
连翘GLK基因家族鉴定及响应光照表达分析
2022-12-28 10:34赵峥畑张晓璐程堂仁王佳张启翔潘会堂
河南农业大学学报 2022年5期
赵峥畑,张晓璐,程堂仁,王佳,张启翔,潘会堂
(北京林业大学园林学院/花卉种质创新与分子育种北京市重点实验室国家花卉工程技术研究中心/城乡生态环境北京实验室,北京 100083)
Golden2-like转录因子,又称为GLK或G2-like,在植物中广泛存在,属于GARP转录因子超家族的一员。GLK首次在黄化的玉米(Zeamays)中被发现并命名[1],随后研究人员在拟南芥(Arabidopsisthaliana)[2]、水稻(Oryzasativa)、辣椒(Capsicumannuum)[3]和番茄(Solanumlycopersicum)[4]等多种植物中发现了GLK基因。GLK转录因子包含2个高度保守的结构域:一个Myb-DNA结合域(DBD),一个位于C末端的GCT-box[5]。DBD处的AREAAA六肽序列在GARP家族中高度保守,在绿藻和陆地植物中均有[6],而GCT-box仅存在于陆地植物中。此外,一些亚族的成员具有保守的Myb-CC-LHEQLE基序[7]。
GLK在调控植物叶绿体发育和光合作用中发挥着重要的作用[8-9],此外参与细胞分化、逆境胁迫响应、果实品质和叶片衰老等生物学进程[10-11]。研究表明,GLK蛋白可以直接结合到一些天线蛋白如LHCs和叶绿素合成酶HEMA1、CHLH和CRD1等基因的启动子上,正调控其转录,从而影响叶绿体发育和色素合成[10]。在甜瓜(Cucumismelo),红掌(Anthuriumandraeanum)和紫薇(Lagerstroemiaindica)等多数黄叶突变体中发现,其叶绿素含量偏低、叶绿体结构存在一定缺陷,GLK表现为下调表达[12-14]。而分别过表达苹果的MdGLK1基因和甜瓜的CmGLK基因能够恢复拟南芥glk1glk2突变体的黄化表型[14-15]。
连翘属(Forsythia)植物为木犀科(Oleaceae)的落叶灌木,早春先叶开花,在园林绿化中应用广泛。金叶连翘(F.koreana‘Suwon Gold’)兼具观叶和观花的效果,是重要的彩叶灌木;同时作为光敏型叶色突变体,在荫蔽条件下叶色转绿,是研究色素生理、光合作用等的理想材料[16]。申建双[17],孙旺旺等[18]发现,金叶连翘叶片的叶绿素含量显著低于绿叶对照组,叶绿体结构发育停滞、类囊体结构异常。2020年,LI等[19]首次对连翘基因组进行了测序,为连翘的分子研究提供了重要的参考基础。SHEN等[20]建立了连翘病毒介导的基因沉默(virus induced gene silencing,VIGS)体系,为基因功能研究提供了便利的技术条件。
目前,人们在葡萄(Vitisvinifera)[21]、巨桉(Eucalyptusgrandis)[22]、番茄[23]等多种植物中已报道GLK基因家族信息,但对连翘GLK基因家族的系统研究尚未见报道。本研究根据已发表的连翘基因组数据进行GLK基因家族的全面鉴定,采用生物信息学方法对家族成员进行蛋白理化性质、系统进化、基因结构,以及启动子顺式作用元件的分析,并通过转录组数据分析FsGLK基因在黄叶型和绿叶型连翘在遮光前后的表达变化,结合实时荧光定量PCR技术锚定关键基因,为进一步研究连翘FsGLK转录因子的功能奠定基础。
1 材料与方法
1.1 连翘GLK家族成员提取鉴定及蛋白理化性质预测
连翘全基因组数据下载自NCBI数据库(WIPI00000000.1)[19],拟南芥GLK基因家族的蛋白序列下载自TAIR数据库(https://www.arabidopsis.org/browse/genefamily/G2like.jsp)。首先以拟南芥GLK蛋白作为种子序列,利用BLASTP程序在连翘蛋白序列中搜索比对,以e<1e-5为阈值,手动剔除氨基酸数小于100的序列,得到候选的连翘GLK基因。再用HMMER 3.0程序对候选序列进行保守结构域PF00249鉴定。该结构域的隐马尔可夫模型(hidden Markov model,HMM)下载自Pfam数据库(https://pfam.xfam.org/),检索阈值设置为e<1e-5。进一步用在线工具SMART(http://smart.embl-heidelberg.de/)对所有候选序列进行保守结构域验证。最终获取连翘基因组库中GLK成员的基因号及对应的DNA和蛋白序列,依据在染色体上的位置对其进行命名。利用在线工具Prot Param(https://web.expasy.org/protparam/)预测连翘GLK蛋白的理化性质,包括氨基酸数量、分子量、等电点、不稳定性指数和等[24]。通过WoLF PSORT(https://wolf psort.hgc.jp/)预测连翘GLK的亚细胞定位[25]。
1.2 GLK基因家族系统进化树构建
用MEGA 11软件对拟南芥和连翘GLK家族蛋白序列进行Clustal W比对,并构建系统进化树[26],选择邻接法(NJ),自展值为1 000。进化树结果用EvolView进行绘制[27]。
1.3 连翘GLK基因家族的保守基序和基因结构分析
使用在线软件MEME进行保守基序鉴定和分析(最大基序数选择20,基序长度6~50)[28]。根据连翘基因组的注释文件对FsGLK的基因结构进行分析。用TBtools软件对上述结果进行绘图[29]。
1.4 连翘GLK基因家族启动子顺式作用元件预测
提取连翘GLK家族成员上游2 kb碱基序列作为启动子区域,提交至PlantCARE数据库预测顺式作用元件[30],结果用TBtools软件进行可视化分析[29]。
1.5 连翘GLK基因家族响应光照表达分析
潘会堂课题组前期以黄叶型的金叶连翘为母本、绿叶型的连翘品种‘Courtaner’为父本进行杂交获得F1代叶色分离群体,从中筛选出黄叶型和绿叶型各10个株系,每个株系扩繁30株以上,以1 a生扦插苗为材料于2017年5月进行遮光处理,以全光条件作为对照。遮光30 d后,取全光下的绿叶型(L-G)、全光下的黄叶型(L-Y)、遮光下的绿叶型(S-G)、遮光下的黄叶型(S-Y)连翘叶片进行转录组测序。将4组(L-G、L-Y、S-G、S-Y)材料每个株系各取一片完整叶片(用10个株系混样提取RNA,作为一次生物学重复,重复3次取样),液氮冷冻后送公司测序(诺禾致源科技股份有限公司,北京)。采取Trizol的方法分别提取总RNA,并分别对RNA的浓度、纯度及其样品的完整性进行检测,随后分别建库。文库构建并质检合格后,随即进行上机进行Illumina HiSeq测序[17]。处理期间全光下的日最大光照度为1 300~1 500 μmol·m-2·s-1,遮光下的日最大光照度为390~450 μmol·m-2·s-1。
本研究根据转录组数据进行连翘GLK基因在不同叶色及不同光照时间的表达分析,并用TBtools软件绘制FsGLK基因表达热图[29]。
进一步设置单层遮荫网(遮光率50%,S1-Y)、双层遮荫网(遮光率75%,S2-Y)两种处理,对黄叶型连翘进行遮光。在遮光15 d后撤掉遮荫网恢复全光照。试验期间全光下的日最大光照度为500~1 500 μmol·m-2·s-1。分别在遮光后24 h(T1)、复光后24 h(T2)时采集叶片,提取RNA并反转录合成cDNA,两时期均以全光下的绿叶型(L-G)和黄叶型(L-Y)作为对照,利用实时荧光定量PCR技术检测关键GLK基因在不同处理条件下叶片中的表达情况。
2 结果与分析
2.1 连翘GLK基因家族成员及其理化性质
通过从连翘的基因组库中查找筛选,鉴定出44个GLK基因家族成员,将其命名为FsGLK1~FsGLK44。SMART验证保守结构域发现,除了myb_SHAQKYF以外,部分家族成员含有Myb_CC_LHEQLE(pfam14379)或REC_typeB_ARR-like(cd17584)的保守结构域。FsGLK家族成员的具体蛋白理化性质分析结果如表1所示。FsGLKs蛋白的氨基酸数量为197~693,相对分子质量为 22.6~76.1 kD。理论等电点为5.37~9.28,其中19个FsGLK为碱性蛋白(PI>7),25个FsGLK为酸性蛋白(PI<7)。不稳定系数为32.04~69.39,其中仅FsGLK14、FsGLK30、FsGLK35系数小于40,为稳定蛋白,其余均为不稳定蛋白。由亲水性指数分析可知,GLK基因家族编码的蛋白均为亲水蛋白。亚细胞定位预测结果显示,FsGLK7定位在微体中,FsGLK30和FsGLK43定位于叶绿体中,其余31个成员均定位在细胞核中。
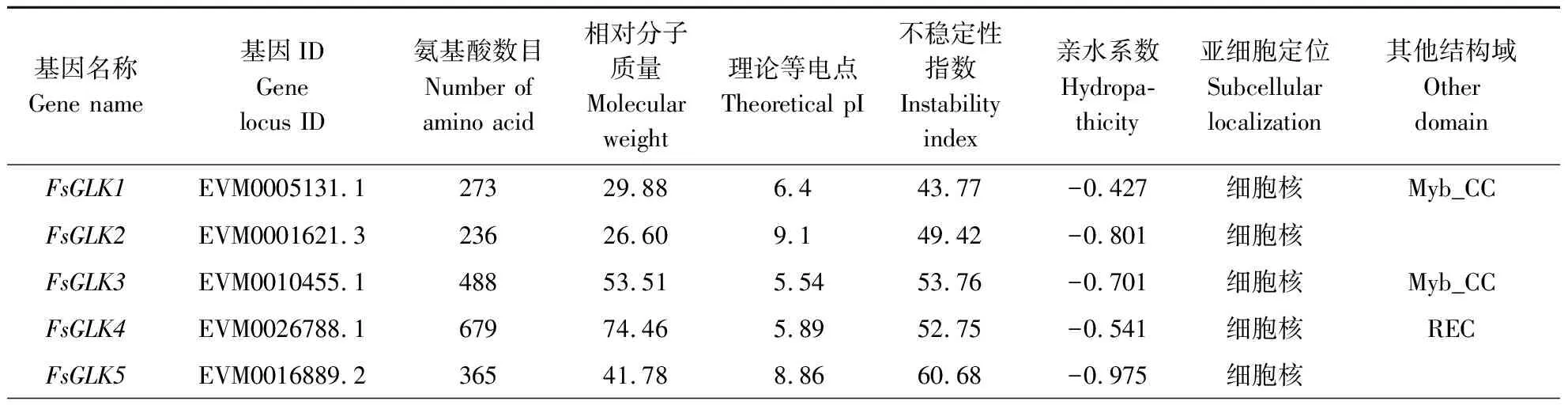
表1 连翘GLK转录因子家族信息及理化性质Table 1 Informationandphysicochemicalpropertiesofthe GLK TF familyof F.suspensa
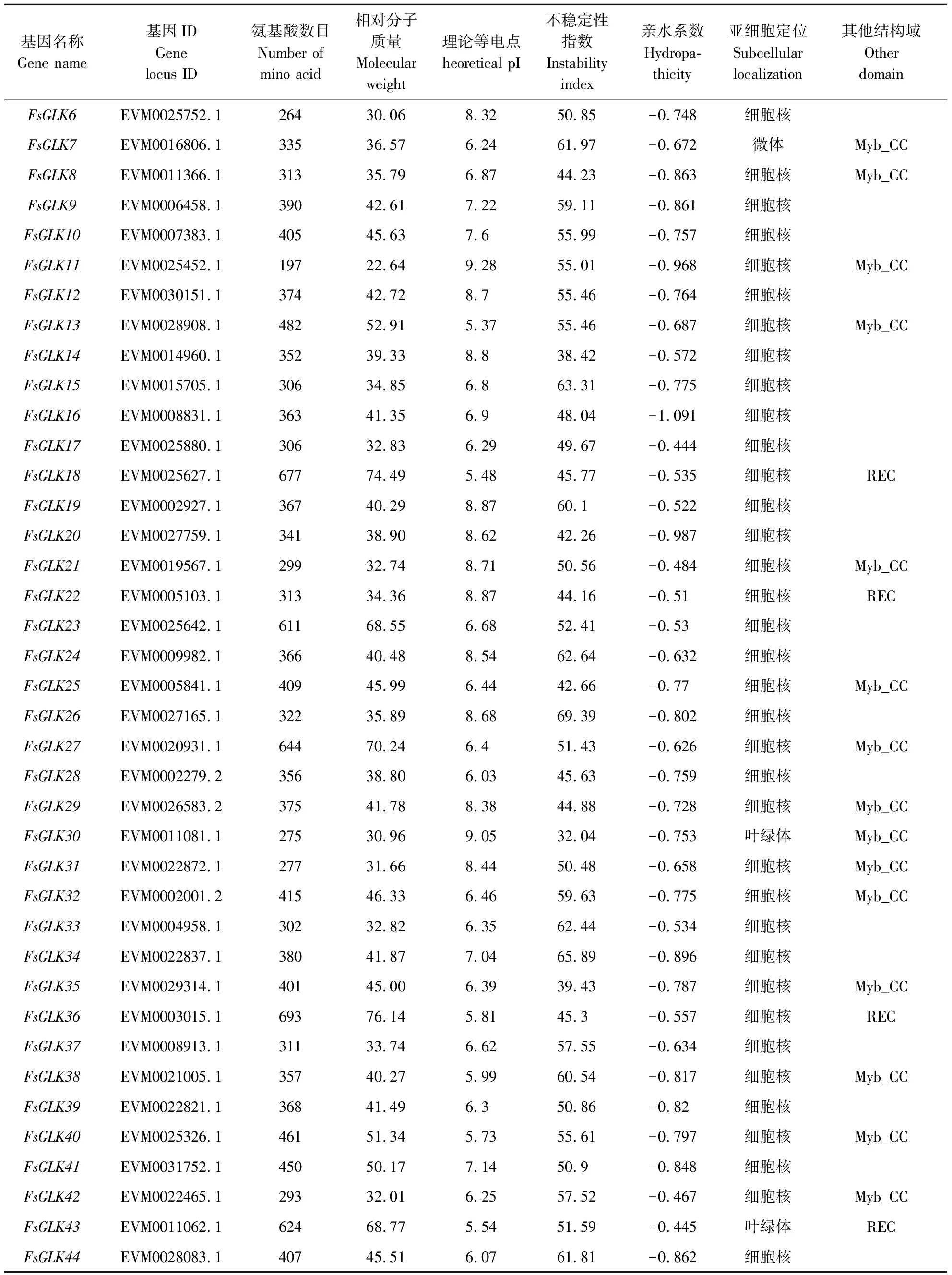
续表 Continuing table
2.2 GLK基因家族的系统发育分析
以拟南芥GLK蛋白序列和鉴定到的44个连翘GLK蛋白序列构建系统进化树,结果如图2所示。连翘的44个GLK基因分为5个亚族Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ,分别包含8、11、8、8和9个连翘GLK成员和20、12、8、11、13个拟南芥GLK成员。其中第Ⅰ亚族中的FsGLK21与AT2G06020.1、第Ⅲ亚族中的FsGLK24与AT2G03500.1、第Ⅳ亚族中的FsGLK5与AT1G32240.1、第Ⅴ亚族中的FsGLK30与AT5G45580.1分别聚类为1支,说明上述基因同源性较高。结合2.1中保守结构域分析,发现第Ⅰ和第Ⅴ亚族的17个成员均含有Myb_CC_LHEQLE保守结构域,含有REC_typeB_ARR-like保守结构域的4个成员均属于第Ⅱ亚族。
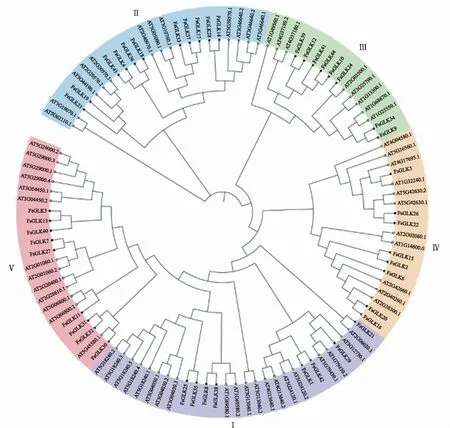
图1 连翘和拟南芥GLK蛋白的系统进化树Fig.1 Phylogenetic tree of GLK proteins in F.suspensa and A.thaliana
2.3 连翘GLK基因的保守基序和基因结构分析
为了进一步研究连翘GLK基因家族成员在结构上的进化关系,对44个FsGLK基因上的保守基序(motif)和基因结构进行了分析。保守基序分析结果(图2-B)显示,每个连翘FsGLK蛋白序列中都包含2~10个motif,其中第Ⅴ亚族的成员motif数量最少,仅有2个,第Ⅲ亚族的motif数量最多,有7~10个。各亚族内的motif种类和数量相似,保守性较强,其中motif 1和motif 2在FsGLK所有蛋白序列中保守分布。基因结构分析结果(图2-C)显示,连翘FsGLK家族各成员的非编码区(UTR)、内含子(Intron)数量、基因长度等均存在差异。其中FsGLK14等10个成员不含UTR,FsGLK41仅含有5′端UTR,FsGLK11仅含有3′端UTR,认为这些基因功能可能存在差异。FsGLK基因长度为2~15 kb,内含子数量为0~11,其中FsGLK33没有内含子结构,FsGLK17、FsGLK37、FsGLK14和FsGLK28具有1个内含子,均属于第Ⅱ亚族。其余的家族成员均含有1个以上的内含子,大多数基因具有4~5个内含子,其中FsGLK23和FsGLK27的内含子数量为11。

注:A为基于FsGLK蛋白序列的系统发育树;B为FsGLK的保守基序,底部刻度尺是蛋白质的长度/aa;C为FsGLK的基因结构,底部刻度尺是基因的长度/bp。 Note:A represents phylogenetic tree based on the amino acid sequences of FsGLK; B represents the motif composition of FsGLK,and the bottom scale shows the protein length/aa.C represents the structure of FsGLK,and the bottom scale shows the gene length/bp.
2.4 连翘GLK基因启动子顺式作用元件分析
分析FsGLK基因启动子区上游2 000 bp的序列,预测的顺式作用元件结果如图3所示。连翘GLK基因家族成员的启动子类型主要包括光响应、胁迫响应(干旱、低温、防御应激、伤口等)、植物激素响应(脱落酸、茉莉酸甲酯、赤霉素、生长素、水杨酸等)元件,其次是一些参与细胞周期调控、昼夜节律、类黄酮生物合成基因调控相关的元件,还包括部分器官特异性表达(种子、根、胚乳等)相关的元件。在所有顺式调节元件中,各基因的启动子均含有与光响应相关的元件,且数量最多,平均占各基因元件总数的49%。光响应元件的种类主要有G-box、Box 4、TCT-motif、GT1-motif、GATA-motif、3-AF1 binding site、chs-CMA1a等23种。此外,FsGLK36和FsGLK44基因的启动子均含有1个与类黄酮生物合成调控基因相关的MYB结合位点(MBSI)。推测该基因能够响应上游基因的调控,参与调控类黄酮的合成。综上所述,FsGLK基因可能在连翘生长发育、响应环境刺激,特别是对光响应起到重要的作用。
2.5 连翘GLK基因在不同叶色及光强下的表达分析
对黄叶型和绿叶型的连翘杂交后代株系进行为期30 d的遮光处理,如图4所示。遮光的黄叶型连翘(S-Y)明显转绿,遮光的绿叶型连翘(S-G)的叶色变化不明显。取不同处理植株的叶片提取RNA并进行转录组测序,从测序数据中筛选出44个FsGLK基因的表达量数据,绘制热图如图5所示。
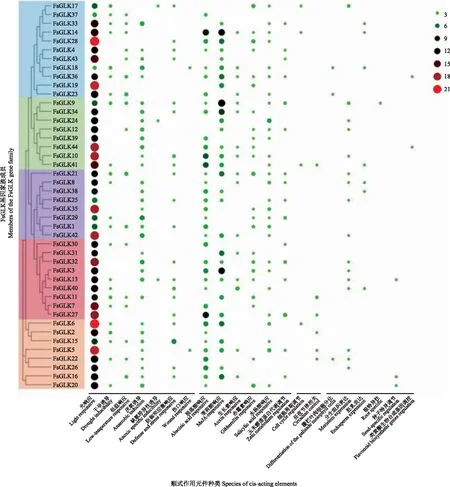
注:气泡大小代表顺式作用元件的数量。Note:The bubble size represents the number of cis acting elements.

注:L-G代表全光下绿叶型连翘;L-Y代表全光下黄叶型连翘;S-G代表遮光下绿叶型连翘;S-Y代表遮光性黄叶型连翘。Note:L-G represents green-leafed Forsythia under full light;L-Y represents yellow-leafed Forsythia under full light;S-G represents green-leafed Forsythia under shading; S-Y represents yellow-leafed Forsythia under shading.
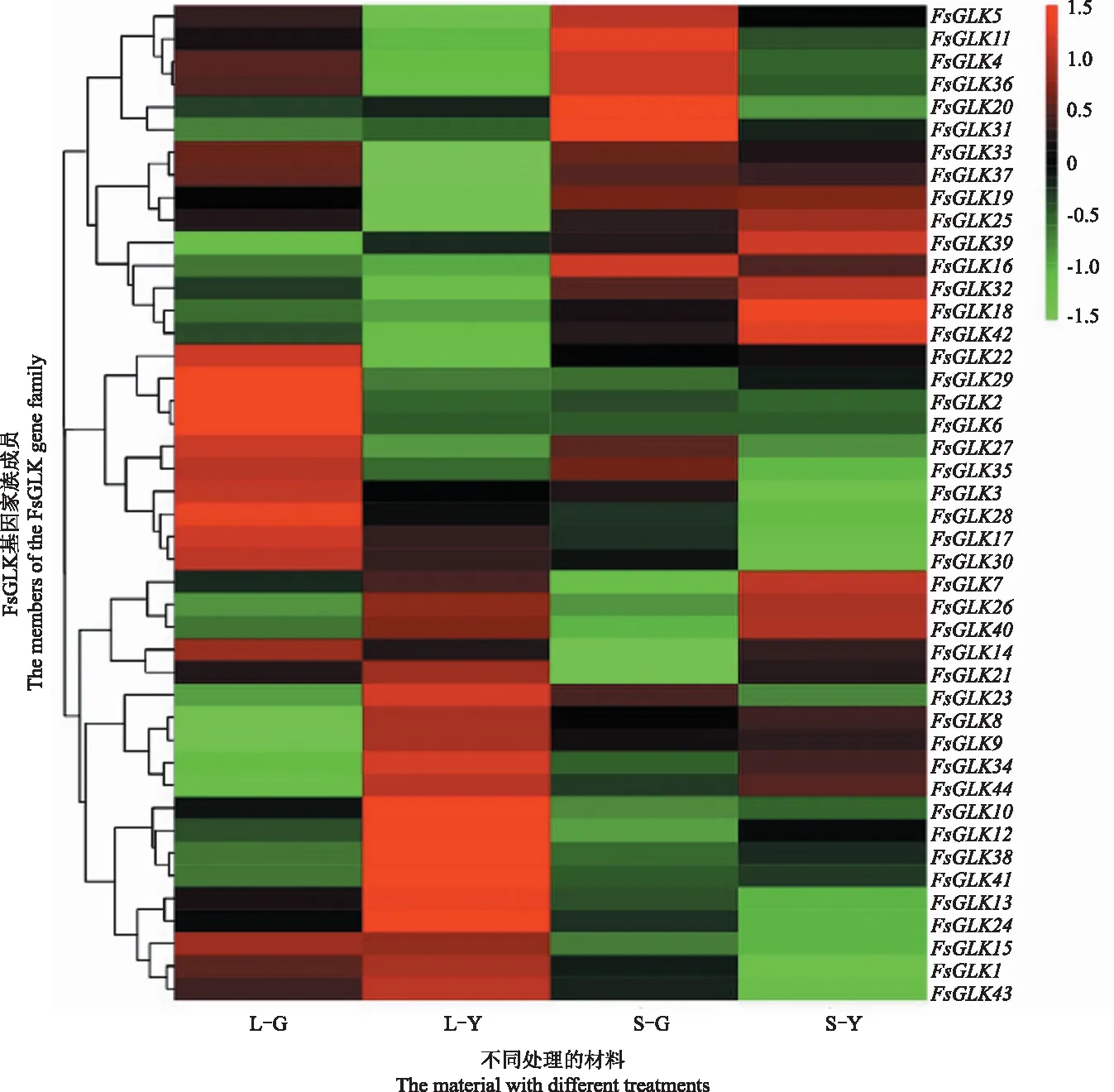
注:每1列代表不同处理的材料。L-G代表全光下绿叶型连翘;L-Y代表全光下黄叶型连翘;S-G代表遮光下绿叶型连翘;S-Y代表遮光性黄叶型连翘。Note:Each column represents a different treated material.L-G represents green-leafed Forsythia under full light;L-Y represents yellow-leafed Forsythia under full light;S-G represents green-leafed Forsythia under shading; S-Y represents yellow-leafed Forsythia under shading.
44个家族成员中,同一亚家族的表达趋势并非完全一致,推测不同的家族成员在对叶色的影响中具有特异的功能。在全光条件下有5个FsGLK基因在不同叶色连翘中差异表达,其中FsGLK9、FsGLK19、FsGLK27在绿叶型连翘中优势表达,FsGLK34、FsGLK41在黄叶型连翘中优势表达,推测这些基因与连翘的叶色呈现有直接关系。3个FsGLK基因在全光照和遮光处理条件下黄叶型连翘中差异表达,FsGLK19和FsGLK32在遮光后上调表达,FsGLK41在遮光后下调表达,可能与黄叶型连翘叶色响应光强环境有直接关系。综合来看,FsGLK19的表达模式与黄叶型连翘的形成及响应光照呈色的趋势一致。
进一步对黄叶型连翘进行50%、75%遮光及遮光后复光处理,探究FsGLK19的表达情况。结果如图6所示,FsGLK19在全光下黄色叶中(L-Y)的表达量显著低于绿色叶(L-G),遮光条件下随遮光率的提高,FsGLK19在黄色叶中显著上调表达,50%遮光(S1-Y)时的表达量是全光下的2.77倍,75%遮光(S2-Y)时的表达量是全光下的3.70倍。在恢复光照后,S1-Y和S2-Y处理的FsGLK19表达量均下调,且与全光下黄叶型连翘的表达量无显著差异(P>0.05)。可见FsGLK19在黄叶型连翘中受光照度的负调控,推测其在金叶连翘叶色呈现中具有重要作用。
3 结论与讨论
GLK首次在黄化的玉米中被发现,被认为与植物的黄化突变有关[1]。目前在多种植物中已鉴定出GLK基因家族的成员,例如葡萄有46个[20]、巨桉有40个[22]、陆地棉有146个[31]。本研究从连翘基因组中鉴定了44个GLK基因家族成员,蛋白理化性质分析表明,其序列长度、相对分子质量和等电点分布范围广。研究表明,GLK是植物细胞核主导叶绿体发育的正向调控转录因子,主要转录激活两类光合相关核基因的表达:一类是编码叶绿素生物合成的一系列关键酶,另一类是编码光系统Ⅰ、光系统Ⅱ、光电子传递复合体的各类亚基[32]。FsGLK成员主要定位在细胞核中,部分成员如FsGLK30、FsGLK43定位于叶绿体,可能与叶绿体的发育或叶绿素合成的功能实现相关。
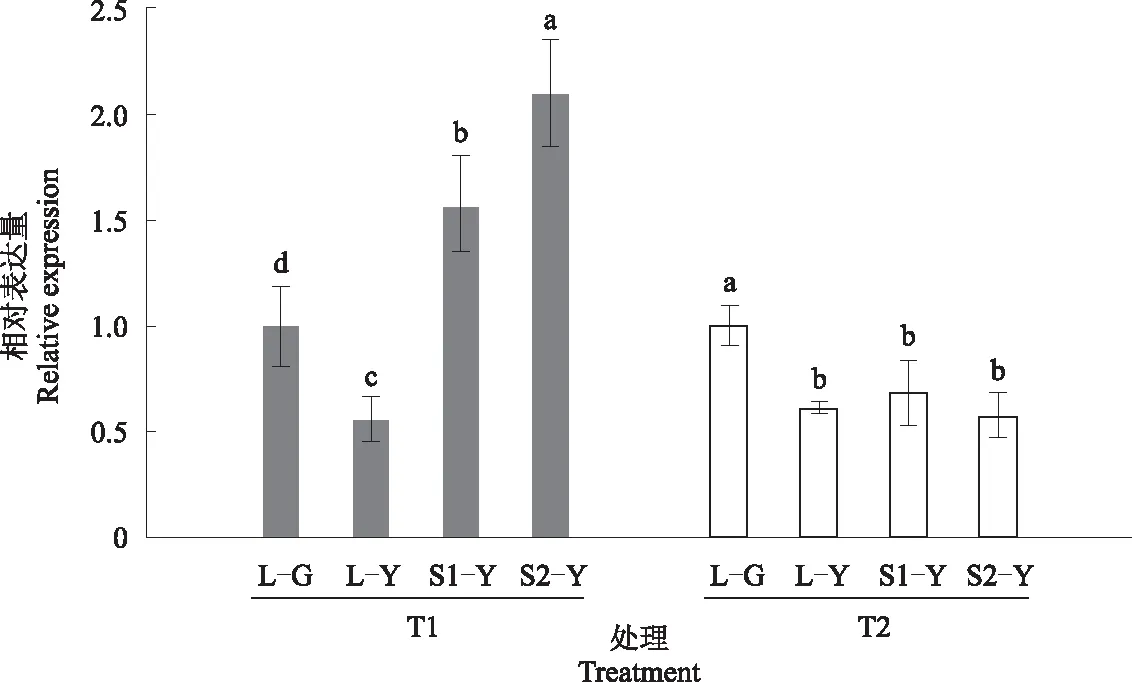
注:L-G代表全光下绿叶型连翘;L-Y代表全光下黄叶型连翘;S-G代表遮光下绿叶型连翘;S-Y代表遮光性黄叶型连翘。T1代表遮光后24 h;T2代表复光后24 h。不同小写字母代表不同处理之间差异显著(P<0.05)。Note:L-G represents green-leafed Forsythia under full light;L-Y represents yellow-leafed Forsythia under full light;S-G represents green-leafed Forsythia under shading;S-Y represents yellow-leafed Forsythia under shading.T1 represents 24 h after shading;T2 represents 24 h after re-lighting.Different lowercase letters represent significant differences between different treatments (P<0.05).
系统发育分析将连翘FsGLK家族归为5个亚族,各亚族成员具有相似的保守基序种类和分布形式,例如第Ⅰ和Ⅴ亚族均含有Myb_CC_LHEQLE保守结构域和motif 3,表明这些基因可能具有相似的功能[7]。而各成员的基因外显子与内含子结构有着较大的差异,与巨桉等的研究相似[22]。
FsGLK基因的启动子上含有大量顺式作用元件,含有重要的基因表达调控信息。对FsGLK家族成员基因的启动子序列进行顺式作用元件预测分析,显示各基因与光响应相关的元件数量最多,如G-box、Box 4、TCT-motif等。在水稻中的研究表明,OsPIL1能通过结合OsGLK1和OsGLK2启动子上的G-box(CACGTG)和E-box(CACATG)元件上调基因的表达,进而启动CAO、POR及一些捕光系统基因的表达,促进叶绿素合成[33]。
光敏黄叶突变体一般叶绿素含量降低,叶绿体结构发育不完全,而在遮光之后有所恢复。多数黄叶突变体研究均发现GLK基因下调表达,此外有研究表明,GLK基因能够响应光照度,在遮光后上调表达,例如,在楠木在遮光后,GLK及其他光合作用和叶绿素生物合成相关的基因表达量显著升高,认为该基因促进了叶片叶绿素含量增加和净光合速率的提高[34]。本研究以遮光和全光条件下不同叶色材料的转录组数据为基础,对FsGLK家族成员进行表达模式分析,发现FsGLK19在黄色叶和绿色叶连翘中差异显著,在黄叶遮光后显著上调表达。进一步通过实时荧光定量PCR的方法对该基因在遮光后和恢复光照处理过程中的表达情况进行分析,发现其受到光照度的负调控,推测FsGLK19在黄叶型连翘响应光照呈色中具有重要的作用。
综上所述,本研究利用生物信息学方法在连翘基因组中鉴定出44个FsGLK基因。结合在黄、绿叶型连翘中的表达差异及黄叶型连翘响应光照过程中基因表达量的变化,发现FsGLK19在黄叶中受到光照度的负调控,后续可进一步对该基因进行功能分析。
猜你喜欢
今日农业(2022年2期)2022-11-16
今日农业(2022年16期)2022-09-22
通信电源技术(2022年1期)2022-06-16
今日农业(2021年21期)2022-01-12
科学导报(2021年48期)2021-08-10
今日农业(2020年19期)2020-12-14
今日农业(2020年16期)2020-09-25
中国计算机报(2019年45期)2019-12-27
中华诗词(2019年12期)2019-09-21
阅读与作文(英语高中版)(2018年9期)2018-12-28
