布的短腺毛覆盖(图版Ⅱ,A);含表皮组织、薄壁组织、维管束组织等结构,表皮细胞较小、排列紧实,而薄壁细胞较表皮细胞大且排列紧密(图版Ⅱ,E)。在胞芽原基发育时期,叶轴分叉处可见有点状突起(图版Ⅱ,B);叶轴分叉处表皮细胞下的薄壁组织中形成一团体积较小、排列紧密的细胞团并继续分裂生长形成胞芽原基(图版Ⅱ,F)。在胞芽分化时期,胞芽原基不断分化叶原基最终呈指形“锚”状结构(图版Ⅱ,C),原基分化时顶端分生组织分化出第一重叶原基,包裹着亚顶端分生组织顺序分化出的第二重叶原基(图版Ⅱ,G)。在胞芽分化完全时期,肉眼可见胞芽分化出多个叶原基,整体呈指形“锚”状,最先分化出的外围叶原基包围着中心正在分化的叶原基(图版Ⅱ,D);单个叶原基横切图显示整体细胞呈圆形,内含两束维管束,呈“八”字形排列(图版Ⅱ,H),维管束内木质部和韧皮部呈“一”字形排列,木质部为“海马”形(图版Ⅱ,I)。

图版Ⅱ 稀子蕨胞芽发育过程Vb. 维管束; Pc. 薄壁细胞; Pr. 原生木质部; Me. 后生木质部; Ph. 韧皮部。箭头所示为胞芽。A~D. 胞芽发育的外部形态变化: A. 胞芽未分化期, B. 胞芽原基发育期, C. 胞芽分化期, D. 成熟胞芽;E~I. 孢芽发育的解剖学变化: E. 胞芽未分化期, F. 胞芽原基发育期, G. 胞芽分化期, H. 成熟胞芽(横切), I. 成熟胞芽内部维管束;J~L. 胞芽发育的外部超微形态变化: J. 胞芽未分化期, K. 胞芽原基发育期, L. 胞芽分化期Plate Ⅱ The gemmae development of M. henryiVb. Vascular bundle; Pc. Parenchyma cell; Pr. Protoxylem; Me. Metaxylem; Ph. Phloem. The arrows refer to gemmae. Fig.A-D: Morphological characteristics of gemmae development: Fig.A. Without gemmae, Fig.B. Gemmae primordia, Fig.C. Differentiation of gemmae, Fig.D. Mature gemmae; Fig.E-I: Anatomical characteristics of gemmae development: Fig.E. Without gemmae, Fig.F. Gemmae primordia, Fig.G. Differentiation of gemmae, Fig.H. Mature gemmae (transverse), Fig.I. The vascular bundle of the mature gemmae; Fig.J-L: Electron scanning microscope observation of the gemmae development: Fig.J. Without gemmae, Fig.K. Gemmae primordia, Fig.L. Differentiation of gemmae
扫描电子显微镜观察胞芽未分化时期、胞芽原基发育时期、胞芽分化时期,结果皆显示叶轴表面覆盖大量腺毛,并且胞芽原基和胞芽表面皆被腺毛覆盖,因此大量腺毛的存在影响了对于胞芽发育的观察(图版Ⅱ,J、K、L)。
2.3 稀子蕨营养器官解剖结构
稀子蕨的小羽叶叶片较薄,由6~7层细胞组成(图版Ⅲ,A):上表皮细胞呈椭圆形(长22.59 ± 4.52 μm,宽22.23 ± 2.97 μm);下表皮细胞亦呈椭圆形(长21.59 ± 4.69 μm,宽17.84 ± 3.16 μm),其间分布有气孔;叶肉细胞排列疏松,无明显栅栏组织和海绵组织分化,呈椭圆形或者无规则形(长20.80 ± 4.36 μm,宽17.51 ± 2.95 μm),含叶绿体,细胞间隙大,形成发达的通气组织。稀子蕨小羽叶的叶脉呈分叉状,无主脉侧脉之分,含维管束,木质部(直径17.45 ± 4.28 μm)细胞数量少为4~5个(图版Ⅲ,B)。
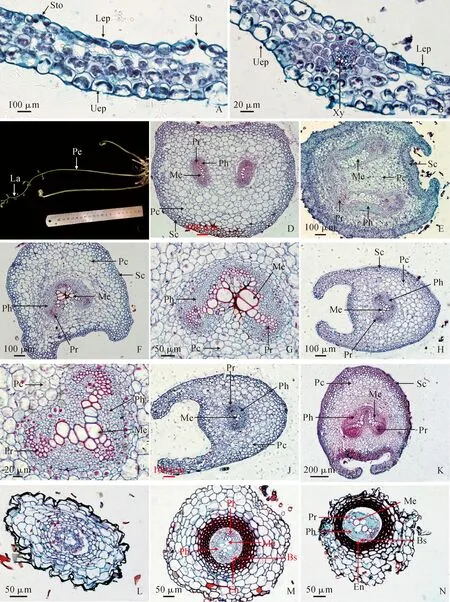
图版Ⅲ 稀子蕨羽叶、叶柄、叶轴和根的解剖结构Sto. 气孔;Uep. 上表皮;Lep. 下表皮;Xy. 木质部;Pe. 叶柄;La. 叶轴;Pc. 薄壁细胞;Pr. 原生木质部;Me. 后生木质部;Ph. 韧皮部;Sc. 厚壁细胞;En. 内皮层;Bs. 中柱鞘。A. 羽叶;B. 羽叶的叶脉;C. 稀子蕨;D. 叶柄基部横切;E. 叶柄中部横切;F. 叶柄上部横切;G. 叶柄上部横切维管束;H. 叶轴下部横切;I. 叶轴横切维管束;J~K. 叶轴上部横切;L. 幼根横切; M. 成熟根横切;N. 老根横切Plate Ⅲ Anatomy of the leaves, petioles, leaf axes, and roots of M. henryiSto. Stomata; Uep. Upper epidermis; Lep. Lower epidermis; Xy. Xylem; Pe. Petiole; La. Leaf axis; Pc. Parenchyma cell; Pr. Protoxylem; Me. Metaxylem; Ph. Phloem; Sc. Sclerenchyma; En. Endothelium; Bs. Bundle sheath. Fig.A. Feather leaf; Fig.B. Vein of the feather leaf; Fig.C. Seedling of M. henryi; Fig.D. The base of petiole (transverse); Fig.E. The middle petiole (transverse); Fig.F. The upper petiole (transverse); Fig.G. Vascular bundle of upper petiole; Fig.H. Lower leaf axis (transverse); Fig.I. Vascular bundle of leaf axis (transverse); Fig.J-K. Upper leaf axis (transverse); Fig.L. Young root (transverse); Fig.M. Mature root (transverse); Fig.N. Old root (transverse)
稀子蕨叶柄覆盖稀疏短腺毛,横切整体呈椭圆形,含有1~2束维管束。叶柄基部、中部含2束维管束呈“八”字形,木质部韧皮部呈“一”字形排列,木质部为“海马”形(图版Ⅲ,D、E),叶柄中部近轴面的周皮组织会突出从边缘两端同向延伸,使中部近轴面外表形成“沟”(图版Ⅲ,E);叶柄上部含一束维管束,呈“心形”形状,其中木质部排列形状呈两“海马”头部结合在一起的倒“V”字形(图版Ⅲ,F、G),叶柄上部的近轴面的周皮组织也会突出从边缘两端同向延伸形成“沟”(图版Ⅲ, F)。稀子蕨叶轴也有较稀疏的短腺毛覆盖,石蜡切片可观察到叶轴横切整体呈椭圆状,含一束维管束,呈“心形”形状,其木质部排列形状呈两“海马”头部相结合组成的倒“V”字形(图版Ⅲ,I)。在近轴面同样部分周皮组织会突出从边缘两端向内同向延伸形成“沟”(图版Ⅲ,H、J、K)。
稀子蕨根横切呈椭圆形,由表皮、皮层、中柱构成。幼根表皮结构不清晰;外皮层中靠近维管束的有由4~6层皮层细胞形成的厚壁细胞,其染色较周围皮层细胞略深,呈长方形;维管束内结构分化不清晰、无髓分化(图版Ⅲ,L),表皮和维管束结构不清晰可能是因为切片质量不佳。成熟根的表皮、皮层、中柱结构完整:表皮由一层细胞构成,呈椭圆形;皮层细胞由外皮层、内皮层组成,外皮层大多由11层细胞组成,接近表皮的5~6层细胞较大且形状不规则,靠近维管束的4~6层外皮层细胞形成的厚壁细胞染色加深,细胞较小且为长方形,内皮层紧挨中柱,由一层细胞构成,呈椭圆形;维管束结构分化完整,中柱鞘由一层细胞构成且紧挨内皮层,木质部为二原型含初生木质部与后生木质部,韧皮部分布于木质部两侧(图版Ⅲ,M)。老根可明显观察到结构较成熟根时期发生改变,具体表现为:表皮已脱落不在,皮层正在脱落中,维管束外由皮层形成的厚壁细胞染色变得更深,中柱内也有部分脱落(图版Ⅲ,N)。
3 讨 论
在叶片发育上,稀子蕨和大多数蕨类一样经历萌动期、卷拳期、展叶期、成熟期和衰老期。蕨类植物羽叶叶肉细胞无或有栅栏组织与海绵组织分化:海金沙(Lygodiumjaponicum)、井栏边草(Pterismultifida)、肾蕨(Nephrolepiscordifolia)、铁线蕨(Adiantumcapillus-veneris)、银脉凤尾蕨(P.ensiformisvictoriae)和顶芽狗脊等羽叶无分化[11,17-18],而对卤蕨(Acrostichumaureum)和尖叶卤蕨(A.speciosum)羽叶有分化[19]。本研究显示稀子蕨羽叶叶肉细胞中不含栅栏组织和海绵组织分化,较大的叶肉细胞间隙能储存较多气体,是其阴生环境的适应特征[17]。稀子蕨叶柄基部、中部含2束维管束,上部及叶轴含1束维管束,这种自叶柄基部向上到叶轴维管束向内靠近融合成一束的结果与叶红环[11]、郭庆梅等[20]、周凤琴等[21]报道的结果相似。稀子蕨的根无髓,在维管束外有一圈由多层皮层细胞构成的染色较深的环,这一结果与单芽狗脊蕨、爬树蕨(Arthropterispalisotii)、龙津蕨(Mesopteristonkinensis)、铁角蕨属与鸟巢蕨属等的研究结果相似[11,22-24]。
与其他蕨类植物相比,稀子蕨复叶上发育胞芽的数量及其在叶片上的相对位置不同:稀子蕨每片复叶(具5~20个二回羽叶)上发育的0~4个胞芽发生于第2~11个二回羽叶与叶轴分叉处。其他蕨类植物的复叶上常单生胞芽,或着生于叶轴近先端[如铁角蕨科倒挂铁角蕨(Aspleniumnormale)、蹄盖蕨科芽胞蹄盖蕨、鳞毛蕨科陈氏耳蕨(Polystichumchunii)、乌毛蕨科单芽狗脊等蕨类],或着生于叶轴先端以下[如蹄盖蕨科胎生蹄盖蕨(A.viviparum),或着生于叶轴上部[如铁角蕨科胎生铁角蕨(A.indicum),或着生于叶轴基部[如铁角蕨科滇南铁角蕨(A.microtum)][25]。稀子蕨胞芽发育过程包括胞芽原基发育期、胞芽分化期、胞芽膨大期、胞芽成熟期四个时期。稀子蕨和单芽狗脊蕨的胞芽发育过程类似:在展叶期于叶轴与羽叶分叉处的表皮细胞下的薄壁细胞形成胞芽原基,之后胞芽原基分化并膨大,最后胞芽成熟[11]。成熟胞芽基部与母体组织间形成离层后断裂脱离母体掉落土壤,待环境适宜胞芽萌发形成新的植株;或者胞芽在母体上直接发育长出须根和新叶,待落地后直接发育成完整植株[10]。稀子蕨和单芽狗脊蕨胞芽发育的不同之处在于:单芽狗脊蕨在分化时期,胞芽原基表皮外的部分细胞会形成表皮衍生结构即鳞片,鳞片包裹胞芽且成熟后呈红棕色,胞芽近球形其着生位置大多分布于倒数第三回羽叶和叶轴分叉处且位于叶片背面;而稀子蕨胞芽发育中并没有表皮衍生结构的分化即无鳞片,但被覆短腺毛成熟呈黄棕色,胞芽呈指状“锚”形且叶原基为有限分化,胞芽着生于第2~11回羽叶和叶轴分叉处,位于叶片正面[11]。
稀子蕨胞芽发育过程和天南星科(弥勒魔芋、滴水珠、半夏)、薯蓣科(薯蓣)、百合科(卷丹百合、淡黄花百合)的珠芽在发生位置和发育过程上具有相似特征,在发生位置上都发生于表皮细胞下的薄壁细胞层;在发育过程上都有原基发育期、分化期、膨大期和成熟期[2-4, 6-8]。无性繁殖器官与有性生殖的种子类似,都需要在发育中后期积累有机物供其在后续生长季节中的萌发与生长。与淡黄花百合珠芽类似[3],稀子蕨胞芽从膨大期开始储藏物在已分化的叶器官中积累(图版Ⅱ,D、H);而单芽狗脊蕨胞芽与半夏、薯蓣等珠芽内的储藏物从膨大期开始分别积累在着生叶的茎组织或形成的小块茎内[2,8,11]。种皮与果皮等结构是种子的保护结构,无性繁殖器官通常也发育出保护结构,如半夏等珠芽小块茎顶端的膜质鳞片[8]、淡黄花百合珠芽的膨大增厚鳞片[3]、单芽狗脊蕨胞芽外部的鳞片等[11]。但是,稀子蕨的胞芽在发育过程中缺少鳞片结构(图版Ⅱ,C、G),这可能与稀子蕨适应分布于湿润、温暖的环境中有关。
猜你喜欢
科学大众·教师版(2022年6期)2022-05-23
河北农业大学学报(2021年5期)2021-11-10
北京航空航天大学学报(2021年9期)2021-11-02
植物研究(2021年1期)2021-02-26
新疆农业科学(2020年9期)2020-10-13
植物研究(2020年6期)2020-03-05
河南农业科学(2019年9期)2019-09-24
计算机应用(2018年10期)2018-11-22
图书与情报(2017年2期)2017-06-05
儿童故事画报·发现号趣味百科(2015年7期)2015-10-23
