
古基因组学与人类起源演化的研究进展
——2022年诺贝尔生理学或医学奖解读
2022-12-22 04:12吉植王传超
自然杂志 2022年6期
吉植,王传超②†
①厦门大学 社会与人类学院,福建 厦门 361005;②厦门大学 生命科学学院细胞应激生物学国家重点实验室,福建 厦门 361102
北京时间2022年10月3日,瑞典生物学家、进化遗传学家斯万特·帕博(Svante Pääbo)成为最新的诺贝尔生理学或医学奖的获奖者。斯万特·帕博带领团队测序了已灭绝的古人类——尼安德特人(尼人)和丹尼索瓦人的基因组,发现这些已灭绝的古人类与现代人的祖先存在基因交流,非洲以外的现代人群都带有古人类的基因。现代人基因组中的古人类基因遗存有些是具有生理学意义的,例如,来自古人类的基因可能会影响现代人免疫系统面对感染时的反应。诺贝尔奖委员会表示通过揭示现代人与已灭绝的古人类在基因方面的差异,帕博的发现提供了探索人类独特性的基础。
古DNA研究是以分子生物学技术为基础发展起来的一个跨学科领域,通过结合考古学、历史学、语言学和古生物学等知识,分析古代生物的谱系、分子演化理论、人类的起源和迁徙、动植物的家养和驯化过程等。人类的DNA由30多亿对碱基组成,每一代人身上都会有一些碱基发生改变,叫基因突变,而有些突变会对一些性状产生影响。根据某些突变在人群中的分布和突变速率,我们可以了解人群扩张、缩减、分裂和融合等事件的历史情况,包括这些事件发生的大致时间[1]。研究古人类DNA可以帮助我们厘清人群之间的亲缘关系,以及具体基因的演化过程。这有助于我们了解现代人类一些表型特征的出现时间,进而推测当时的演化环境和自然选择条件,是探索人类演化背后动力和机制的重要一环。
研究几千甚至数万年前古人类的基因是一项充满挑战的任务。DNA极易随时间发生降解和化学修饰等现象,前者会使DNA片段变短,后者甚至会改变基因的碱基序列。另外,来自细菌和现代人类DNA的污染也是一个令人头疼的问题,有时污染的比例可能高达99.9%。帕博的部分研究与如何解决这些技术方面的问题有关[2-6]。他和团队制定了一系列的实验规范与流程,包括“超净室”的使用等。本文将从帕博对古DNA技术方法的探索、他的团队关于尼人和丹尼索瓦人的重要研究及发现、古基因组学的发展历程和最新进展等方面进行综述。
1 帕博与早期古DNA研究
斯万特·帕博1955年出生于瑞典的斯德哥尔摩,1997年成为新成立的马克斯·普朗克进化人类学研究所的5个负责人之一,并在那里一直工作至今。1985年,还是博士生的帕博在Nature上发表了一篇用分子克隆法研究埃及木乃伊DNA的文章。当时,30岁的帕博从一具2 400年前的木乃伊表层组织上提取到了一些古DNA片段。这些克隆出的古DNA片段长短不一,大部分只有不到500 bp(base pairs),但小部分超过了5 000 bp,克隆后甚至出现长达3.4 kb的片段[7]。根据我们今天对古DNA片段的了解,过长的片段很可能是由污染造成的[8-9],但这并不影响帕博的这次尝试对于古DNA研究的重要性。在这项研究中,帕博将分子克隆技术完整地应用到古DNA上,成为将DNA技术应用于考古人类学的先行者。这篇文章的最后,帕博还提出古DNA研究可以帮助人们回答人群或个体亲缘关系问题这一远见。
当时帕博使用的分子克隆法是20世纪70年代发展起来的一项技术,它可以帮助研究者分离和研究特定的基因片段[10]。不过这种DNA片段扩增的方法也有一些不足之处。分子克隆需要在活的组织内进行,且克隆结束后要对结果进行筛查才能获得最终需要的重组DNA。另外,这项技术需要特定的限制性内切酶将DNA切割成研究所需的片段。同时,完成一次分子克隆实验往往需要2~4天。
20世纪80年代发展出另一种扩增DNA片段的技术——聚合酶链式反应 (polymerase chain reaction, PCR)。至今这项技术仍在遗传学研究中起着至关重要的作用。样本保存情况不理想或样本量过少,都有可能导致没有足够量的DNA进行后续研究。PCR可以快速、大量地复制DNA片段,只需几纳克DNA就能实现扩增,在一定程度上降低了古DNA研究的门槛。PCR用控制温度来控制DNA的变性(双链变单链)和复性(单链变双链),并通过加入引物、聚合酶和脱氧核糖核酸实现DNA片段的不断复制[11]。这项技术的发明者Kary Mullis因此获得1993年诺贝尔化学奖。
帕博紧跟科学技术发展前沿,在1988年就利用PCR技术从7 000年前的脑组织中测序到古人类的线粒体DNA(mtDNA)片段[12]。帕博发现与现代人类的DNA不同,可以成功扩增的古DNA片段都不超过200 bp。在这篇文章中,帕博特别提到可以通过片段的长短来区分古人类和现代人类DNA,以及使用个体不同位置的组织进行实验等方法来控制古DNA实验中可能来自其他个体的污染。此后几年里,帕博和其他研究者将古DNA的研究方法应用到哺乳动物上,并在这些研究中不断完善以PCR为核心的古DNA研究技术[13-15]。
2 mtDNA研究
随着古DNA研究技术的不断成熟,越来越多研究者开始利用古人类和现代人类的DNA来研究人类起源和扩散问题。研究发现非洲人群拥有很多其他地区没有的mtDNA单倍群[16]。非洲人群mtDNA序列的多样性是非洲以外人群的两倍之多,这表明非洲人群可能有着更长的遗传历史[17-18]。根据DNA多态性和全线粒体DNA序列建立的谱系树也支持现代人类的晚近非洲起源这一观点[17,19]。Cann、Stoneking和Wilson在1987年发表在Nature上的文章表明现代人类的mtDNA可以追溯到约20万年前的一位非洲女性身上[20],这一发现后来被称为“夏娃理论”。说到现代人类的祖先就不得不提尼人,尼人是一群在30万~3万年前生活在欧洲和西亚的古人类。尼人与现代人的祖先生活过的时空是有重合的,相较于猿类,尼人与现代人是更晚才分化开的。研究尼人有助于我们了解现代人类在与猿类分离之后,最近的几十万年中演化出了哪些新的特征。对人类起源问题和考古发现的关注让帕博将注意力放到尼人身上。
帕博和他的团队在对比了现代人类和尼人的mtDNA序列后发现,现代人类之间在mtDNA高变I区(hypervariable region I)的碱基突变平均差异为(8±3)个,而现代人和尼人在这一区间的平均碱基突变差异达到(27±2)个[21]。虽然这个数字要远小于人类和黑猩猩之间的平均差异,但是也说明了尼人与现代人类的遗传距离较远。同时,遗传谱系树和基于形态学的观察也支持这一结论[22]。根据计算,现代人类最近的共同祖先出现在12万~15万年前[20],而现代人和尼人的共祖时间要早于50万年前[21,23]。这表明尼人不是现代人的一支,他们与现代人分离的时间要远早于现代人类内部分化的时间。后续的研究发现尼人之间的mtDNA序列十分相似,但是在约25 000年前的现代人mtDNA序列中找不到这些尼人的序列[24]。这进一步说明尼人没有与现代人进行大规模的融合,但不能完全排除少量融合的可能性。如果想完全排除融合的可能性就需要非常大量的古人类样本[25]。
对于mtDNA的研究开始只局限于控制区(D-loop),这一区域长度占mtDNA长度的不到7%[17],但是高通量测序的出现改变了这一局面。比起传统的Sanger末端终止测序法,基于大规模平行测序的第二代高通量测序法将DNA分子打断成更短的片段,可以通过边合成边测序的方法一次性对数百万个片段进行测序。高通量测序法也大大降低了实验成本,使得对尼人mtDNA全序列和核基因组测序成为可能。根据尼人mtDNA全序列得出的结论表明他们与现代人的分化时间约为66万年,与此前研究得出的结论基本一致[26]。在尼人mtDNA的13个编码蛋白质的基因中,研究者发现了更高的非同义突变比例,这可能与尼人的有效人口规模更小有关[26]。非同义突变多数是有害的。由于在规模较小的群体里遗传时的抽样误差更大,也就是说自然选择的效力更弱而遗传漂变的效力更强,因此轻微有害突变更有可能因随机事件被固定下来[27-29]。对5个尼人mtDNA全序列的进一步研究发现,他们的mtDNA多样性只有现代人类的三分之一,但非同义突变与同义突变比(0.168)约是现代人类(0.082)的两倍(图1)[30]。这也表明尼人可能长期有着较小的有效人口。帕博的实验室先是用高通量测序法测出尼人核基因组中100万对碱基的序列[23],为后续全基因组序列的获取打下良好的基础。未来对于来自更多时间与空间的尼人样本的研究可以帮助我们更好地认识这一问题。
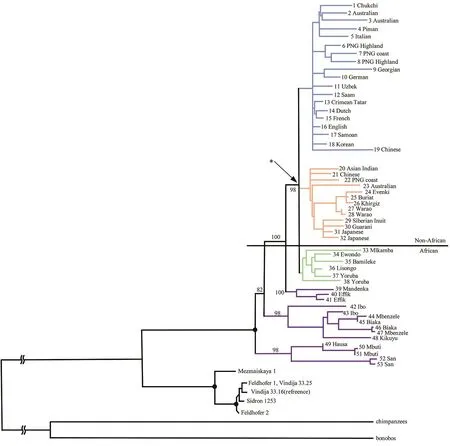
图1 现代人类、尼人和猿类的mtDNA进化树。箭头所指的位置为现代非洲人和非洲以外人群最近的共祖时间(MRCA)(根据[17]和[30]绘制)
一节来自西伯利亚阿尔泰山丹尼索瓦洞穴的指骨为研究者们提供了认识尼人以外古人类的契机。2010年帕博的团队从这节指骨上用DNA捕获技术提取到完整的mtDNA序列[31]。他们发现这些mtDNA序列不属于已知的任何古人类。这些古人类与现代人的mtDNA有385个核苷酸的差异,而现代人与尼人的mtDNA核苷酸差异只有202个[31]。这说明这些古人类与现代人的母系遗传距离要远于尼人与现代人。在后续的研究中,他们将此次新发现的古人类命名为丹尼索瓦人。根据尼人、现代人和丹尼索瓦人的mtDNA所构建的进化树显示,丹尼索瓦人与尼人和现代人有共同祖先[31]。丹尼索瓦人与现代人和尼人的共同祖先分化的时间在距今约100万年前,大约是现代人与尼人分化时间的两倍。研究者们也提到,mtDNA更易受到遗传漂变和正向选择的影响,因此全基因组的研究对于进一步明确丹尼索瓦人、尼人和现代人之间的关系及分化时间是十分有必要的[31-32]。考古学的证据还表明丹尼索瓦人与尼人和现代人曾经在3万~5万年前共同生活在阿尔泰山地区,这也使该地区成为研究现代人与古人类人群交流的重要地区[31]。
mtDNA的研究还被国内外学者应用在许多其他人群上。帕博等学者通过mtDNA探索了欧洲狩猎采集人群(H-Gs)和农业人群在新石器时期的变迁历史。通过对来自现代欧洲人的1 151个mtDNA序列的研究,他们发现与早期农业人群相关的H单倍群和与H-Gs相关的U单倍群[33]。分析结果显示,欧洲的H-Gs在15 000~10 000年前的这段时间呈增长趋势,而在10 000~5 000年前的时间段出现人口规模的缩减[33]。这一变化可能与欧洲农业的发展以及农业人口从9 000年前开始的扩张有关。但是从约4 000年前开始,携带H型和U型单倍群的人群又同时开始增长[33]。这可能与农业人群大规模地在欧洲扩张之后同化了各地的H-Gs有关。研究者们发现,现代欧洲人的mtDNA有约20%是来自于H-Gs的(图2)[33]。这也表明农业人群不是完全地替换了H-Gs,而是在一定程度上与他们发生了融合。

图2 现代欧洲人群及其祖先人群中各类mtDNA单倍群的频率占比(根据[33]绘制)
3 古人类全基因组研究
随着DNA测序技术和古DNA提取方法的不断进步,遗传学家研究的样本从mtDNA逐渐扩大到整个基因组。帕博的团队在2010年成功测序出尼人的全基因组[34],论文发表在Science上,该文章是帕博最重要的学术成果之一。根据尼人和不同地区现代人全基因组的对比,研究者们发现尼人和非洲人群的遗传距离要稍远于他们和非洲以外人群的遗传距离[33]。但是,尼人与欧洲人群和东亚人群的遗传距离是一样的,说明尼人与现代人类的接触可能发生在欧洲与亚洲人分化之前。通过对比不同地区现代人群、尼人和黑猩猩之间的遗传距离,研究者们认为大部分的基因交流都是从尼人到现代人的[33]。同时,该研究还算出非洲以外人群中来自尼人的基因占1%~4%[33],且这些基因交流发生在3.7万~8.6万年前[35]。在后续的研究中,帕博的团队发现,一群来自阿尔泰山的古老现代人类可能对同一地区的尼人有基因的渗入[36]。他们发现比起其他地方的尼人,阿尔泰地区尼人的第21号染色体与非洲人群的这一染色体共享更多等位基因。基于这些位点,研究者们认为在约11万年前,早期现代人类的一个分支对阿尔泰地区尼人有0.1%~2.1%的基因渗入[33]。对尼人全基因组的研究,证明现代人类和尼人是存在双向的基因交流的。这也说明将古DNA的研究范围扩大到全基因组是必要的,将使我们得到更多更全面的信息。
除了对尼人的研究以外,帕博的团队还对丹尼索瓦人的核基因组进行了测序。从全基因组的数据来看,丹尼索瓦人与尼人有更近的共同祖先,而且他们与现代人类的遗传距离和现代人类与尼人的遗传距离是差不多的[32]。这与基于mtDNA所得到的结论不完全一致[31,37]。这种不一致也许可以用不完全谱系分选来解释,即由于基因组中不同片段的进化速率和保守性不同,某些基因的多态性可能会被随机固定下来,因此通过不同片段构建的进化树之间可能会有差别。进一步研究发现,丹尼索瓦人在更新世晚期曾广泛分布于亚洲,在今天太平洋的美拉尼西亚人中有4%~6%的丹尼索瓦人基因[32]。在澳大利亚土著、新几内亚人、菲律宾南部的尼格利陀人群、波利尼西亚和东印度尼西亚的人群中都有发现来自于丹尼索瓦人的遗传混合[38]。在亚洲人群中可能同时存在属于尼人和丹尼索瓦人的基因片段,但是在欧洲几乎很难找到丹尼索瓦人的痕迹(图3)[39]。从距今34 000年前来自于蒙古东北部肯特省萨勒黑特(Salkhit)峡谷的古人类头盖骨提取到的基因组中,研究者们发现了约1.7%的来自尼人的基因,而来自丹尼索瓦人的基因只占0.127 5% (图4)[40]。研究还发现,在东亚地区古人类DNA中发现的丹尼索瓦人基因组片段与现代亚洲人群中的丹尼索瓦人的片段是高度重合的。然而在新几内亚人或澳大利亚土著中则找不到这些东亚古人类特有的丹尼索瓦人基因片段[40]。这说明丹尼索瓦人与现代人类至少有过两次独立的人群混合,分别发生在亚洲大陆和太平洋地区。
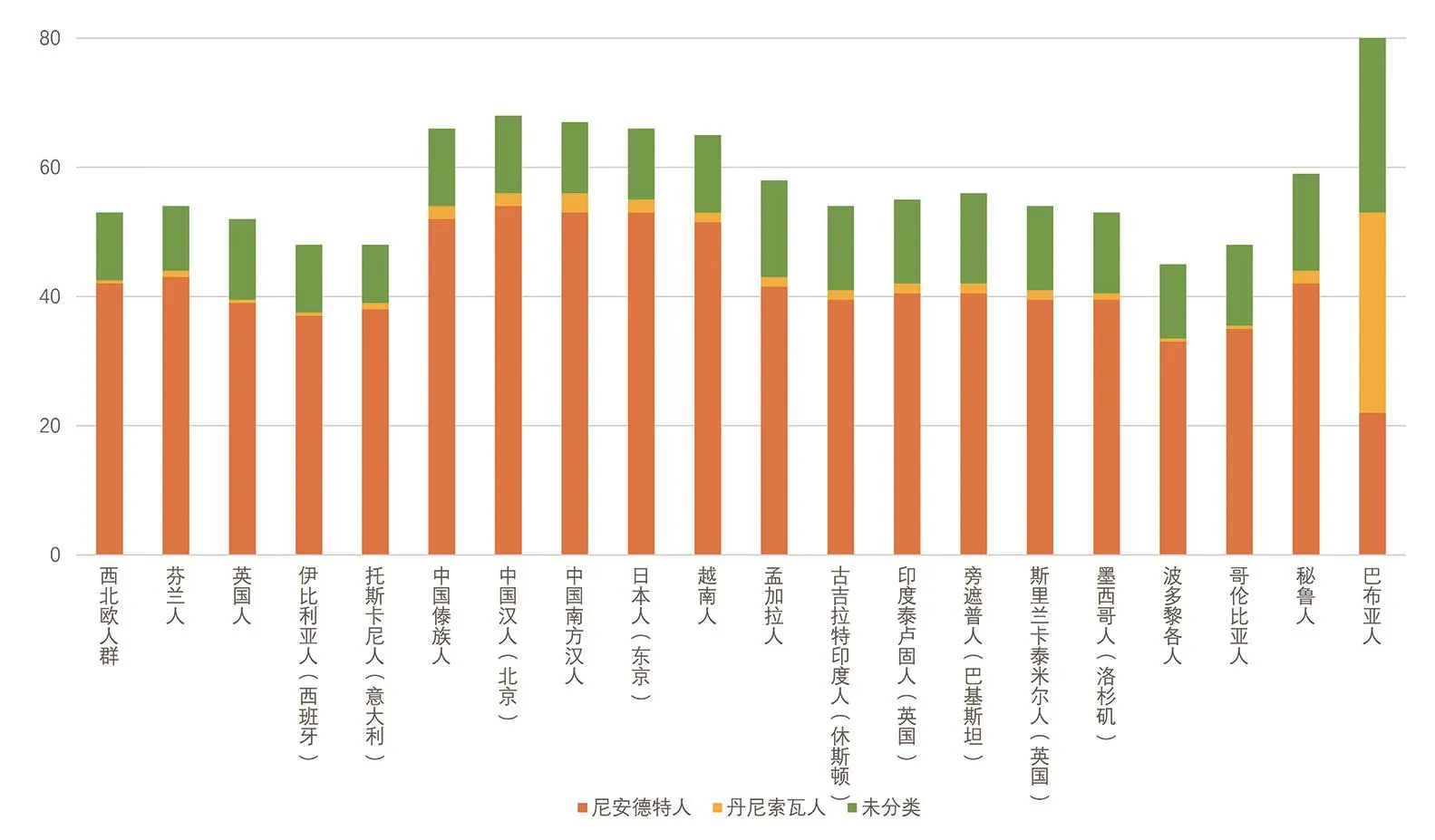
图3 不同地区人群平均每个个体中检测到的阿尔泰尼人和阿尔泰丹尼索瓦人基因片段的长度(单位:Mb)(根据[39]绘制)

图4 基于全基因组的现代人、尼人和丹尼索瓦人的进化树。红色箭头代表尼人向丹尼索瓦人的基因流;黄色箭头代表尼人向现代人的基因流;绿色箭头代表丹尼索瓦人向太平洋人群的基因流;蓝色箭头代表丹尼索瓦人向东亚人群的基因流(根据[32]和[36]绘制)
德国马克斯·普朗克所的Johannes Krause和哈佛医学院的David Reich团队通过古代人群全基因组的研究进一步明确了欧洲H-Gs和早期农业人群迁徙与融合的过程,并探索了现代欧洲人的祖先成分来源。他们找到了现代欧洲人的三个主要祖先人群,西欧的H-Gs、欧亚大陆北部的古西伯利亚人和来自近东的早期农业人群[41]。f4检验的结果显示,在近东的农业人群扩张到欧洲之后,来自西伯利亚地区的古代人群越来越多地与西欧人群发生融合。另外,f4检验还表明西欧的H-Gs没有和近东人群发生太多基因交流[41]。不同祖先人群对欧洲人的影响也与地域有一定关系。西欧H-Gs对北欧的影响更大,近东早期农业人群对南欧的影响更大(图5)。从整体来看,比起古西伯利亚人,欧洲人与欧洲西部的狩猎采集者有更多的基因交流。但是在近东早期农业人口中有约29%的古西伯利亚人基因,却没有西欧狩猎采集者的基因[41]。这说明古西伯利亚人基因可能以较小的占比更广泛地分布在当今的欧洲人群中。全基因组研究结果表明欧洲人的祖先成分是复杂的,不是简单地由农业人群替换了H-Gs。
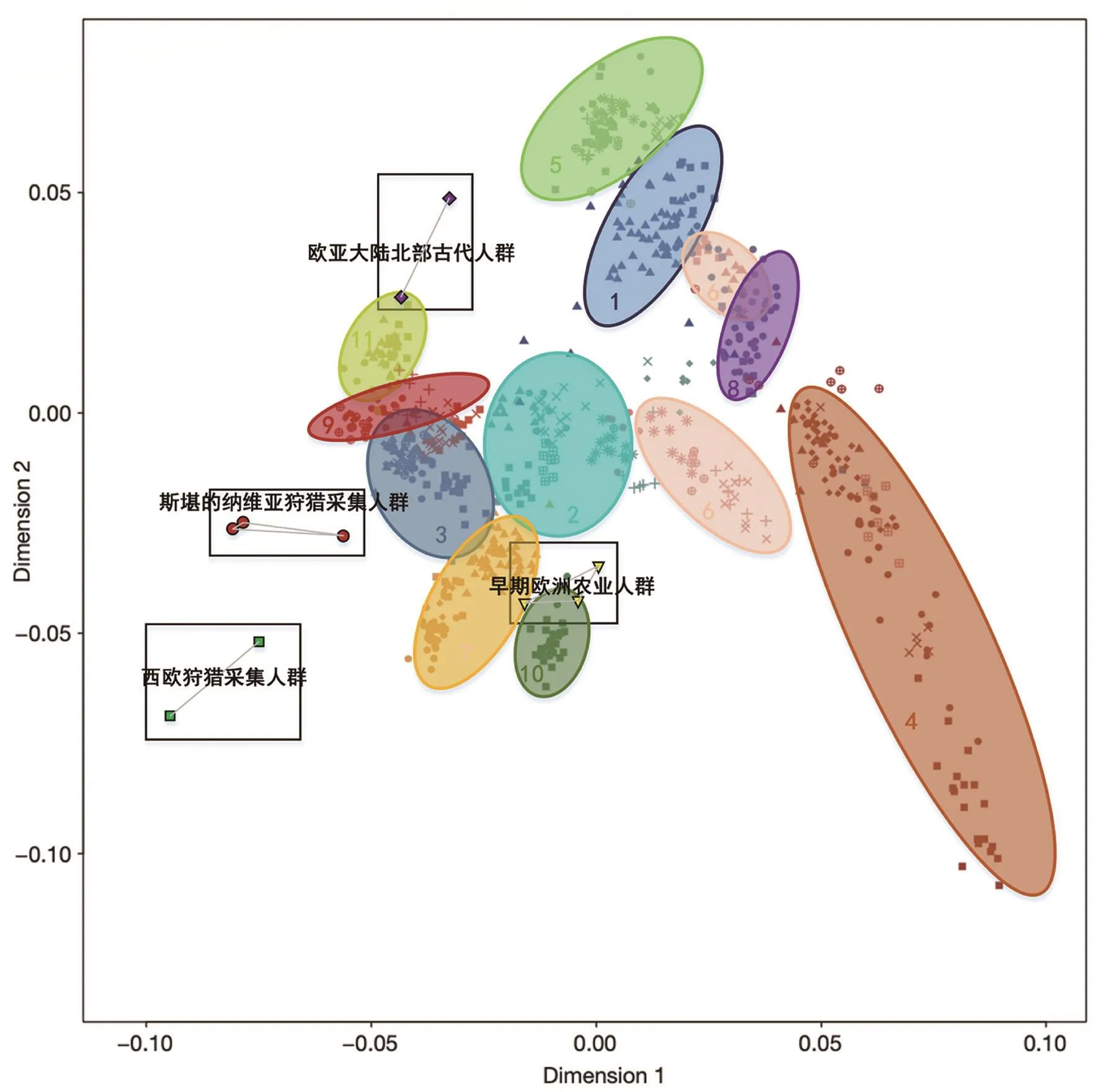
图5 PCA图。1~11号椭圆中的点代表不同地区的现代西欧人群,方框中的点代表不同地区的古人类样本。从PCA图中可以看出H-Gs与现代西欧人的距离较远。早期农业人群与西班牙(7和10号椭圆)、法国南部(7号椭圆)和地中海的意大利撒丁岛(10号椭圆)人群距离最近。古西伯利亚人群落在北欧和俄罗斯人群(11号椭圆)附近(根据[41]绘制)
基于全基因组的研究可以提供更精确的人群迁徙和融合的时间。近几年国内外学者还利用全基因组进一步解释了东亚和东南亚人群形成的过程,以及不同时期人群交流的情况。研究发现3 300~1 200年前的台湾地区人群有75%的祖先成分是与现代南岛语人群、壮侗语人群、南亚语人群以及长江流域的农业人群同源的,但另外25%的祖先成分则与中国北方黄河流域的早期农业人群有关[42]。在新石器晚期(4 600~4 200年前),东亚南部人群中就已经出现来自东亚北部人群的基因成分[43-44]。同时,福建与台湾地区之间的亮岛人被认为与南岛语人群、壮侗语人群、青铜器时代的东南亚人群以及南亚语人的祖先人群有关[42]。这为南岛语的祖先人群从中国华南地区扩散到台湾地区进而向东南亚扩散这一观点提供了证据支持。不过东南亚H-Gs的基因依然存在于现代东南亚人群中。这说明来自东亚的南岛语祖先人群在到达东南亚后与当地的原住民发生融合而不是直接替换了原有的人群。这一人群交流的过程与近东早期农业人群在欧洲扩散的过程有相似之处,即不同人群之间都倾向于融合而不是替换。另外,研究发现太平洋瓦努阿图地区的古代人群与新石器晚期的东亚南部人群的遗传距离比与新石器早期的更近[43-44]。因此可以推测,新石器晚期有更多南岛语祖先人群从东亚南部向外扩散[45]。进一步研究发现,来自1 800~2 300年前的印度尼西亚和菲律宾的古代个体可以被模拟为南岛和南亚语祖先人群的混合[45]。这表明南岛语人群到达台湾后,在距今2 000年前后到达了东南亚岛屿地区。
4 Y染色体研究
古DNA研究是从mtDNA开始的,而对于Y染色体的探索开始得晚一些。1995年,帕博在Science杂志上发表了一篇关于Y染色体的综述文章[46],在这篇文章中他对mtDNA和Y染色体进行了初步的比较。mtDNA在人体中有较多的拷贝数,具有适中的突变率,不重组,且只能由女性遗传给后代,因此很适合用来研究群体间的母系亲缘关系[17]。由于mtDNA的多样性更容易受到遗传漂变、自然选择、人口规模变化等原因的影响[32],研究者们渐渐意识到只依靠mtDNA追溯人类的起源和发展还是有局限性的。对于DNA不同区域的研究可以帮助人们更全面地了解人类进化的相关问题,于是很多研究者开始将目光投向Y染色体[47-48]。Y染色体比mtDNA结构更复杂,片段更长,也有更丰富的多态性,而且不同位点的突变速率是不同的[49]。在同一人群中,由于婚配制度和遗传方式的限制,Y染色体和mtDNA的多样性可能存在差异[50-51]。根据多态性可以区分出Y染色体上特定基因或区域的不同单倍型(haplotype),进而追踪不同人群的父系祖先来源[52]。
由于高质量男性尼人样本的缺乏,关于尼人和丹尼索瓦人的Y染色体的研究直到近几年才取得突破。对3个尼人和2个丹尼索瓦人的Y染色体研究的结果显示,丹尼索瓦人与尼人和现代人的父系共同祖先分化的时间在距今70万年前,而实验中的3个尼人与现代人Y染色体的分化时间在37万年前[53]。对比基于mtDNA、Y染色体和全基因组所构建的谱系树,研究者们发现丹尼索瓦人与尼人和现代人遗传关系的不一致可能是由于晚期尼人的线粒体和Y染色体被早期现代人的一个分支人群替换了[53]。因此,丹尼索瓦人与现代人的分化时间在三个谱系树中是相对一致的,而根据Y染色体和mtDNA建立的谱系树会显示晚期尼人与现代人的距离更近。在阿尔泰地区发现的有现代人类基因渗入的尼人为这一论点提供了有力的支持[36]。关于尼人的最新研究发现,尼人Y染色体的多样性要远小于mtDNA的,而且被研究的13个尼人的Y染色体共祖时间也远小于mtDNA的[54]。这可能是因为尼人女性在不同群体间的迁徙更频繁。根据Y染色体和mtDNA所建立的模型显示,尼人群体的平均人口为20人,其中有60%~100%的女性来自其他群体[54]。通过对古人类基因的深入探索和先进分析方法的应用,研究者们可以对古人类从人群交流到社会结构的多个维度进行探索。
5 古DNA与现代人类特征的演化
除了研究人类起源和不同人群之间亲缘关系与迁徙历史等问题,古DNA还可以通过研究某些基因突变或者渗入的时间,来推测现代人类某些特征出现的时间和背景。帕博等[34]通过对比现代人类和尼人的全基因组,找到了现代人类中212个发生了核苷酸多态性选择性清除(selective sweep)的区域。这些区域往往与自然选择中对基因的正向选择有关。选择的作用越强烈,发生选择性清除的区域可能就会越大。在该项研究中找到的最大区域包含THADA基因,这个基因附近的单核苷酸多态性(SNP)位点与2型糖尿病有关。另外,他们还发现一个与颅骨锁骨发育不良有关的基因RUNX2 (CBFA1)。研究者们认为这个基因的突变与现代人和包括尼人在内的其他古人类在形态上的一些差别有关[34]。RUNX2基因与颅骨缝的延迟闭合、锁骨的发育不良、牙齿畸形和钟形胸廓有关[55]。其中,钟形胸廓是一个在尼人和其他智人身上很典型的特征。现代人类在这些形态上的变化很可能为其提供了某种进化方面的优势。对于人类基因正向选择历史的探索可以帮助我们更好地了解现代人类重要特征进化的历程,以及现代人类是如何应对自然选择的压力的。
现代人中一些尼人和丹尼索瓦人的基因由于有利于人类对环境的适应,从而被保留下来。研究发现,与免疫相关的STAT2基因在欧亚人群中的一类单倍型(N)与尼人的高度相似,且在撒哈拉以南的非洲人群中没有出现[56]。这类单倍型在欧亚人群中的平均占比为5%,但在太平洋的美拉尼西亚人中的出现频率高达54%[56]。美拉尼西亚人中并没有更多尼人祖先成分,因此带有单倍型的免疫基因STAT2可能在美拉尼西亚地区受到正向选择。丹尼索瓦人的基因也对现代人的免疫系统有一定的积极影响。与人类白细胞抗原相关的一个等位基因HLA-B*73就是由于丹尼索瓦人在西亚的基因渗透,进而扩散到整个欧亚地区的[57]。这个等位基因可以编码许多独特的自然杀伤细胞受体的配体,对自然杀伤细胞的运转有着重要作用。自然杀伤细胞是一类与人体抗病毒抗感染等免疫调节功能有关的重要免疫细胞。HLA-B*73在调节免疫系统运作方面的优势使它在欧亚人群中的占比超过50%[57]。除了免疫系统外,丹尼索瓦人的基因渗透还对高海拔的青藏高原人群适应低氧环境有一定贡献。EPAS1基因是一个与个体对低氧环境反应相关的基因,它通过在低氧情况下调节血红蛋白的浓度来降低个体高原反应等心血管疾病的风险[58]。研究发现,青藏高原人群特有的更利于低氧环境的EPAS1基因单倍型大多存在于藏族人中,且与丹尼索瓦人的EPAS1基因单倍型十分相似,而这一单倍型在包括汉族人在内的世界其他族群中是很罕见的[59],这说明藏族人的EPAS1基因更有可能是来自丹尼索瓦人而不是现代人群。
帕博和Zeberg还发现了一些可以增加或减少新型冠状病毒感染后重症概率的相关基因,而这些基因与尼人有关。他们先是发现在3号染色体上的与感染新型冠状病毒后呼吸衰竭有关的基因[60],这组基因被认为是新型冠状病毒重症的主要风险因素之一。涵盖这组基因的片段在南亚人群中的占比为50%,在欧洲人群中为16%(图6)。而这个基因片段的序列与尼人基因中对应区域的序列十分相似,因此推断这一新型冠状病毒重症的风险因素可能来自尼人。在后续的研究中,他们又在12号染色体上发现一个与降低新型冠状病毒感染后重症风险有关的单倍型[61]。这个长度为75 kb的片段会在个体受到病毒感染时编码一些有重要作用的蛋白质,在非洲以外地区都有较高的出现频率(图6)。研究者们在尼人基因相对应的区域中找到与这个单倍型一致的片段,因此现代人很有可能是从尼人的基因中获得这一单倍型的;而且,这个片段长达75 kb,说明它不太可能来自于现代人类和尼人的共同祖先[61]。研究结果表明,尼人对现代人类的基因渗透既有积极也有消极的影响,但是基因的功能大多不是单一的,因此不能只从一个方面去判断一个基因的利弊。

图6 与冠状病毒重症风险相关的单倍型的频率分布地图。蓝色代表与重症风险增加有关的单倍型,红色代表与重症风险降低有关的单倍型(根据[60]和[61]绘制)
通过对有语言障碍的家族和个体的探索,研究者们发现在FOXP2基因上发生的突变与语言能力的发展息息相关。FOXP2基因的缺陷会影响大脑语言相关区域的发育,从而影响口面部的活动能力和语言的表达与认知能力[62]。FOXP2基因负责编码一个由715个氨基酸组成的蛋白质。人类的FOXP2蛋白质与老鼠的只有3个氨基酸的差别。黑猩猩和大猩猩的FOXP2蛋白质与老鼠的只有1个氨基酸的差异,而与现代人类有2个位点的差异[63]。黑猩猩与老鼠的FOXP2蛋白质结构基本一致,而人类的FOXP2蛋白质与前两者在结构和功能上是有区别的[63]。这说明人类FOXP2基因的关键突变更可能发生在人类与猿类进化分离以后,而且不同地区现代人的FOXP2蛋白质是一样的,不存在多态性[63],这说明FOXP2在现代人类中已经经过正向选择被固定下来。为了进一步验证正向选择对现代人类FOXP2基因的作用,研究人员分别测序了现代人类和黑猩猩该基因上非编码区域的14 063组碱基对。他们发现一些在黑猩猩中频率很低的等位基因在现代人类中有着较高的频率[63]。这说明在现代人类中这些非编码区域的多样性由于编码区域被正向选择而受到了选择性清除。根据似然法建立的模型显示,现代人类FOXP2的固定可能发生在距今20万年前,这一时间点可以对应到目前已知的现代人类祖先人群走出非洲向欧亚扩张的时间点之一[63]。人类语言的进化可能为人类的扩张提供了有利的条件。
帕博对尼人FOXP2基因的研究发现,尼人和现代人在这个基因上的位点突变是一样的。同时,这些突变附近由于正向选择所发生的选择性清除的信号也是一致的。因此,现代人类FOXP2基因出现的时间有可能比20万年前更早,在尼人与现代人分化之前[64]。也有研究显示尼人与现代人一致的FOXP2基因可能是由于早期现代人类向尼人的基因渗入而导致的[36]。随着研究的不断深入,对于这一问题的认识会不断更新。在探索现代人类特征对应的基因突变是在何时产生的这类问题时,尼人和丹尼索瓦人这样的古人类就像人类漫长演化历史中的坐标。通过寻找这些坐标并进行比对,现代人类进化历程的时间轴将得到更精确的矫正。
6 结语
斯万特·帕博通过30多年在古DNA领域的探索,实现了从mtDNA到全基因组的测序。他对于古人类研究的专注与奉献为我们打开了一扇了解人类进化历程的窗户。同时,帕博对前沿技术的敏锐洞察力,也使他一直处于古DNA技术革新的前沿。研究像尼人和丹尼索瓦人这样的已灭绝古人类,可以深化我们对于现代人类演化过程的了解。虽然帕博的研究重点集中在尼人和丹尼索瓦人上,但是他所开创或优化的古DNA研究方法使得许多研究者受益,从而使越来越多的人加入到了古基因组学的行列。研究古基因组不仅可以回答有关人群起源与扩张的问题,还可以回答与现代人类特征的进化有关的问题。依托先进的实验与分析技术,可供研究的古DNA范围不断扩大,可以回答的问题将不断细化与深化。
猜你喜欢
科学大众(2022年23期)2023-01-30
河北地质大学学报(2021年5期)2021-11-08
中国自行车(2017年5期)2017-06-24
飞碟探索(2016年8期)2016-09-06
大众考古(2015年5期)2015-06-26
分忧(2015年6期)2015-06-09
故事会(2006年3期)2006-01-12
