
丛枝菌根真菌与植物共生影响植物水分状态的研究进展①
2022-12-09 01:04蒲子天王鑫鑫
土壤 2022年5期
蒲子天,张 林,张 弛,王 红,王鑫鑫,6*
丛枝菌根真菌与植物共生影响植物水分状态的研究进展①
蒲子天1,张 林2,张 弛3,4,5,王 红3,4,5,王鑫鑫3,4,5,6*
(1 河北农业大学资源与环境科学学院,河北保定 071001;2 中国农业大学资源与环境学院,北京 100193;3 国家北方山区农业工程技术研究中心,河北保定 071001;4 河北省山区农业技术创新中心,河北保定 071001;5 河北农业大学河北省山区研究所,河北保定 071001;6 农业农村部华北节水农业重点实验室,河北保定 071001)
丛枝菌根真菌(arbuscular mycorrhizal fungi,AMF)能与宿主植物形成共生体,广泛存在于陆地生态系统中。大量研究表明,不同水分条件下,植物通过接种AMF比未接种AMF的植物具有更强的水分吸收能力和更高的水分利用效率。同时在干旱胁迫与盐胁迫下,接种AMF能有效提高宿主植物的耐旱性与耐盐性。本文综述了不同水分条件下,AMF与植物共生通过扩大植物根系吸收面积、改善根系结构,增强植物根系吸收水分能力的相关研究进展。土壤中根外菌丝网络的形成,不但为植物增加了水分吸收途径(菌根途径),还通过改善植物体内的矿质营养来调节植物对水分的吸收,进而影响植物的水分吸收状况;不同水分条件下,根系被AMF侵染后植物的光合作用、蒸腾作用以及气孔导度都得到增强,植物蒸腾作用的增强能够直接有效地提升植物的蒸腾拉力,因此植物对水分的吸收能力得以提升。同时,被AMF侵染的植物的水分利用率、蒸腾速率以及净光合速率得以提升从而提高了植物的水分利用能力。进一步总结了缺水胁迫(干旱胁迫、盐胁迫)严重影响植物体内的水分状况,通过接种AMF可以有效调节植物在缺水胁迫下植物体内渗透调节物质的含量、抗氧化酶的活性,平衡植物体内离子平衡,提升植物光合、蒸腾作用水平,从而提高植物的耐胁迫能力。本文通过综述不同水分条件下,AMF影响植物水分状况和抗盐性机制的研究,以期为菌根互作对植物水分状况的改善提供参考。
丛枝菌根真菌;根系;盐胁迫;干旱胁迫;水分状况
土壤水资源短缺随之产生的土地问题便是土地干旱与土壤次生盐渍化。如何提高植物的耐旱性与耐盐性成为当前的热点问题,而AMF与根系相互作用对植物水分的获取和利用成为提高植物抗旱性与抗盐性方面近年来的研究热点。干旱与盐胁迫条件下,AMF对植物的部分影响机制类似:一方面AMF均能通过增强植物的根系结构,提高植物根系吸水动力进而增强植物根系对水分的吸收能力;另一方面接种AMF的植物通过调节细胞内的渗透调节物质含量,提高抗氧化酶活性以及提升植物的水分利用率[8-9]。因此将AMF在干旱与盐胁迫下对植物的影响进行对比,能够更好地总结出AMF对不同土壤环境中植物影响的共性。
多年生植物、一年生植物与AMF共生之间的差异与宿主植物效益积累的滞后性相一致[10]。为了建立与AMF共生的联系,植物往往需要在生长的早期阶段投入更多的资源。多年生植物在第一个生长季投资生长,使氮、磷能够将互补资源再投资于植物。然而,一年生植物在第一个生长季将资源从生长转移到繁殖,因此无法长时间来实现协同生长,这使得互补共生体生长的好处受到时间延迟的影响[11]。与外生菌根真菌相比,AMF能够更好地缓解植物面临的水和营养缺乏问题[12]。AMF的根外菌丝相当于根系增加了吸收面积,增强了对水分、矿物营养的吸收能力(特别是对于磷的吸收),进而促进植物生长、增加植物的耐干旱能力等。Primieri等[13]通过Meta研究表明,与一年生植物相比较,多年生植物虽然依赖外生菌根真菌对养分、水分的吸收,但也依赖AMF对资源的吸收。
本文主要对近年来在不同水分条件和土壤盐含量下,AMF对不同类型植物(多年生木本植物和一年生作物)根系结构和吸水动力影响以及植物应对干旱胁迫与盐胁迫时的体内组织抵抗胁迫机制变化的研究进行了总结。以期为不同水分条件和土壤盐含量下AMF影响植物水分吸收和利用的研究提供参考。
1 AMF对植株根系结构的影响
1.1 增强水分吸收特征
根是植物吸收水分的主要器官,植物自身根系特征增强能够提升植物对水分吸收的能力。不同水分条件下,植物接种AMF后,其根体积、根长度、根吸收面积等根系特征增强,从而改善植物的吸水能力。研究表明正常水分供给,植物接种AMF后根系结构发生明显变化,其中主根长、侧根长、侧根数等根系结构增强并且根系体积和吸收面积均有所增大[14-16]。例如,正常水分条件下,大青杨苗木()接种摩西球囊霉(,F.m.)、根内球囊霉(,R.i.)、弯丝球囊霉(,G.s.)和地表球囊霉(,G.v.)4种AMF菌剂,杨青苗木根系的吸收面积和活跃吸收面积均明显扩宽,其中接种R.i.的效果最好,苗木根系总吸收面积扩大56.2%,活跃吸收面积增加150.2%[17]。除多年生木本植物外,辣椒(L.)通过接种幼套球囊霉(,G.e.)、F.m. 和G.v. 3种AMF,在生长的苗期、始花始果期、盛花盛果期、衰老期4个时期均表现出根系体积、根表面积以及一级侧根数等根系特征增强,相较于其他两种AMF,F.m.也使辣椒根系特征增幅更加明显[18]。
干旱或盐胁迫下,AMF通过促进植物根长增加、根毛密度增大、根体积和根表面积扩大等方式提高根系对水分的吸收能力,来缓解干旱或盐胁迫对植物造成的危害[19-20]。例如,在3种不同水分环境(正常水分、中度干旱、重度干旱)中对荆条(L.)接种F.m.,其根长、根体积、根表面积以及根尖数均显著提高,但根的平均密度却低于非菌根荆条。重度干旱条件下,F.m. 对荆条根系的影响最为显著,其中AMF促进荆条根系伸长高达131.1%[21]。同时,AMF能使柳枝稷(L.)根系结构增强,并随着水分胁迫的加强,AMF对根系的增强效果愈加明显。当土壤含水率仅有20% 时,AMF对柳枝稷根系的增强效果最为显著[22]。有研究发现在不同程度的盐分胁迫下,对白三叶(L.)接种R.i.和F.m. 两种AMF均能够使得根系总长度、总体积、总面积以及根尖数增加,其中两种AMF混合接种的效果最好,根尖数提升117.7%,总长度提升54.3%[23]。并且盐胁迫下,尖瓣海莲(L.)以及沙枣(L.)接种AMF,通过提升植株的地下部生物量来缓解盐胁迫带来的危害[24-25]。
上述研究表明,不同水分条件下,不同种类的AMF能够通过改善不同类型植物(多年生木本植物与一年生作物)的根长、根体积、根表面积等根系特征,增强植物根系的水分吸收能力。但不同水分条件下,不同种类AMF对植物根系的改善效果并不相同,如何在不同水分条件下快速确定对植物调节效果最优的菌种,对未来发展尤为重要。
1.2 菌丝直接或间接增强植物水分吸收
根内AMF菌丝生长在植物根皮层并形成致密的菌丝网络,最终成为真菌和植物之间营养物质和代谢物的相互交换以及水分吸收的通道[26]。除根内菌丝外,根外菌丝通过在土壤中的延伸,扩大了根系对营养物质和水分的吸收范围[27]。有研究表明,庞大的菌丝网络,能够为植物吸收水分[28]。Ruth等人[29]的研究结果表明,菌丝的直接和间接吸水对植物总吸水率的贡献约为20%。并且不同的水分条件根外菌丝在水分吸收的过程中,不会影响植物自根系从土壤中吸收水分的速率,这更加表明了菌丝的形成能够提高植物的水分吸收速率[30]。同时菌根菌丝直径小(2 ~ 5 μm),既能更好地接触土壤微域(直径10 ~ 20 μm)中的土壤水,又能为菌丝与水膜提供更好的接触,从而吸收到植物根无法触及的水分[31]。另外根外菌丝形成活跃的吸收水分表面,因此菌丝从水势较低的土壤中吸收水分的能力更强[32]。
AMF菌丝可为植物提供矿质养分,植物通过菌丝吸收的养分甚至可高达100%,植物养分吸收的提高可以缓解植物的干旱胁迫[33]。例如,葡萄(L.)扦插苗接种AMF后,植株体内氮、磷、钾含量均明显提升,提高了水分吸收能力[34]。除了AMF的营养功能外,被AMF侵染的植物还显示出更好的水分利用效率[35]。与未接种AMF的植物相比,被AMF侵染的植物能够通过自身根系直接吸水和根外菌丝间接吸水两种方式进行水分吸收[36-37]。
这是沈家煊(1995)的认知语法学观点[5]。在时间轴上,“买”“(动作)的起点是“相中某种商品”,终点是“付钱买回完成商品交易行为”,为有界的动作。“吃亏、上当”是贬义词,也是有界的,两个有界的部分能够互相匹配,所以能说“你买不了吃亏,你买不了上当”。但我们不禁再问:为什么“买”所表示的行为动作时有界的,就必须要求其所带的宾语也是有界的?同是有界的“伤心、失望”为何不能进入此格式?既然同是有界的,为何其所带的宾语有句法搭配上的限制(后多带体词性宾语)?看来这样的解释仍然有缺陷。
上述研究表明,AMF的根外菌丝对于水分的吸收有着直接或者间接的作用。但是目前对水的同位素研究较为缺失,菌丝吸收水分和根系吸收水分如何区分还需要更为精细的研究。
2 AMF对植物根系吸水动力的影响
根系吸水主要取决于两种动力:横向根压与纵向蒸腾力。蒸腾拉力是植物根系吸水的主要驱动力,植物蒸腾作用的增强能够直接有效地提升植物的蒸腾拉力。有研究表明,正常水分条件下对水曲柳(R.)接种F.m. 和G.e. 两种AMF均能够提升植株的光合、蒸腾速率并有效调节叶片的气孔导度,并且当F.m. 和G.e. 混合接种时效果最好;与未接种AMF相比,菌根苗木生长的速生期,光合速率提升43%,蒸腾速率提升86%,气孔导度提升高达104%[38]。
除了直接增强植物蒸腾作用外,AMF还可通过抑制植物的叶片气孔阻力,增强气孔蒸腾作用,提升植物根系的蒸腾拉力。如,韩冰等[39]对黄瓜(L)施入(GMA)、(GMS)、(GMH)及(G.v.)4种不同的AMF菌剂后,黄瓜的气孔限制值均显著降低,且光合作用、蒸腾作用以及气孔导度均明显增强,其中GMA的提升效果良好,但GMH对水分利用率的提高最为显著。
干旱条件下随着土壤含水量的降低会限制植物的水分吸收能力[40]。研究表明,干旱胁迫下接种AMF能够调节植物的气孔导度、蒸腾速率以及光合作用,极大改善了植物的纵向蒸腾力,从而提高植物的水分吸收能力。例如,Augé等[41]发现,干旱条件下接种AMF的植物其碳交换率能够增加49%,叶片的气孔导度增加28%,蒸腾速率提升26%。与正常供水相比,干旱条件下植物从AMF获得的效益往往更多,干旱条件下AMF对植物光合作用的影响大于正常供水,而并未对气孔导度和蒸腾作用产生明显影响,使得干旱条件下接种AMF的植物的净光合效应大于正常水分条件。如,三叶鬼针草(L.)在正常水分条件下接种F.m.,植株的光合速率、蒸腾速率以及气孔导度都有明显的提升,但在重度干旱条件下接种F.m. 只对植株的光合速率有较为明显的提升,而蒸腾速率与气孔导度却没有明显的提升作用[42]。干旱条件下,由于接种AMF的植物净光合速率更高,因此接种AMF的植物往往比未接种的植物拥有更高的生物量[43]。
盐胁迫下,接种AMF植物的光合作用、蒸腾作用以及气孔导度均增强,并且有更高的水分利用效率(光合作用和蒸腾作用的比率)[44]。如AMF对桢楠(B.)净光合速率、气孔导度、蒸腾速率以及胞间CO2浓度的增强作用随着盐分的增加而更加显著[45]。盐胁迫使得桑树(L.)体内净光合速率、气孔导度、蒸腾速率降低,但接种AMF的植株能够有效缓解这种抑制效应,并且随着土壤盐分的增加其缓解效果也更为明显[46]。
上述研究表明,不同水分条件下AMF通过直接增强植物蒸腾作用以及抑制植物的叶片气孔阻力两种方式提升植物的纵向蒸腾力,增加植物的根系吸水动力。但AMF如何增强植物的蒸腾速率以及降低叶片的气孔阻力方面的研究较少,并且AMF对哪种提升根系吸水动力的方式影响更为剧烈也未有体现。
3 缺水胁迫下AMF对宿主植物的影响
3.1 干旱胁迫下AMF对宿主植物的影响
干旱胁迫使植物萎蔫并影响植物的生理生化功能,严重阻碍了植物的生长。植物通过改变其生理、形态特性来缓解干旱胁迫的危害。例如,干旱胁迫下,刺槐(L.)会降低株高、地上部干重以及地下部干重来对抗干旱胁迫所带来的负面影响[47]。除自身变化外,AMF对于植物在干旱胁迫下减少干旱损伤是非常有益的,接种AMF的植物抗干旱能力更强,这种现象体现在可溶性蛋白、可溶性氨基酸等渗透调节物质的含量得以调节,植物体内抗氧化酶活性的增强。干旱胁迫下,接种AMF后,植物通过改善根系特征,增强光合、蒸腾作用,调节气孔行为,调节植物体内渗透调节物质的含量,提高植物组织的抗氧化能力等方式提高抗旱能力。由上述总结AMF提高宿主植物抗旱性部分机制的模式图(图 1)。
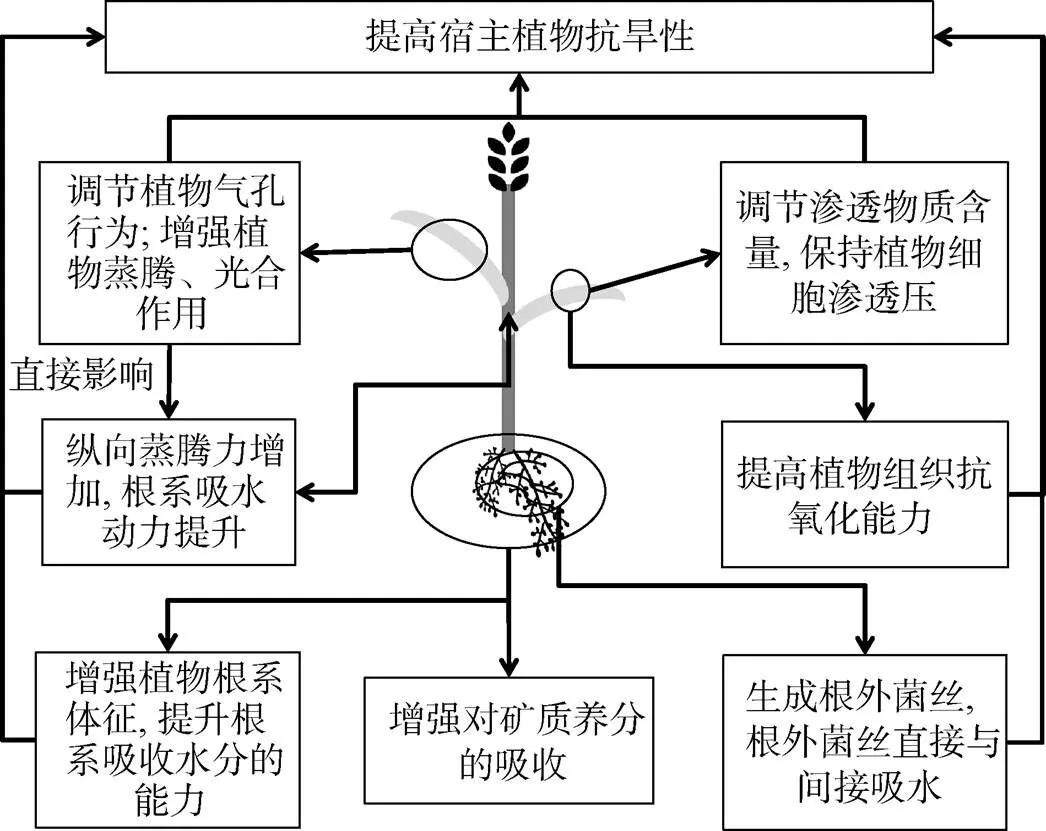
图1 AMF提高宿主植物抗旱性部分机制的模式图
3.1.1 改善植物的渗透调节 植物细胞在面对土壤水分缺乏时,一般通过调节胞内可溶性糖、可溶性氨基酸等物质的含量保持细胞渗透压,维持植物细胞渗透势,从而调节植物细胞对水分的吸收状况。干旱条件下AMF对植物体内脯氨酸含量的变化研究相对较多,部分观点认为接种AMF提升植物体内脯氨酸含量以应对干旱胁迫对植物细胞所造成的渗透胁迫,因此植物体内脯氨酸含量升高。如,岳海等[48]通过对澳洲坚果幼苗(F.)进行水分胁迫试验(土壤含水量为9% ~ 10%),发现接种F.m. 和土著AMF两种菌剂后,其植物体内脯氨酸含量均有明显提升,并且,澳洲坚果幼苗的根系以及排根分泌的脯氨酸含量也均有明显提高。也有观点认为,干旱条件下接种AMF有效调节了植物的水分状况,所受干旱胁迫影响得以缓解,使得植物的脯氨酸水平降低[49]。此外,不同程度的干旱水平也是造成脯氨酸水平不同变化的原因,例如,木棉幼苗(L.)在中等干旱胁迫下接种AMF植株叶片的脯氨酸含量小于未接种植株叶片的脯氨酸含量;但在重度干旱胁迫下,接种AMF的植株叶片的脯氨酸含量远远高于未接种植株叶片的含量,可以推测接种AMF的植物会根据不同的干旱程度对植物造成的影响来调节植物体内脯氨酸的含量[50]。
可溶性糖是植物体内重要的渗透调节物质,主要包括蔗糖、肌醇等。干旱胁迫下,植物体内可溶性糖类含量明显提升,通过接种AMF能够有效调节植物体内可溶性糖含量,从而提高植物耐旱性。如,干旱胁迫下,对紫穗槐()接种F.m. 和缩球囊霉(,F.c.)两种AMF,与未接种植物相比,接种AMF的植物体内可溶性糖含量提升更加明显,并且当土壤相对含水量为30% 时,植物体内可溶性糖的含量最高[51]。
3.1.2 提高植物抗氧化能力 干旱胁迫下,植物体内会产生过量的活性氧(ROS)来抵抗干旱胁迫对植物产生的危害,但过量的ROS会对植物组织产生氧化损害[52]。ROS一般留存在叶绿体和线粒体等细胞器中,过量的ROS会影响细胞膜系统的结构与功能,因此往往会提升丙二醇(MDA)的含量[53-54]。研究表明,干旱胁迫下AMF可以通过增强植物体内抗氧化酶的活性,降低植物细胞内MDA的含量,提高植物的抗氧化能力,从而提升植物的耐旱性[55]。如,干旱胁迫下,对菊苣(L.)接种R.i.,植物体内超氧化物歧化酶(SOD)、过氧化物酶(POD)、谷胱甘肽过氧化物酶(GPX)和抗坏血酸过氧化物酶(APX)等抗氧化酶活性明显增强,并且植物细胞内MDA以及过氧化氢(H2O2)的含量减少[56]。
3.2 盐胁迫下AMF对宿主植物的影响
植物的盐胁迫是由于植物生长在高盐度生境受到高渗透势影响所产生。因此,盐胁迫首先造成植物体内Na+过量累积,破坏了植物细胞内Na+、K+平衡;另外,植物体内水分的大量外流,对植物细胞造成了渗透胁迫;与干旱胁迫相同,盐胁迫也会引起植物细胞内ROS的过量产生,从而对植物产生氧化胁迫。因此水分胁迫(植物生理干旱)也可能是由高浓度盐(盐度胁迫)引起的。Chandrasekaran等人[57]通过Meta分析发现,盐胁迫下接种AMF能够促进植株对氮、磷、钾等营养元素的吸收,并显著降低对Na+的吸收来缓解盐胁迫带来的离子胁迫;其植株体内抗氧化酶、超氧化物歧化酶、过氧化氢酶、过氧化物酶和抗坏血酸过氧化物酶的活性显著升高能够缓解植株的氧化胁迫。同时,有研究表明,AMF缓解盐胁迫导致植物生长的减弱机制与植物应对干旱的机制相似[58]。
3.2.1 改善植物的渗透调节 盐胁迫下接种AMF同样能够调节植物渗透调节物质的含量来减弱胁迫带来的影响。例如,不同程度的盐胁迫下(0、50、100、150、200 mmol/L NaCl),紫花苜蓿(cv.)接种G.e.、F.m. 和R.i. 3种AMF,在不同盐浓度下均为接种F.m. 效果最好,植物体内可溶性糖的含量升高,脯氨酸的含量降低,超氧化物歧化酶(SOD)与过氧化物酶(POD)的活性显著增强,所受盐胁迫影响得到有效缓解[59]。同时,盐胁迫下尖瓣海莲(L.)通过体内脯氨酸含量升高来保持植物细胞渗透压,防止细胞脱水。接种F.m.、R.i. 后,尖瓣海莲体内脯氨酸含量与同盐度下未接种AMF的植物相比有了明显的降低,因此表明AMF可以有效缓解盐迫害对尖瓣海莲的影响[60]。
3.2.2 提高植物抗氧化能力 盐胁迫下AMF能够提高植物体内抗氧化酶的活性,缓解氧化损伤,提高植物抗盐性[61-63]。Pan等人[64]通过Meta分析进一步证实,盐胁迫下AMF可以通过增强植物的SOD、CAT活性,降低MDA的含量以及调节植物体内脯氨酸的含量来缓解盐胁迫带来的危害。有研究表明,接种AMF对刺槐叶片氧化损伤的缓解作用比根系更明显。盐胁迫下接种R.i. 使刺槐叶片中MDA和H2O2的水平明显降低,叶片中SOD、CAT的活性增强;但刺槐根系中抗坏血酸盐过氧化物酶(APX)活性降低,H2O2水平升高,MDA水平与未接种AMF条件相比没有显著变化[65]。
3.2.3 缓解植物体内的离子胁迫 植物体内对K+和Na+吸收之间的平衡的变化(在芽和根中有更有利的K/Na平衡)可能是应对高盐度水平的主要机制。因此,盐胁迫下接种AMF的植物组织中K/Na比值增加,减轻高盐度水平所带来的影响[66-67]。例如,盐胁迫条件迫使酸枣(Hu)中Na+的含量上升,接种AMF后,植株的体内Na+的含量下降,有效地缓解了盐胁迫给植物带来的危害[68];对生菜(L. var. ramosa Hort.)接种两种AMF(F.m.和),与没有接种AMF的植物相比,接种AMF的植物体内K/Na比值上升,并且研究发现接种AMF的生菜可以通过增加PIP2的丰度以及上调LsaNHX基因表达,促使植物体内K/Na比值维持稳态,从而提高了植物的水分状况[69]。
4 总结与展望
AMF对植物水分吸收和利用的影响以及抗旱、盐胁迫主要体现在:①不同水分条件下,AMF通过改变宿主植物的根系结构,扩大根系的吸收总面积与活跃吸收面积,提高根对水分吸收的速率;在植物根部生成菌丝,菌丝吸收土壤水分并将水分运移至植物根系,提高植物对水分的吸收能力;②植物被AMF侵染后蒸腾作用明显提升,直接影响植物根系水分吸收纵向蒸腾拉力的提高;同时AMF通过抑制植物叶片的气孔阻力,改善植物叶片的气孔导度,从而提高植物的蒸腾拉力,提升植物的水分吸收能力;③干旱、盐胁迫下,植物通过接种AMF能够调节植物体内渗透调节物质的含量,改善植物体内抗氧化酶活性以及调节植物细胞内离子平衡,从而改善植物的水分状况,提高植物的耐旱性和耐盐性。
虽然菌根在植物水分的利用和获取方向有很大进展,但仍有很多问题需要进一步研究:①目前,大多数研究仍采用单一植物,在植物水分的吸收和利用上依旧是采用单一菌株或少量菌株的水平,不利于研究自然条件下菌根植物的水分状况;同时,对菌丝网络的研究中发现,不同植物能够通过菌丝网络形成一个整体[70],菌丝吸收水分后如何在不同植物间分配,现阶段也未有明确的表述;②干旱与盐胁迫下,不同种类的真菌与不同的植物形成共生体,植物激素与渗透调节物质显现的结果是不确定的,并且胁迫强度的不同也对菌根植物有很大的影响,探究AMF以何种方式影响植物体内抗氧化酶活性与渗透调节物质的含量还需进一步探究;③有研究发现,在不同程度的盐胁迫下,AMF对植物的侵染程度不同,并且盐胁迫程度越强,植物根系的AMF侵染率越低[71]。因此,在特定程度的盐胁迫下,如何提升AMF对植物的侵染率,是增强植物耐盐性的有效手段。另一方面,针对不同植物接种不同的AMF菌剂,使其在较低的菌根侵染率情况下,也能增强植物抵抗盐胁迫的能力,这特别对农作物有着显著的意义。
[1] 李涛, 杜娟, 郝志鹏, 等. 丛枝菌根提高宿主植物抗旱性分子机制研究进展[J]. 生态学报, 2012, 32(22): 7169–7176.
[2] Wang X X, Hoffland E, Feng G, et al. Phosphate uptake from phytate due to hyphae-mediated phytase activity by arbuscular mycorrhizal maize[J]. Frontiers in Plant Science, 2017, 8: 684.
[3] Wang X X, Wang X J, Sun Y, et al. Arbuscular mycorrhizal fungi negatively affect nitrogen acquisition and grain yield of maize in a N deficient soil[J]. Frontiers in Microbiology, 2018, 9: 418.
[4] Gao X P, Kuyper T W, Zou C Q, et al. Mycorrhizal responsiveness of aerobic rice genotypes is negatively correlated with their zinc uptake when nonmycorrhizal[J]. Plant and Soil, 2007, 290(1/2): 283–291.
[5] 袁丽环, 闫桂琴, 朱志敏. 丛枝菌根(AM)真菌对翅果油树幼苗根系的影响[J]. 西北植物学报, 2009, 29(3): 580–585.
[6] Marulanda A, Azcon R, Ruiz-Lozano J M. Contribution of six arbuscular mycorrhizal fungal isolates to water uptake byplants under drought stress[J]. Physiologia Plantarum, 2003, 119(4): 526–533.
[7] Wang W X, Zhang F, Chen Z L, et al. Responses of phytohormones and gas exchange to mycorrhizal colonization in trifoliate orange subjected to drought stress[J]. Archives of Agronomy and Soil Science, 2017, 63(1): 14–23.
[8] Li J Q, Meng B, Chai H, et al. Arbuscular mycorrhizal fungi alleviate drought stress in C3() and C4() grasses via altering antioxidant enzyme activities and photosynthesis[J]. Frontiers in Plant Science, 2019, 10: 499.
[9] Abdel-Salam E, Alatar A, El-Sheikh M A. Inoculation with arbuscular mycorrhizal fungi alleviates harmful effects of drought stress on damask rose[J]. Saudi Journal of Biological Sciences, 2018, 25(8): 1772–1780.
[10] Johnson N C, Graham J H, Smith F A. Functioning of mycorrhizal associations along the mutualism-parasitism continuum[J]. New Phytologist, 1997, 135(4): 575–585.
[11] Portes T A, Araújo B R. Comparison of the allocation of phytomass in soybean and bean and its potential role in biological nitrogen fixation[J]. Acta Scientiarum Agronomy, 2012, 34(3): 285–292.
[12] Wang B, Qiu Y L. Phylogenetic distribution and evolution of mycorrhizas in land plants[J]. Mycorrhiza, 2006, 16(5): 299–363.
[13] Primieri S, Magnoli S M, Koffel T, et al. Perennial, but not annual legumes synergistically benefit from infection with arbuscular mycorrhizal fungi and rhizobia: A meta-analysis[J]. The New Phytologist, 2022, 233(1): 505–514.
[14] Liu J, Guo C, Chen Z L, et al. Mycorrhizal inoculation modulates root morphology and root phytohormone responses in trifoliate orange under drought stress[J]. Emirates Journal of Food and Agriculture, 2016, 28(4): 251.
[15] Baldi E, Amadei P, Pelliconi F, et al. Use ofspp. and arbuscular mycorrhizal fungi to increase soil beneficial population of bacteria in a nectarine commercial orchard: Effect on root growth, nutrient acquisition and replanting disease[J]. Journal of Plant Nutrition, 2016, 39(8): 1147–1155.
[16] Whipps J M. Prospects and limitations for mycorrhizas in biocontrol of root pathogens[J]. Canadian Journal of Botany, 2004, 82(8): 1198–1227.
[17] 宋福强, 杨国亭, 孟繁荣, 等. 丛枝菌根(AM)真菌对大青杨苗木根系的影响[J]. 南京林业大学学报(自然科学版), 2005, 29(6): 35–39.
[18] 江盼盼, 宋述尧, 赵春波, 等. 三种丛枝菌根真菌对辣椒根系生长的影响及效应分析[J]. 中国蔬菜, 2010(6): 58–62.
[19] Liu C Y, Zhang F, Zhang D J, et al. Mycorrhiza stimulates root-hair growth and IAA synthesis and transport in trifoliate orange under drought stress[J]. Scientific Reports, 2018, 8(1): 1978.
[20] Hashem A, Kumar A, Al-Dbass A M, et al. Arbuscular mycorrhizal fungi and biochar improves drought tolerance in chickpea[J]. Saudi Journal of Biological Sciences, 2019, 26(3): 614–624.
[21] 余洁. 丛枝菌根真菌对荆条生长和抗旱性的影响[D]. 郑州: 河南农业大学, 2019.
[22] Basyal B, Emery S M. An arbuscular mycorrhizal fungus alters switchgrass growth, root architecture, and cell wall chemistry across a soil moisture gradient[J]. Mycorrhiza, 2021, 31(2): 251–258.
[23] 杨海霞, 李士美, 李敏, 等. 丛枝菌根真菌对白三叶耐盐性的影响[J]. 青岛农业大学学报(自然科学版), 2014, 31(2): 85–90.
[24] 苗启晨, 张晓楠, 张世杰, 等. 盐胁迫下丛枝菌根真菌(AMF)对尖瓣海莲幼苗生物量变化的影响[J]. 海南师范大学学报(自然科学版), 2017, 30(1): 14–18.
[25] 徐娜, 宋福强, 范晓旭, 等. 盐胁迫对菌根化沙枣幼苗生长性状的影响[J]. 东北林业大学学报, 2021, 49(6): 29–33.
[26] Anderson I C, Cairney J W G. Ectomycorrhizal fungi: Exploring the mycelial frontier[J]. FEMS Microbiology Reviews, 2007, 31(4): 388–406.
[27] Li X L, George E, Marschner H. Extension of the phosphorus depletion zone in VA-mycorrhizal white clover in a calcareous soil[J]. Plant and Soil, 1991, 136(1): 41–48.
[28] Khalvati M A, Hu Y, Mozafar A, et al. Quantification of water uptake by arbuscular mycorrhizal hyphae and its significance for leaf growth, water relations, and gas exchange of barley subjected to drought stress[J]. Plant Biology (Stuttgart, Germany), 2005, 7(6): 706–712.
[29] Ruth B, Khalvati M, Schmidhalter U. Quantification of mycorrhizal water uptake via high-resolution on-line water content sensors[J]. Plant and Soil, 2011, 342(1/2): 459–468.
[30] Bryla D, Duniway J M. Effects of mycorrhizal infection on drought tolerance and recovery in safflower and wheat[J]. Plant and Soil, 1997, 197(1): 95–103.
[31] Ruiz-Lozano J M. Arbuscular mycorrhizal symbiosis and alleviation of osmotic stress. New perspectives for molecular studies[J]. Mycorrhiza, 2003, 13(6): 309–317.
[32] Sands R, Theodorou C. Water uptake by mycorrhizal roots of radiata pine seedlings[J]. Functional Plant Biology, 1978, 5(3): 301.
[33] Smith S E, Jakobsen I, Grønlund M, et al. Roles of arbuscular mycorrhizas in plant phosphorus nutrition: Interactions between pathways of phosphorus uptake in arbuscular mycorrhizal roots have important implications for understanding and manipulating plant phosphorus acquisition[J]. Plant Physiology, 2011, 156(3): 1050–1057.
[34] 刘晓捷, 曾明, 杜建斌, 等. AMF对葡萄扦插苗矿质营养及生长的影响[J]. 西南农业大学学报(自然科学版), 2006, 28(2): 286–289, 318.
[35] Loewe A, Einig W, Shi L B, et al. Mycorrhiza formation and elevated CO2both increase the capacity for sucrose synthesis in source leaves of spruce and aspen[J]. The New Phytologist, 2000, 145(3): 565–574.
[36] Theodorou C. Soil moisture and the mycorrhizal association ofD. don[J]. Soil Biology and Biochemistry, 1978, 10(1): 33–37.
[37] Mexal J, Reid C P P. The growth of selected mycorrhizal fungi in response to induced water stress[J]. Canadian Journal of Botany, 1973, 51(9): 1579–1588.
[38] 李虹谕. 接种菌根真菌对水曲柳苗木根系吸收策略和叶片光合特性的影响[D]. 哈尔滨: 东北林业大学, 2019.
[39] 韩冰, 徐刚, 郭世荣, 等. 丛枝菌根真菌对苗期黄瓜生长及生理特性的影响[J]. 江苏农业学报, 2012, 28(6): 1392–1397.
[40] 董梅. 柴达木地区主要树种抗旱耐盐生理研究[D]. 北京: 北京林业大学, 2013.
[41] Augé R M, Toler H D, Saxton A M. Mycorrhizal stimulation of leaf gas exchange in relation to root colonization, shoot size, leaf phosphorus and nitrogen: A quantitative analysis of the literature using meta-regression[J]. Frontiers in Plant Science, 2016, 7: 1084.
[42] 宋会兴, 彭远英, 钟章成. 干旱生境中接种丛枝菌根真菌对三叶鬼针草(L.)光合特征的影响[J]. 生态学报, 2008, 28(8): 3744–3751.
[43] Sebastiana M, da Silva A B, Matos A R, et al. Ectomycorrhizal inoculation withtinctorius reduces stress induced by drought in cork oak[J]. Mycorrhiza, 2018, 28(3): 247–258.
[44] Dastogeer K M G, Zahan M I, Tahjib-Ul-Arif M, et al. Plant salinity tolerance conferred by arbuscular mycorrhizal fungi and associated mechanisms: A meta-analysis[J]. Frontiers in Plant Science, 2020, 11: 588550.
[45] 崔令军, 刘瑜霞, 林健, 等. 丛枝菌根真菌对盐胁迫下桢楠光合生理的影响[J]. 南京林业大学学报(自然科学版), 2021, 45(1): 101–106.
[46] 郑亚茹, 唐明. 丛枝菌根真菌对盐胁迫下桑树生长及光合特性的影响[J]. 蚕业科学, 2020, 46(6): 669–677.
[47] 田帅, 刘振坤, 唐明. 不同水分条件下丛枝菌根真菌对刺槐生长和光合特性的影响[J]. 西北林学院学报, 2013, 28(4): 111–115, 208.
[48] 岳海, 何双凌, 耿建建, 等. 丛枝菌根真菌对澳洲坚果幼苗耐旱性的影响[J]. 西部林业科学, 2020, 49(2): 30–35, 42.
[49] 孔静. 丛枝菌根真菌对几种草本植物抗旱性的影响研究[D]. 徐州: 中国矿业大学, 2015.
[50] 马坤, 杨建军, 李璐, 等. 接种丛枝菌根真菌后干旱胁迫对木棉根区土壤和体内养分的影响[J]. 中南林业科技大学学报, 2017, 37(11): 90–95, 102.
[51] 陈婕, 谢靖, 唐明. 水分胁迫下丛枝菌根真菌对紫穗槐生长和抗旱性的影响[J]. 北京林业大学学报, 2014, 36(6): 142–148.
[52] Wu Q S, He J D, Srivastava A K, et al. Mycorrhizas enhance drought tolerance ofby altering root fatty acid compositions and their saturation levels[J]. Tree Physiology, 2019, 39(7): 1149–1158.
[53] Huang Y M, Zou Y N, Wu Q S. Alleviation of drought stress by mycorrhizas is related to increased root H2O2efflux in trifoliate orange[J]. Scientific Reports, 2017, 7: 42335.
[54] Azmat R, Moin S. The remediation of drought stress under VAM inoculation through proline chemical transformation action[J]. Journal of Photochemistry and Photobiology B, Biology, 2019, 193: 155–161.
[55] Aalipour H, Nikbakht A, Etemadi N, et al. Biochemical response and interactions between arbuscular mycorrhizal fungi and plant growth promoting rhizobacteria during establishment and stimulating growth ofcypress (G.) under drought stress[J]. Scientia Horticulturae, 2020, 261: 108923.
[56] Langeroodi A R S, Osipitan O A, Radicetti E, et al. To what extent arbuscular mycorrhiza can protect chicory (L.) against drought stress[J]. Scientia Horticulturae, 2020, 263: 109109.
[57] Chandrasekaran M, Boughattas S, Hu S J, et al. A meta-analysis of arbuscular mycorrhizal effects on plants grown under salt stress[J]. Mycorrhiza, 2014, 24(8): 611–625.
[58] Evelin H, Kapoor R, Giri B. Arbuscular mycorrhizal fungi in alleviation of salt stress: A review[J]. Annals of Botany, 2009, 104(7): 1263–1280.
[59] 李倩, 郑爱琴, 马玉一, 等. AMF对紫花苜蓿抗盐生理特性的影响[J]. 草原与草坪, 2017, 37(5): 85–91.
[60] 金映虹, 严廷良, 叶绵源, 等. 丛枝菌根真菌对红树植物耐盐性的影响[J]. 湖北农业科学, 2016, 55(7): 1735–1738, 1743.
[61] Chang W, Sui X, Fan X X, et al. Arbuscular mycorrhizal symbiosis modulates antioxidant response and ion distribution in salt-stressedseedlings[J]. Frontiers in Microbiology, 2018, 9: 652.
[62] 高璿濛, 王玉丹, 杨春雪. 松嫩盐碱草地蒲公英根围AM真菌侵染特性及种类多样性[J]. 中国农业大学学报, 2019, 24(5): 90–97.
[63] 冯固, 李晓林, 张福锁, 李生秀. 施磷和接种AM真菌对玉米耐盐性的影响[J]. 植物资源与环境学报, 2000, 9(2): 22–26.
[64] Pan J, Peng F, Tedeschi A, et al. Do halophytes and glycophytes differ in their interactions with arbuscular mycorrhizal fungi under salt stress? A meta-analysis[J]. Botanical Studies, 2020, 61(1): 13.
[65] Chen J, Zhang H Q, Zhang X L, et al. Arbuscular mycorrhizal symbiosis mitigates oxidative injury in black locust under salt stress through modulating antioxidant defence of the plant[J]. Environmental and Experimental Botany, 2020, 175: 104034.
[66] Augé R M, Toler H D, Saxton A M. Arbuscular mycorrhizal symbiosis and osmotic adjustment in response to NaCl stress: A meta-analysis[J]. Frontiers in Plant Science, 2014, 5: 562.
[67] Klinsukon C, Lumyong S, Kuyper T W, et al. Colonization by arbuscular mycorrhizal fungi improves salinity tolerance of() seedlings[J]. Scientific Reports, 2021, 11(1): 4362.
[68] 申连英, 毛永民, 鹿金颖, 等. 丛枝菌根对酸枣实生苗耐盐性的影响[J]. 土壤学报, 2004, 41(3): 426–433.
[69] Santander C, Aroca R, Cartes P, et al. Aquaporins and cation transporters are differentially regulated by two arbuscular mycorrhizal fungi strains in lettuce cultivars growing under salinity conditions[J]. Plant Physiology and Biochemistry, 2021, 158: 396–409.
[70] Newman E I. Mycorrhizal links between plants: Their functioning and ecological significance[J]. Advances in Ecological Research, 1988, 18: 243–270.
[71] 张爱娣, 郑仰雄, 黄东兵. 丛枝菌根真菌对大叶女贞耐盐性的影响[J]. 江苏农业科学, 2018, 46(19): 129–133.
Research Progress of Arbuscular Mycorrhizal Fungi and Plant Symbiosis Affecting Plant Water Regime
PU Zitian1, ZHANG Lin2, ZHANG Chi3,4,5, WANG Hong3,4,5, WANG Xinxin3,4,5,6*
(1 College of Resources and Environment Science, Hebei Agricultural University, Baoding, Hebei 071001, China; 2 College of Resources and Environmental Sciences, China Agricultural University, Beijing 100193, China; 3 Agricultural Engineering Technology Research Center of National North Mountainous Area, Baoding, Hebei 071001, China; 4 Agricultural Technology Innovation Center in Mountainous Areas of Hebei Province, Baoding, Hebei 071001, China; 5 Hebei Mountain Research Institute, Hebei Agricultural University, Baoding, Hebei 071001, China; 6 Key Laboratory of North China Water-Saving Agriculture of Ministry of Agriculture and Rural Affairs, Baoding, Hebei 071001, China)
Arbuscular mycorrhizal fungi (AMF) can form symbionts with host plants and are widely found in terrestrial ecosystems. A large number of studies have shown that plants inoculated with AMF have stronger water absorption capacity and higher water use efficiency than plants not inoculated with AMF under different water conditions. Under drought and salt stress, AMF inoculation can effectively improve the drought and salt tolerances of host plants. This paper summarizes the research progresses of AMF, which is symbiotic with plants and can enhance the water absorption capacity of plant roots by enlarging root absorption area and improving root structure under different water conditions. The formation of extra-root hyphae networks in soil not only adds a new water absorption pathway (mycorrhizal pathway) to plants, but also regulates the water absorption by improving the mineral nutrition in plants, thus affecting the water absorption status of plants. Under different water conditions, the photosynthesis, transpiration and stomatal conductance of plants are all enhanced after AMF infection of roots. The enhancement of transpiration can directly and effectively improve the transpiration force of plants, thereby improving the absorption capacity of plants to water. At the same time, AMF inoculation can improve the water use efficiency, transpiration rate and net photosynthetic rate of plants, and improve the water use capacity of plants. Further, this paper reviews the serious influence of water stress (including drought and salt stresses) on plant water status, and inoculating AMF can improve the stress tolerance of plants by effectively adjusting the content of osmotic regulation substances in plants under water stress, the activity of antioxidant enzymes, the balance of ions in plants, and by enhancing the levels of plant photosynthesis and transpiration. This paper also discusses the research frontiers on the effects of AMF on water status and salt resistance of plants under different water conditions in order to provide theoretical reference for the improvement of plant water status through mycorrhizal interaction.
Arbuscular mycorrhizal fungi; Root system; Salinity stress; Drought stress; Water condition
S182
A
10.13758/j.cnki.tr.2022.05.002
蒲子天, 张林, 张弛, 等. 丛枝菌根真菌与植物共生影响植物水分状态的研究进展. 土壤, 2022, 54(5): 882–889.
华北作物改良与调控国家重点实验室自主课题(NCCIR2021ZZ-18)、国家重点研发计划项目(2021YFD1901001)和河北省重点研发计划项目(21327005D)资助。
(sywxx@hebau.edu.cn)
蒲子天(1996—),男,河北沧州人,硕士研究生,主要研究方向为农业资源与环境。E-mail: 2922321598@qq.com
猜你喜欢
陕西农业科学(2022年2期)2022-03-28
浙江农业学报(2021年5期)2021-05-31
园林科技(2020年2期)2020-01-18
中成药(2018年3期)2018-05-07
上海农业学报(2017年4期)2017-04-10
广东饲料(2016年5期)2016-12-01
环境与可持续发展(2013年6期)2013-03-11
植物营养与肥料学报(2011年5期)2011-11-06
植物营养与肥料学报(2011年6期)2011-10-24
植物营养与肥料学报(2010年3期)2010-11-16
