
褪黑素对低温胁迫下番茄多胺代谢及耐冷基因表达的影响
2022-12-02 05:29吴凤芝
福建农业学报 2022年7期
李 贤,杨 莲,吴凤芝
(东北农业大学园艺园林学院,黑龙江 哈尔滨 150030)
0 引言
【研究意义】番茄(Solanum lycopersicum)作为一种重要的蔬菜作物,喜温,对低温极其敏感,当环境温度低于12 ℃时易遭受冷害[1]。在中国北方地区,低温是越冬栽培的主要逆境,严重抑制番茄的生长发育[2]。因此,提高番茄的低温耐受性对番茄的生产具有重要意义。【前人研究进展】前人研究发现褪黑素和多胺在调节植物抗逆过程中有着相似的作用,另外胁迫环境下褪黑素还可以调控植物体内的多胺代谢。在干旱胁迫下褪黑素通过调节多胺代谢相关酶活性和基因表达水平,从而增强番茄耐热性[3]。Gong等[4]研究发现,在碱性环境中,经褪黑素处理的苹果植株中多胺合成酶ADC基因表达量提高,内源Put、Spd和Spm的积累量增多,电解质渗漏率降低,减轻了细胞膜损伤。张贵友等[5]发现褪黑素提高低温环境下烟草细胞ADC的活性,促进Put的合成来提高烟草悬浮细胞存活率。而Ke等[6]研究表明,盐胁迫下褪黑素通过抑制小麦叶片中多胺分解酶DAO和PAO活性,提高了Spd和Spm含量,降低了Put含量,有利于清除活性氧。这些结果表明,在不同植物中褪黑素通过调节多胺代谢,诱导不同种类多胺的积累,提高植物对胁迫的抗性[7]。【本研究切入点】低温胁迫下,褪黑素对番茄幼苗多胺代谢的影响如何?褪黑素主要通过影响哪种多胺来增强番茄耐冷性?该多胺主要通过哪条途径合成的?尚不完全清楚。除此之外,前人研究发现在拟南芥和西瓜等植物中褪黑素能诱导C2H2型锌指转录因子基因、AtCOR15以及热激蛋白基因(HSFs)等耐冷基因的表达,提高植物低温胁迫耐受性[8-10],在番茄响应低温胁迫时,ICE-CBF-COR信号转导通路基 因 (SlICE1、SlCBF1、SlCBF3和SlCOR413)、MAPK激酶(SlMAPK3、SlZAT12)表达水平的上调,对提高番茄植株耐寒性具有重要意义。然而褪黑素是否可以通过提高番茄ICE-CBF-COR信号转导通路基因(SlICE1、SlCBF1、SlCBF3和SlCOR413)、MAPK激酶(SlMAPK3、SlZAT12)的表达来响应低温胁迫增强番茄幼苗耐寒性尚不完全清楚。【拟解决的关键问题】以低温敏感型番茄品种东农708为试验材料,采用褪黑素预处理的方法,分析低温胁迫下番茄幼苗生长、多胺代谢相关指标以及耐冷基因表达的变化;并施用腐胺合成抑制剂,分析其对褪黑素诱导的番茄叶片中多胺含量、膜脂过氧化损伤以及抗氧化酶活性的影响。通过研究明确褪黑素如何对低温胁迫下番茄幼苗多胺代谢进行调控以及褪黑素对低温胁迫下番茄叶片耐冷基因表达的影响,以期进一步揭示褪黑素增强番茄幼苗耐冷性的机制,为褪黑素在生产上的应用提供理论支撑。
1 材料与方法
1.1 试验设计
试验于2019年5—12月进行。试验材料为东农708番茄品种(由东北农业大学番茄研究所提供),褪黑素(MT)、腐胺(Put)、Put合成抑制剂D-精氨酸(D-Arg)和二氟甲基鸟氨酸(DFMO),均购买自上海源叶生物技术公司。
1.1.1 褪黑素浓度筛选 将番茄种子于55 ℃下浸种消毒20 min后,置于恒温培养箱中催芽,种子露白后在智能人工气候植物培养箱(HPG-280HG)中播种于育苗盘内,培养条件:温度昼25 ℃/夜18 ℃(常温),光照强度20 000 lx,光周期昼14 h/夜10 h,空气湿度70%。幼苗长至2片真叶时分苗于塑料盆中(10 cm×10 cm)。待幼苗长至4叶1心时,分别用蒸馏水(CK)和浓度为 50 、100、200、300 μmol·L-1的外源MT于培养箱为黑夜条件时喷施于番茄叶片,以叶片滴水为度,每天喷1次,连续喷5 d后[11],将番茄幼苗转至另一培养箱进行低温胁迫处理(昼15 ℃/夜10 ℃),培养箱除温度设置改变外,其他参数保持不变。试验设6个处理:①蒸馏水处理后常温处理(CK);②蒸馏水处理后低温(C);③50 μmol·L-1褪黑素处理后低温处理(C+MT1);④100 μmol·L-1褪黑素喷施后置于昼15 ℃/夜10 ℃(C+MT2);⑤200 μmol·L-1褪黑素预喷施后置于昼 15 ℃/夜 10 ℃(C+MT3);⑥300 μmol·L-1褪黑素喷施后置于昼 15 ℃/夜10 ℃(C+MT4)。在低温处理的第0、2、4、8天采集番茄叶片,参考Li[12]的方法,根据番茄叶片MDA含量和电解质渗漏率筛选出缓解番茄植株膜损伤效果最佳的褪黑素浓度用于后续试验。
1.1.2 褪黑素对低温胁迫下番茄多胺代谢的影响番茄幼苗的培育和褪黑素(浓度100 μmol·L-1)的处理方式及时间同褪黑素浓度筛选试验,试验设计4个处理:①常温对照(CK);②常温下褪黑素处理(MT);③低温处理(C);④褪黑素预处理后进行低温处理(C+MT)。在低温处理的第0、2、4、6、8天进行番茄叶片取样,用于测定多胺代谢相关指标(多胺和前体物质含量、合成和分解酶活性及基因表达量);在低温处理的第8天,测定番茄幼苗的生长指标。
为了探究低温胁迫下褪黑素通过哪条途径诱导Put的合成,以及Put是否在褪黑素提高番茄幼苗耐冷性中起重要作用,选用Put合成抑制剂:0.1 mmol·L-1的D-精氨酸(D-Arg,抑制精氨酸途径)和二氟甲基鸟氨酸(DFMO,抑制鸟氨酸途径)[13]。试验共设5个处理:①常温处理(CK);②低温处理(C);③褪黑素预处理后低温处理(C+MT);④早上6:00用D-Arg预处理12 h后用褪黑素处理,联合处理5 d后低温处理(C+D-Arg+MT);⑤早上 6:00用 DFMO处理,12 h后用褪黑素处理,联合处理5 d后低温处理(C+DFMO+MT)。未经抑制剂处理的组(①-③)在同一时间喷施蒸馏水或褪黑素。在低温处理的第4天进行番茄叶片取样,测定多胺含量、MDA含量、电解质渗透率和抗氧化酶活性。
1.1.3 褪黑素对低温胁迫下番茄耐冷基因表达的影响试验设计及褪黑素处理方式同1.1.2,褪黑素预处理后进行5 ℃低温处理,在低温处理的第0、1、3、6、12和24 h选取番茄幼嫩叶片取样,分析SlMAPK3、SlICE1、SlCBF1、SlCBF3、SlCOR413和SlZAT12基因的表达情况。
1.2 测定项目与方法
1.2.1 丙二醛含量和电解质渗漏率的测定 采用硫代巴比妥酸法测定MDA含量[14];电解质渗透率的测定参考Ishitani等[15]的方法。
1.2.2 生长指标的测定 用直尺测量番茄植株茎基部到生长点的长度作为株高,然后用清水将植株洗干净,擦干水分后将植株地上部和地下部分开,用电子天平称量鲜质量;然后在80 ℃下烘干至恒重后称量干质量。
1.2.3 抗氧化酶活性的测定 配制磷酸缓冲溶液,作为番茄中的抗氧化酶的提取液,缓冲溶液由50 mmol·L-1磷酸盐缓冲溶液(pH 7.8)、50 mmol·L-1EDTA二钠盐和2%(m/V)聚乙烯吡咯烷酮(PVP)组成。称取0.3 g番茄叶片于研钵中,加入5 mL预冷的磷酸缓冲溶液,在冰浴中快速研磨成匀浆,然后转移至10 mL离心管中,在4 ℃下离心(15 000 g,25 min)。取上清液用于测定抗氧化酶活性,SOD酶活性参考Giannopotitis[16]的方法进行测定;POD活性根据Nickel[17]的方法进行测定;CAT活性按照Dhindsa等[18]的方法测定;APX活性的测定参考Pinhero[19]的方法,略有改动。
1.2.4 多胺含量的测定 多胺含量的测定方法参考Duan等[20]的方法,略作修改。番茄叶片中的多胺用5%高氯酸(HClO4)提取,经苯甲酰氯衍生化后用乙醚萃取,多胺的分离采用高效液相色谱法,色谱柱采用 C18液相色谱柱(Hypersil BDS C18,4.0×150 mm,5 μm ),检测器为 Waters 2998紫外检测器,检测波长为254 nm,流动相为V(甲醇)∶V(水)=3∶2,流速保持 1 mL· min-1。
1.2.5 多胺合成酶活性测定 参考Zhao等[21]的方法测定番茄叶片中多胺合成酶活性,略作修改。番茄叶片在预冷的磷酸盐缓冲溶液(pH 8.0)中研磨,离心所得的上清液为酶提取液,取酶提取液,与L-精氨酸(25 mmol·L-1,测定 ADC 酶活性)、L-鸟氨酸(25 mmol·L-1,测定ODC酶活性)或S-腺苷甲硫氨酸(25 mmol·L-1,测定SAMDC活性)进行反应,离心后上清液用苯甲酰氯衍生,衍生产物用乙醚萃取,收集乙醚相吹干并溶于甲醇中,然后检测254 nm处的OD值。
1.2.6 多胺分解酶活性测定 参考Su[22]的方法测定番茄叶片中多胺分解酶活性。配制磷酸缓冲液(pH 6.5),用于提取番茄叶片中的酶,取酶提取液,加入辣根过氧化物酶溶液(250 U·mL-1)和显色液(10 mg 4-氨基氨替吡啉和25 μL N,N-二甲基苯胺),于30 ℃恒温水浴锅中反应 5 min,加入 Put(20 mmol·L-1,测定 DAO活性)或 Spd+Spm(20 mmol·L-1,测定PAO活性),测定555 nm处连续的OD值,以每分钟OD值变化0.001为1个酶活力单位。
1.2.7 多胺合成前体物质含量测定 采用异硫氰酸苯酯(PITC)衍生法[23]分析番茄叶片中多胺前体物质精氨酸、鸟氨酸和甲硫氨酸的含量。取番茄叶片,在液氮中快速研磨成粉末,然后加入HCl进行提取,4 ℃下离心30 min后取上清液进行衍生,衍生液由三乙胺、乙腈和异硫氰酸苯酯溶液组成,混合液在室温下静置1 h,然后加入正己烷进行萃取,收集下层清液用于高效液相色谱-紫外检测分析,波长为254 nm,色谱柱采用ACE Excel SuperC18 5 μm 4.6×250 mm色谱柱,进样量20 μL,流动相A为50 mmol·L-1V(乙酸钠)(用乙酸调节pH至6.5)∶V(乙腈)=93∶7,流动相 B为V(乙腈)∶V(水)=4∶1,梯度洗脱,洗脱程序如表1所示。

表1 多胺合成前体物质检测流动相梯度洗脱程序Table 1 Mobile phase elution for detection of polyamine synthesis precursors
1.2.8 基因表达量的测定 用Trizol试剂提取番茄叶片总RNA,然后用反转录试剂盒合成cDNA,基因特异引物(表2)由生工生物工程(上海)股份有限公司合成。在Analytic jena荧光定量PCR仪上进行扩增反应,采用2-ΔΔt法计算基因的相对表达量。

表2 qRT-PCR引物序列Table 2 Primer sequences used in qRT-PCR
1.3 数据分析
使用SPSS软件分析数据,方差分析采用Turkey’s HSD,在P<0.05水平下进行分析,使用Origin 2017绘图。
2 结果与分析
2.1 褪黑素对低温胁迫下番茄膜脂过氧化损伤和生长的影响
如图1所示,与对照相比,低温胁迫下番茄幼苗叶片中MDA含量和电解质渗漏率显著提高(P<0.05),而MTx预处理不同程度地降低了MDA含量和电解质渗漏率,当褪黑素浓度为100 μmol·L-1时上述两个指标降低幅度最大,且显著低于单独低温处理(P<0.05),因此选用 100 μmol·L-1的 MT 进行后续试验。如表3 所示,常温下MT处理对番茄幼苗的生长无影响,低温胁迫下番茄幼苗的株高、地上部鲜质量和干质量、地下部鲜质量和干质量以及全株鲜重和干重显著低于对照(P<0.05);与低温胁迫相比,低温胁迫下MT预处理显著提高了番茄植株的上述生长指标(P<0.05)。
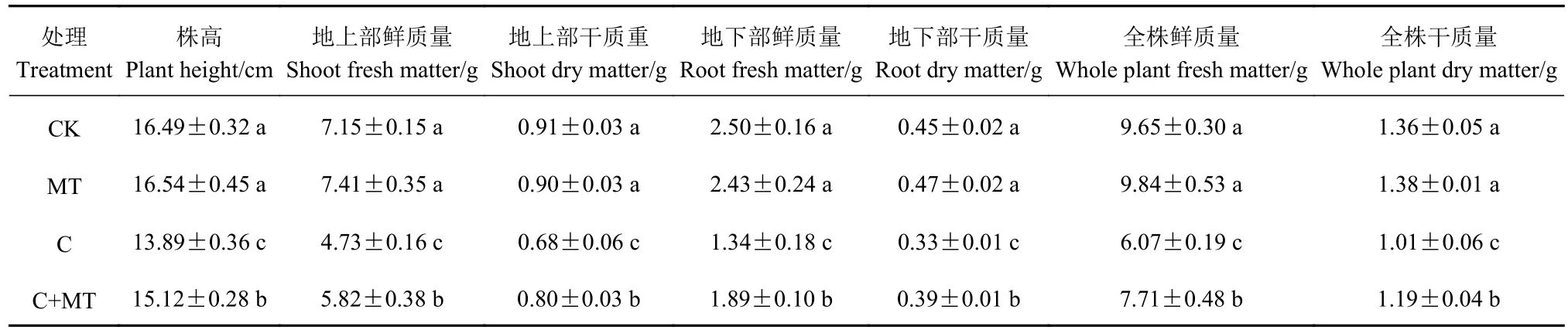
表3 褪黑素对低温胁迫下番茄幼苗生长的影响Table 3 Effects of melatonin spray on growth of tomato seedlings under low temperature stress
2.2 褪黑素对低温胁迫下番茄叶片多胺代谢的影响
2.2.1 褪黑素对低温胁迫下番茄叶片多胺及前体物质含量的影响 如图2所示,常温下MT处理对3种多胺的含量无影响;与对照相比,低温胁迫初期(第2天)番茄叶片中3种多胺以及前体物质Arg与Met的含量显著增加,第4~6天Spd、Spm和Met含量显著降低(P<0.05);低温胁迫下MT预处理的番茄叶片中Put与Arg含量在所有时期显著高于低温处理,Spd、Spm和Met含量仅在第2天差异显著(P<0.05),表明低温胁迫下MT预处理主要提高了低温胁迫下番茄叶片中Put和Arg的含量。
2.2.2 褪黑素对低温胁迫下番茄叶片多胺合成酶活性的影响 如图3 所示,与对照相比,低温胁迫下番茄叶片中的ADC活性在整个低温处理期间显著提高,ODC和SAMDC活性在第2天显著提高,SAMDC在第4~6天显著降低(P<0.05);低温胁迫下MT预处理进一步提高了番茄叶片中ADC活性,且在各时期差异均显著,而ODC活性无明显变化,SAMDC活性仅在第2天差异达到显著水平(P<0.05)。从低温胁迫下经MT预处理的番茄叶片中多胺合成酶和前体物质含量的变化情况可初步推测在Put的两条合成途径中,精氨酸途径可能是MT诱导的番茄叶片中Put合成的主要途径。
2.2.3 褪黑素对低温胁迫下番茄叶片多胺合成酶基因表达的影响 如图4 所示,与对照相比,低温胁迫显著提高了所有时期番茄叶片中SlADC1与SlADC2的相对表达量,SlSAMDC1基因表达量在第2天显著提高,第4~6天显著降低,SlSAMDC2基因表达量在第4~8天显著降低(P<0.05);MT预处理进一步提高了SlADC1与SlADC2的相对表达量,且在低温处理的各时期差异均显著,SlSAMDC1的相对表达量在第2天差异达到显著水平,其余时期差异不显著(P>0.05)。
2.2.4 褪黑素对低温胁迫下番茄叶片多胺分解酶活性及其基因表达的影响 如图5 所示,对照和常温下MT处理的番茄叶片中DAO和PAO活性较稳定;低温胁迫显著提高了番茄叶片DAO和PAO活性及基因相对表达量(SlDAO和SlPAO),且随着低温处理时间的延长呈升高趋势(除了第8天的PAO)(P<0.05);MT预处理显著抑制了低温胁迫下DAO活性和SlDAO相对表达量的升高,且在低温处理的各时期差异均达到显著水平,而对PAO活性及SlPAO相对表达量无显著影响(P>0.05)。
2.3 腐胺合成抑制剂对褪黑素诱导的番茄叶片多胺含量和抗氧化特性的影响
为了进一步探究低温胁迫下MT诱导的番茄叶片中Put的合成主要通过哪条途径,以及探究Put是否介导了MT对番茄幼苗抗氧化特性的增强作用,选用两种Put合成抑制D-Arg(抑制精氨酸途径)和DFMO(抑制鸟氨酸途径)进行研究。
2.3.1 腐胺合成抑制剂对褪黑素诱导的番茄叶片多胺含量的影响 如图6 所示,低温胁迫下MT预处理能显著提高番茄叶片中Put的含量,采用精氨酸脱羧酶抑制剂D-Arg与褪黑素联合预处理,番茄叶片中Put含量显著低于MT预处理的植株,Spd和Spm含量也显著降低(P<0.05);而采用鸟氨酸脱羧酶抑制剂DFMO与褪黑素联合预处理,MT诱导的Put含量不变。说明低温胁迫下MT诱导的番茄叶片中Put的合成主要通过精氨酸途径。
2.3.2 腐胺合成抑制剂对褪黑素诱导的番茄叶片膜脂过氧化损伤和抗氧化酶活性的影响 如图7 所示,低温胁迫下MT预处理的番茄叶片中MDA含量和电解质渗漏率显著低于低温处理,SOD、POD、CAT和APX活性显著高于低温处理,然而与低温胁迫下MT预处理相比,D-Arg与MT联合预处理MDA含量、电解质渗漏率显著提高,抗氧化酶活性显著降低(P<0.05),恢复至单独低温处理的水平,而DFMO与MT联合预处理上述指标没有显著变化。
上述2.2和2.3的结果表明低温胁迫下MT通过正向调节Arg途径促进番茄叶片中Put合成,以及下调Put分解酶DAO基因表达,抑制DAO活性,从而促使Put积累,清除细胞过量ROS,减轻番茄植株所受的膜脂过氧化损伤。
2.4 褪黑素对低温胁迫下番茄叶片耐冷基因表达的影响
如图8 所示,低温胁迫诱导了番茄叶片中耐冷基因的表达,其中SlICE1和SlCBF3在低温处理1 h时达到最大值,SlCBF1和SlMAPK3在3 h时达到最大值,SlCOR413和SlZAT12在6 h时达到最大值;低温胁迫下褪黑素预处理在不同程度上进一步提高了番茄叶片中上述基因的表达量,与低温处理相比SlICE1和SlMAPK3基因相对表达量在1~6 h显著上调(P<0.05);SlCBF1在12 h内差异均达到显著水平,比仅低温处理提高了36.10%~82.14%(P<0.05);SlCBF3除了12 h时,差异均显著(P<0.05);SlCOR413在3~24 h差异显著,6 h时增幅最大,达到49.23%;SlZAT12的表达量在低温处理的所有时期均显著提高,3 h时增幅最大,比低温处理提高了172.63%(P<0.05)。
3 讨论
3.1 褪黑素影响低温胁迫下番茄幼苗膜脂过氧化
细胞膜系统是植物遭受冷害的主要部位[24],低温胁迫下植物CO2同化过程受到抑制,植物对光能的吸收和利用失衡,引起ROS过量积累,导致膜脂过氧化,MDA积累,膜透性变大,导致电解质外渗,引起植物细胞膜氧化损伤,抑制植物生长[25]。因此,MDA和电解质渗漏率通常被认为是判断膜结构是否完整的指标,用来衡量植物遭受胁迫伤害的程度[26]。本研究中,番茄在低温胁迫期间,其叶片MDA含量和电解质渗漏率持续增加,株高和鲜/干质量显著降低,表明低温对番茄造成了氧化损伤,且这一伤害随低温胁迫时间的延续而加剧,这与刁倩楠等[27]的研究结果一致。同时本研究还发现相较于单独低温处理,100 μmol·L-1褪黑素预处理后再进行低温处理能显著降低番茄叶片中MDA含量和电解质渗漏率,增加番茄幼苗株高和干鲜质量,这表明褪黑素能减轻低温胁迫对番茄植株造成的膜脂过氧化损伤,保护细胞膜系统,同时缓解低温胁迫对番茄幼苗生长的抑制作用,这与盐胁迫下褪黑素对黄瓜幼苗的影响研究结果一致[28]。但褪黑素具体是如何缓解低温胁迫导致的膜脂过氧化需做进一步研究探讨。
3.2 褪黑素影响低温胁迫下番茄的多胺代谢
当胁迫存在时,外源褪黑素可以提高植物体内多胺的水平[29]。例如高温胁迫下褪黑素可通过提高多胺合成酶基因的表达促进番茄叶片中多胺的合成,强化抗氧化系统来增强番茄幼苗耐热性[30]。Zhang等[31]发现在涝害环境下外源施用褪黑素,苜蓿叶片中多胺合成酶活性增加,分解酶活性降低,多胺含量大幅度增加。在低温胁迫下,褪黑素通过提高Put和Spd水平,稳定Spm水平,减轻了黄瓜幼苗低温胁迫[32],这与我们的研究结果有所差异,本研究中,低温处理的各个时段,褪黑素预处理后低温处理的番茄叶片中Put含量均显著高于单独低温处理,但Spd和Spm的含量虽有所提高,但除第2天外均未达到显著水平,这可能是由于Put、Spd和Spm这3种多胺在不同的物种中及不同的胁迫下的响应存在差异。本研究结果表明,在低温胁迫下褪黑素对番茄叶片中Put的积累有积极作用。有研究表明腐胺(Put)作为植物抵御冷害的一种保护性物质,其含量越高,植物抗寒能力越强[33],Song[34]的研究也发现低温胁迫下番茄中植株中Put含量增加,植株耐寒性增强,这是因为腐胺带正电荷,能与细胞磷脂和核酸等结合,加强对细胞膜等组织的保护作用;其次腐胺作为一种可溶性物质,在胁迫环境下能够帮助平衡细胞的渗透压,保持细胞膜的稳定性,防止细胞内溶质外渗,以提高植物耐冷性;再者腐胺能调节活性氧代谢平衡减轻植物胁迫损伤[35,36]。上述研究均表明低温胁迫下褪黑素很可能是通过番茄中腐胺含量,从而缓解低温胁迫导致的膜脂过氧化。
对褪黑素具体如何影响低温胁迫下腐胺在植株体内的积累,我们做了进一步的研究探讨,在植物体内Put的合成途径主要有精氨酸(Arg)途径和鸟氨酸(Orn)途径(对应2个合成前体Arg和Orn),分别由精氨酸脱羧酶(ADC)和鸟氨酸脱羧酶(ODC)催化[13]。本试验结果表明低温处理前用褪黑素预处理的番茄叶片中Arg含量、ADC活性以及ADC合成相关基因的表达水平显著高于低温处理,而Orn含量、ODC活性与ODC合成相关基因的表达水平无显著变化,这表明低温胁迫下褪黑素是通过诱导ADC催化精氨酸途径介导腐胺的合成缓解低温胁迫。为了验证这一结论,我们用精氨酸途径抑制剂D-精氨酸(D-Arg)和鸟氨酸途径抑制剂二氟甲基鸟氨酸(DFMO)分别与褪黑素进行了联合预处理,结果发现低温胁迫下,D-Arg显著降低了褪黑素诱导的番茄叶片中Put的含量,而DFMO对褪黑素诱导的Put的含量无影响,这进一步表明褪黑素诱导低温胁迫下番茄叶片中Put的合成主要是通过精氨酸途径,同时,相较于MT预处理,D-Arg与MT联合预处理的番茄在低温胁迫下,其叶片MDA含量、电解质渗漏率显著提高,抗氧化酶活性显著降低,恢复至单独低温处理的水平,而DFMO与MT联合预处理后上述指标没有显著变化。这说明褪黑素可能是通过影响介导Put合成的精氨酸途径来减轻低温胁迫下番茄叶片细胞膜脂过氧化损伤程度以及增强抗氧化酶活性。
3.3 褪黑素影响低温胁迫下番茄幼苗耐冷基因的表达
植物已进化出多种策略应对包括冷胁迫在内的多种环境胁迫。前人研究发现植物的细胞膜首先上感知胁迫信号,随后转导至第二信使,并激活下游胁迫响应基因,最终诱导整个植物水平的保护性生理响应[37]。目前在植物低温响应中,以CBF转录因子为核心的ICE1-CBF-COR信号感知-转导途径被研究得最为清楚。
研究发现植物在感知到低温信号后,其转录因子CBFs基因会被迅速诱导表达,随后CBFs蛋白通过与下游冷响应基因COR的特定顺式元件进行结合来激活冷响应基因,从而增强植物的抗寒性[38,39]。番茄有3个CBFs基因,SlCBF1基因表达与植株耐寒性呈正相关[40],因此,SlCBF1表达水平通常被用来量化番茄植株的抗寒性。此外,ICE1作为CBF的诱导因子,诱导CBF1表达,有研究就表明,低温胁迫下番茄SlICE1的高表达提高了SlCBF1的表达量,增强了番茄的抗寒性[25]。本研究发现,番茄叶片中SlICE1、SlCBF1和SlCBF3在低温处理中的相对表达量显著高于空白对照,同时在褪黑素与低温联合处理中的转录水平显著高于仅低温处理,这与Shi等[8]在拟南芥上的研究结果一致。COR蛋白是植物冷害调节通路ICE1-CBF/DREB1-COR的终末蛋白[32]。有研究发现在番茄植株中SlCOR413的过表达减轻了低温引起的叶绿体膜和结构的损伤,而抑制SlCOR413的表达则加重了叶绿体损伤[41]。低温胁迫会诱导SlCOR413表达,有助于增强番茄对低温胁迫的抗性[42],与本研究的结果一致。另外本研究中还发现褪黑素显著增加了低温胁迫下番茄叶片中SlCOR413的相对表达量,这表明褪黑素可以保护低温胁迫下番茄的叶绿体膜,以维持叶绿体相对正常的功能和较高的光合能力来对抗冷害。综上,褪黑素能够通过提高低温胁迫下番茄叶片ICE1-CBF-COR通路基因(SlICE1、SlCBF1、SlCBF3、SlCOR413)的转录水平,正向调节番茄对低温胁迫的响应。而Kou等[43]的研究发现腐胺处理植株及SaADC1转基因株系中CBFs的表达明显被增强,在转基因株系中CBF下游的基因CORs的表达同样得到增强,说明CBF-COR途径可能会受ADC1介导的腐胺合成途径影响,促进低温相关代谢物质的积累,从而调节马铃薯的低温抗性,因此我们推测,褪黑素可能是通过提高番茄叶片中腐胺含量促进CBFs和CORs基因上调表达进而增强植株耐寒性。
在ICE1-CBF-COR信号通路中,ICE1作为上游关键转录因子直接调控CBFs基因,与植物低温胁迫抗性密切相关[44]。有研究表明植物的CBFs基因会在低温诱导的前期得到一定程度的积累,增强植物对低温的耐受性,但CBFs基因的过高表达对植物反而是一种伤害[45,46],这时植物便会通过MAPK3/6磷酸化ICE1蛋白,抑制CBFs基因的过表达,使植物能够顺利抵御低温胁迫的同时还能良好生长[47,48]。Ramirez等[49]研究也表明MPK3/6-ICE1-CBF调控途径创造了低温胁迫和生长之间的平衡,从而提高植物暴露到低温胁迫下的存活率。Wang等[50]研究表明低温胁迫能激活SlMAPK3增强番茄的耐冷性,这与本研究结果一致。在本研究中我们还发现,低温胁迫下褪黑素处理显著提高了SlMAPK3基因表达量,有研究表明Spm能够活化MAPK,诱导MAPK相关基因的表达[51]。而Spm的合成离不开腐胺,同时我们的研究已发现褪黑素可以促进低温胁迫下番茄叶片中腐胺的合成,因此我们猜测褪黑素可能是通过诱导腐胺的合成影响SlMAPK3基因,促使ICE1蛋白磷酸化减轻因低温胁迫诱导的CBFs基因的过表达而对生长发育造成的抑制作用,使得植株在抵御低温胁迫得同时还能正常生长发育。
ZAT12是锌指蛋白转录因子,在激活活性氧信号通路以及抗氧化系统方面发挥关键作用[52]。Bajwa等[9]发现,C2H2型锌指蛋白和CBFs均参与拟南芥中褪黑激素介导的低温应激反应。本试验研究结果表明低温胁迫下褪黑素预处理显著提高了低温胁迫下SlZAT12基因的表达量,这与Li等[53]在平邑甜茶上的研究结果一致。并且褪黑素预处理提高了低温胁迫下番茄叶片抗氧化酶的活性,因此褪黑素可能通过激活番茄植株中ROS信号通路关键转录因子基因SlZAT12的表达,增强抗氧化系统,缓解番茄植株氧化损伤。除此之外,Zhao等[54]研究发现在低温条件下ZAT12通过上调ADC1和ADC2表达触发精氨酸途径,进而促进Put合成,这表明褪黑素可能通过提高SlZAT12的表达水平促进腐胺的合成,从而增强番茄植株的抗寒性。
4 结论
100 μmol·L-1的褪黑素可通过正向调节精氨酸途径促进Put合成,以及抑制Put分解来增加Put的积累量,这减轻了番茄的膜脂过氧化损伤,提高了番茄抗氧化能力,缓解低温胁迫对番茄幼苗生长的抑制作用。除此之外,褪黑素促进了番茄叶片中与腐胺密切相关的低温响应基因表达,增强了番茄幼苗低温胁迫耐受性。褪黑素可能通过影响腐胺的合成,调控耐冷基因表达,从而提高番茄植株低温胁迫耐受性,更深入的机理有待进一步验证。
猜你喜欢
材料与冶金学报(2022年2期)2022-08-10
温州大学学报(自然科学版)(2022年2期)2022-05-30
今日农业(2021年10期)2021-11-27
今日农业(2021年9期)2021-11-26
军事文摘(2021年16期)2021-11-05
诗潮(2021年1期)2021-03-15
建材发展导向(2021年23期)2021-03-08
今日农业(2020年23期)2020-12-15
劳动保护(2018年8期)2018-09-12
山东工业技术(2016年15期)2016-12-01
