
林隙大小和隙内位置对小兴安岭蒙古栎林内红松光合能力的影响
2022-11-30 01:02李佳欣牟长城田博宇
南京林业大学学报(自然科学版) 2022年2期
李佳欣,牟长城*,田博宇,叶 林
(1.东北林业大学林学院,生态研究中心,黑龙江 哈尔滨 150040;2.伊春市林业科学研究院,黑龙江 伊春 153000)
林隙在森林恢复和管理中发挥着重要作用[1],通过影响直接到达地面的太阳辐射量,进而影响森林植物的生存与生长、更新和群落演替及森林组成与结构的维持[2-4]。因此,林隙管理是建立可持续森林经营管理系统的关键[5]。抚育经营是森林中常见的人工干预方式,对森林采取不同强度的抚育措施会形成大小不同的林隙,而幼苗在不同大小林隙中的生长发育状况能够反映其对林隙内微环境的适应能力和更新潜力,可为优化抚育经营措施提供理论参考[6]。
林隙大小和林隙内位置是林隙的两个关键特征,通过影响林隙内微环境(光照和温度)而对树种更新和生长起重要作用[7-9]。近年来对林隙内树木光合的研究已成为热点,但对热带地区树种研究较多[10-13],对温带地区树种研究较少[14-15];对伴生树种研究较多[16-17],对优势种或濒危种研究较少[18-19]。此外,林隙大小对树种更新组成有较大影响,不耐阴种或早期演替种通常只占据较大林隙[4,20-21],其更新所需最小林隙为400 m2或1 000 m2[22-23];而耐阴种或后期演替种在较小或较老树种林隙中生长较好。一般而言,为获得良好的天然更新,总体上应维持林隙直径与林隙形成木树高比值在1.0~1.5[24]。近年来相关研究主要集中在揭示林隙内更新幼苗光合能力的内在形态和生理生态学响应机制方面,并对一些重要树种更新与生长所需的最适林隙微生境进行筛选[10-19]。目前,学者们对红松(Pinuskoraiensis)光合的研究主要涉及干旱胁迫[25]、边缘效应[26]、不同光照条件[27-29]和叶龄[30-32]等方面,而有关林隙对红松光合作用的影响鲜见报道,仅见温带阔叶红松林中红松(15 a)的最大光合速率随林隙增大呈先增大后减小[33]。此外,以往研究大多是在温室或人工气候箱中进行,且多以幼苗为试验材料,而在自然状态下对幼树的研究则很少[30,32]。
阔叶红松林作为我国东北温带地带性顶极植被,在维护东北区域生态安全及供应优质林木方面有重要作用[34-36],其群落建群种红松不仅是国家保护野生树种,也是东北林区珍贵的用材树种[37]。由于长期不合理的采伐天然红松林已消失殆尽,绝大部分已退化为次生林,而次生林冠下栽植红松是恢复阔叶红松林的有效途径。但对于退化后形成的次生林林隙大小和林隙内位置对优势种红松的光合和生长的影响仍不清楚。为此,本研究以小兴安岭蒙古栎(Quercusmongolica)次生林内红松幼树(15 a)为对象,测定不同大小林隙及林隙内位置的红松光合参数及环境因子,揭示次生林林隙大小和林隙内位置对红松幼树光合能力的影响规律及机制,为红松在天然次生林内的更新及可持续经营提供科学依据。
1 材料与方法
1.1 研究区概况
研究地点位于小兴安岭伊春市金山屯林业局金山屯区育林经营所(129°43′E,47°42′N),属于大陆性湿润季风气候,年均气温1.5 ℃,年均降水量625 mm,多集中在7—9月,无霜期约115 d。地带性土壤为暗棕壤,地带性植被为以红松为群落建群种的温带针阔混交林,但目前多已退化为各类型的次生林(蒙古栎林、杨桦林、硬阔叶混交林和和杂木林等)和人工林。育林经营所过去开展过蒙古栎次生林冠下营造红松恢复红松林的试验,试验地面积为50 hm2,林冠下栽植红松幼苗密度为3 300株/hm2,并在2003年进行了不同强度的上层透光采伐抚育(采伐蓄积占比分别为1/5、2/5和小块皆伐)。试验地乔木层包括蒙古栎(主要组成树种,样地占比67%)、色木枫(Acerpictum)、山杨(Populusdavidiana) 和紫椴(Tiliaamurensis)等;灌木层主要有毛榛(Corylusmandshurica) 和刺五加(Eleutherococcussenticosus)等;草本层主要有大叶章(Deyeuxiapurpurea)。2018年进行本次调查时,透光抚育试验已长达15 a,已初步形成了异龄复层针阔混交林,由于林下红松幼树长期处于不同大小的林隙及林隙内不同位置中,红松生长存在较大差异,且蒙古栎为干旱树种,生长缓慢,透光抚育15 a后上层林木蓄积量并未发生较大改变,故上层林木生长对本试验所选择的各大小林隙样地的影响并不大,这为本研究提供了理想场所。
1.2 研究方法
1.2.1 样地设置
以蒙古栎次生林(林隙形成木蒙古栎在林内平均林分密度为1 774株/hm2,平均胸径10.5 cm,平均树高12.2 m)内的红松幼树(15 a)为研究对象,依据耐阴树种生长所需的最佳林隙直径与林隙形成木树高的比值(D/H)<1.0[38],选取4种不同大小的林隙:大林隙(小块皆伐,206.1 m2)、中林隙(采伐蓄积占比40%,116.9 m2)、小林隙(采伐蓄积占比20%,52.4 m2)和对照(林内未采伐样地,12.6 m2);每个处理3个重复,共计12个林隙样地,且各样地均处于西南坡中部(立地条件一致),各林隙样地具体情况见表1。
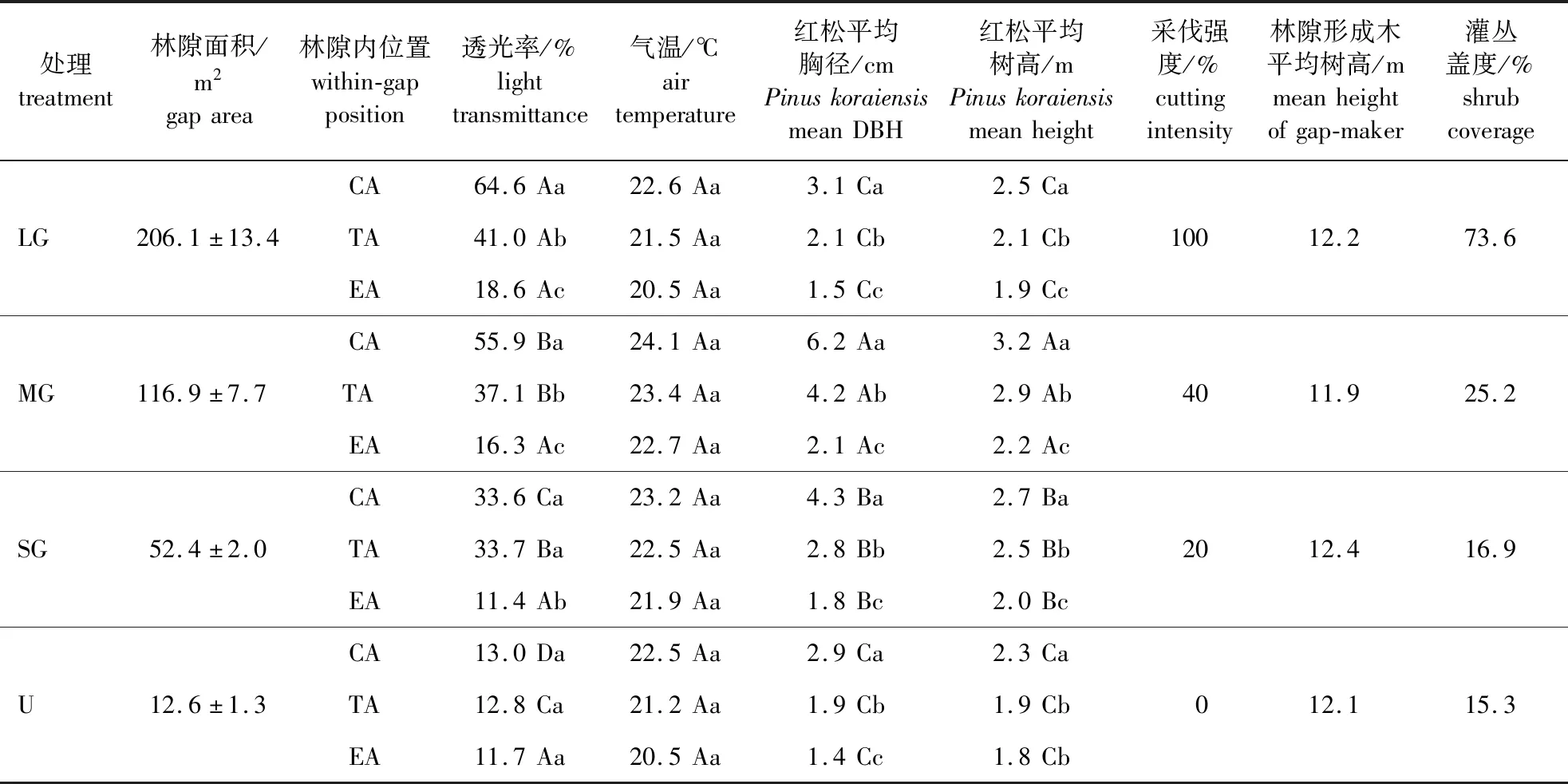
表1 小兴安岭蒙古栎林各大小林隙基本特征Table 1 Basic characteristics of various gaps of the Quercus mongolica forests in Xiaoxing’an Mountains
再将林隙按照中心区、过渡区和边缘区分成3个同心圆环(图1),在每个区内各选择1株红松幼树样木(每个处理12株),共计36株,测定其光合参数、叶绿素含量及环境因子。

图1 小兴安岭蒙古栎林各大小林隙内位置划分示意Fig.1 A schematic diagram of the division of positions in various gaps of the Quercus mongolica forests in Xiaoxing’an Mountains
1.2.2 光合参数的测定
于2018年生长季(5—10月)开展试验,采用CIRAS-2光合仪(PP Systhems,USA),选择晴朗天气的7:30—11:00对各林隙内红松光合参数进行测定,每月1次。即选取红松幼树(15 a)树冠中部同一高度的向阳方位和背阴方位叶片,测量前设定最高光照强度对针叶进行充分光诱导,利用缓冲瓶使叶室CO2浓度与大气CO2浓度保持一致,待仪器稳定后开始记录。测定时将叶簇平铺于叶室内,将LED光源设置自低到高的光照强度0、50、100、200、400、600、800、1 000、1 200、1 400、1 600、1 800、2 000 μmol/(m2·s),记录各光照强度下的光合参数,包括光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)和水分利用效率(EWUE)等。测定后取阴阳面叶片数据的平均值,再将每个月数据计平均得到生长季平均光合数据。通过非直角双曲线模型[39]拟合光响应曲线,得到最大净光合速率(Pn,max)、光补偿点(Lcp)、光饱和点(Lsp)、暗呼吸速率(Rd)和表观量子效率(AQY)。
1.2.3 透光率的测定
针对每个林隙样地,利用半球面影像法[带有180°鱼眼镜头的Nikon CoolPix 4500数码相机(Nikon,日本)]采集半球图片,每个林隙拍摄3张照片(3次重复)。利用Digital Hemispherical Photography 4.5.2处理半球面影像,按照林隙内不同位置(中心区、过渡区和边缘区)统计各个圆环林冠空隙所占的比值(林隙分数),以此计算各林隙内不同位置的透光率,各重复样地的透光率求平均值。本研究中的林隙形状近似于椭圆形,计算公式[38]如下:
Sp=林隙分数×S;
T=Sp/S。
式中:S为林隙面积,m2;L为林隙的最长轴长度,m;W为与L垂直的最宽轴长度,m;Sp为林隙内不同位置的透光面积,m2;T为林隙透光率,%。
1.2.4 叶绿素的测定
将红松针叶密封装入带有冰袋的保温箱中带回实验室提取叶绿素,求得叶绿素a(Chl a)、叶绿素b(Chl b)、总叶绿素(Chl T)含量和叶绿素a/b(Chl a/b)值。采用20 mL体积分数为80%的丙酮溶液将0.1 g针叶置于60 ℃恒温水浴震荡锅内避光提取,用分光光度计,每月测定1次,每个样品测定3次。
1.3 数据处理
采用Excel 2010和SPSS 19.0软件进行方差分析和多重比较,并采用单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)对数据组间差异进行显著性检验(α= 0.05);图表绘制采用SigmaPlot 14.0完成;图表中的数据取平均值±标准误差。
2 结果与分析
2.1 不同大小林隙内红松针叶的光合参数
1)最大净光合速率。小兴安岭蒙古栎林林隙内红松幼树最大净光合速率(Pn,max)在林隙中心区、过渡区和边缘区均呈现中、小林隙显著高于林内和大林隙(20.0%~60.7%和7.8%~60.6%),而大林隙对其无显著影响;且中林隙显著高于小林隙(9.2%~15.1%)(图2A)。因此,中、小林隙显著提高了红松幼树光合能力,且中林隙优于小林隙。
2)光饱和点、光补偿点及表观量子效率。林隙内红松幼树光饱和点(LSP)在林隙中心区、过渡区和边缘区均呈现中、小林隙显著高于林内和大林隙(7.5%~24.5%),而中林隙仅在中心区显著高于小林隙(8.5%),但光补偿点(LCP)则表现为中、小林隙显著低于林内和大林隙(24.8%~42.4%),但二者在大林隙内均与林内相近;其表观量子效率(AQY)仅在过渡区呈现中林隙显著高于大、小林隙和林内(33.3%~45.5%)(图2B-2D)。因此,中、小林隙显著提高了红松幼树的光饱和点,且显著降低了其光补偿点。
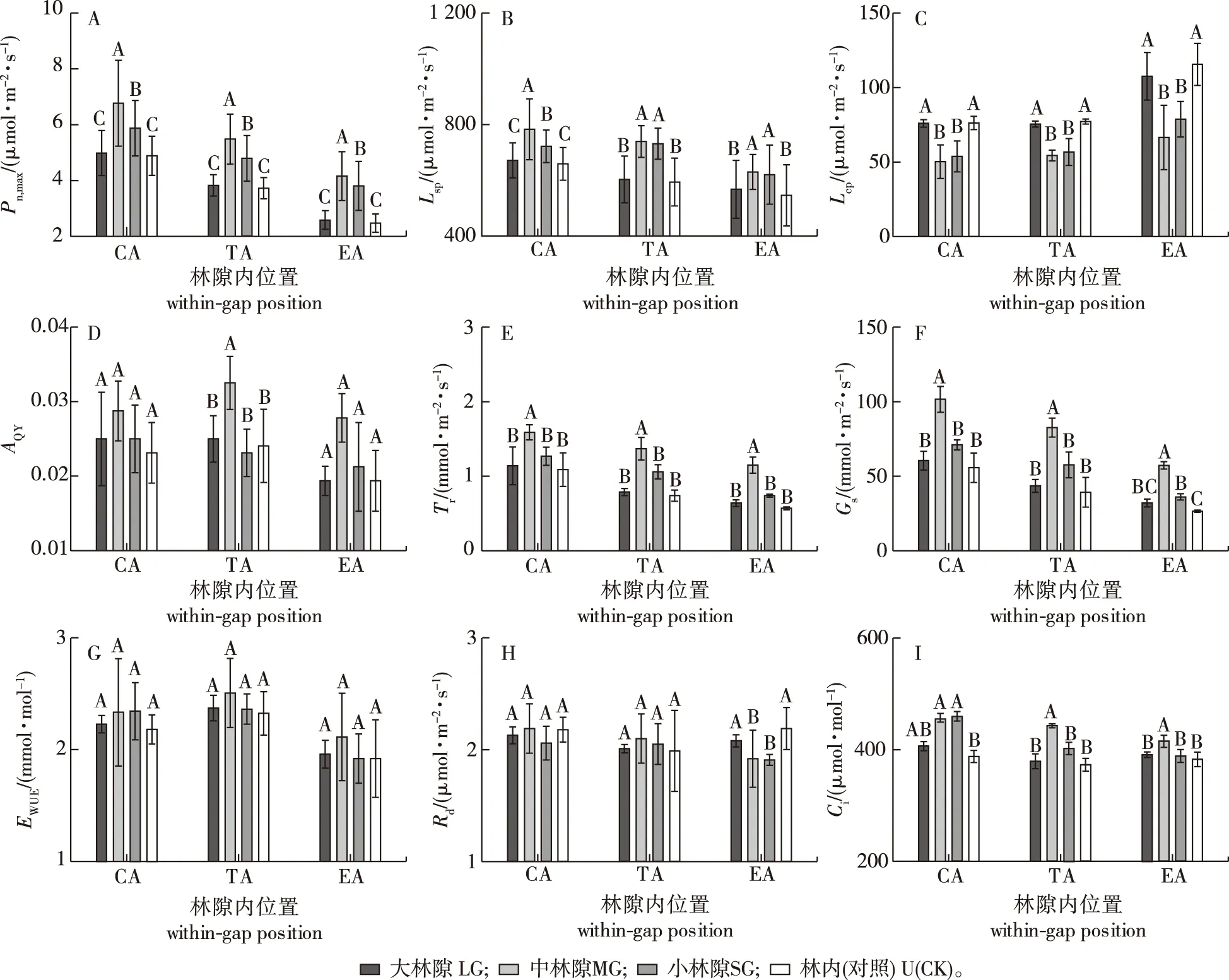
不同大写字母表示在林隙相同位置不同大小林隙间的差异显著(P<0.05)。下同。Different capital letters indicated that it has significant differences among between different gap sizes at 0.05 level at the same position of the gap.The same below.图2 小兴安岭蒙古栎林不同大小林隙内红松幼树的光合参数Fig.2 Photosynthetic parameters of the Pinus koraiensis saplings in different size gaps of the Quercus mongolica forests in Xiaoxing’an Mountains
3)蒸腾速率、气孔导度、水分利用效率、暗呼吸速率和胞间CO2浓度。林隙内红松幼树蒸腾速率(Tr)和气孔导度(Gs)在林隙中心区、过渡区和边缘区均表现为中林隙显著高于大、小林隙和林内(25.2%~101.8%和42.9%~115.5%);但水分利用效率(EWUE)在各大小林隙和林内之间均无显著差异性(图2E-2G)。因此,中林隙显著提高了林隙内红松幼树的蒸腾速率和气孔导度。林隙内红松幼树暗呼吸速率(Rd)仅在林隙边缘区呈现中、小林隙显著低于林内和大林隙(14.8%~23.5%),而胞间CO2浓度(Ci)在林隙中心区呈现中、小林隙显著高于林内(14.6%和15.7%),在过渡区和边缘区则表现为中林隙显著高于大、小林隙和林内(5.3%~15.6%)(图2H-2I)。因此,中、小林隙显著降低了林隙边缘区内红松幼树的暗呼吸速率,且中林隙显著提高了林隙内各位置红松幼树的胞间CO2浓度。
4)叶绿素含量。林隙大小对红松幼树的叶绿素a含量(Chl a)、叶绿素b含量(Chl b)和总叶绿素含量(Chl T)及叶绿素a与b质量比的值(Chl a/b)具有显著影响(图3)。红松幼树的叶绿素a、叶绿素b和叶绿素总量在中林隙内各个位置均显著低于林内和大林隙(5.6%~19.9%),而叶绿素a/b的值仅在中林隙中心区显著高于林内和大、小林隙(11.4%~11.9%)。因此,中林隙显著降低了叶绿素a、叶绿素b和叶绿素总含量,但在中林隙中心区提高了叶绿素a/b值。

图3 小兴安岭蒙古栎林不同大小林隙内红松幼树的叶绿素含量和叶绿素a/bFig.3 Chlorophyll content and radio of chlorophyll a to b of the Pinus koraiensis saplings indifferent size gaps of the Quercus mongolica forests in Xiaoxing’ an Mountains
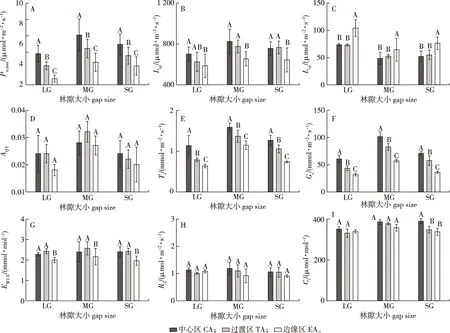
不同大写字母表示同一林隙内不同位置间的差异显著(P<0.05)。下同。Different capital letters indicated significant differences among different positions in the same gap at 0.05 level.The same below.图4 小兴安岭蒙古栎林各大小林隙内不同位置红松幼树的光合参数Fig.4 Photosynthetic parameters of the Pinus koraiensis saplings at different positions of theQuercus mongolica forests gaps in Xiaoxing’an Mountains
2.2 林隙内不同位置红松针叶的光合参数
1)最大净光合速率。林隙内红松幼树的最大净光合速率(Pn,max)在各大小林隙中心区显著高于过渡区和边缘区(22.5%~92.7%),过渡区显著高于边缘区(26.0%~47.9%)(图4A)。因此,各大小林隙内红松幼树的光合能力沿林隙中心区、过渡区至边缘区呈现出规律性递减。
2)光饱和点、光补偿点和表观量子效率。林隙内红松幼树的光饱和点(LSP)在各大小林隙内中心区显著高于边缘区(54.3%~92.7%),而仅在中、小林隙内过渡区显著高于边缘区(32.0%和26.0%);而光补偿点(LCP)在各大小林隙的中心区与过渡区显著低于边缘区(18.1%~31.6%);但表观量子效率(AQY)在各大小林隙内的各个位置之间均无显著差异性(图4B-4D)。因此,各大小林隙内中心区和过渡区红松幼树的光饱和点一般显著高于边缘区(大林隙过渡区光饱和点除外),而其光补偿点显著低于边缘区。
3)蒸腾速率、气孔导度、水分利用效率、暗呼吸速率和胞间CO2浓度。林隙内红松幼树的蒸腾速率(Tr)和气孔导度(Gs)在各大小林隙内均呈现出中心区显著高于过渡区与边缘区(16.1%~78.1% 和22.9%~96.5%),过渡区又显著高于边缘区(19.1%~43.2%和36.1%~59.3%)的递增规律性(图4E-4F)。水分利用效率(WUE)在各大小林隙内中心区和过渡区显著高于边缘区(19.0%~23.5%)(图4G)。林隙内不同位置对红松幼树的暗呼吸速率(Rd)无显著影响;而胞间CO2浓度(Ci)仅在小林隙中心区显著高于过渡区和边缘区(12.1%和15.4%)(图4H-4I)。
4)叶绿素含量。林隙内不同位置对红松幼树叶绿素a(Chl a)、叶绿素b(Chl b)和总叶绿素含量(Chl T)有显著影响,但对叶绿素a/b(Chl a/b)无显著影响(图5),其叶绿素a,叶绿素b和总叶绿素含量在大、中、小林隙内沿中心区、过渡区至边缘区均呈现出递增规律性。
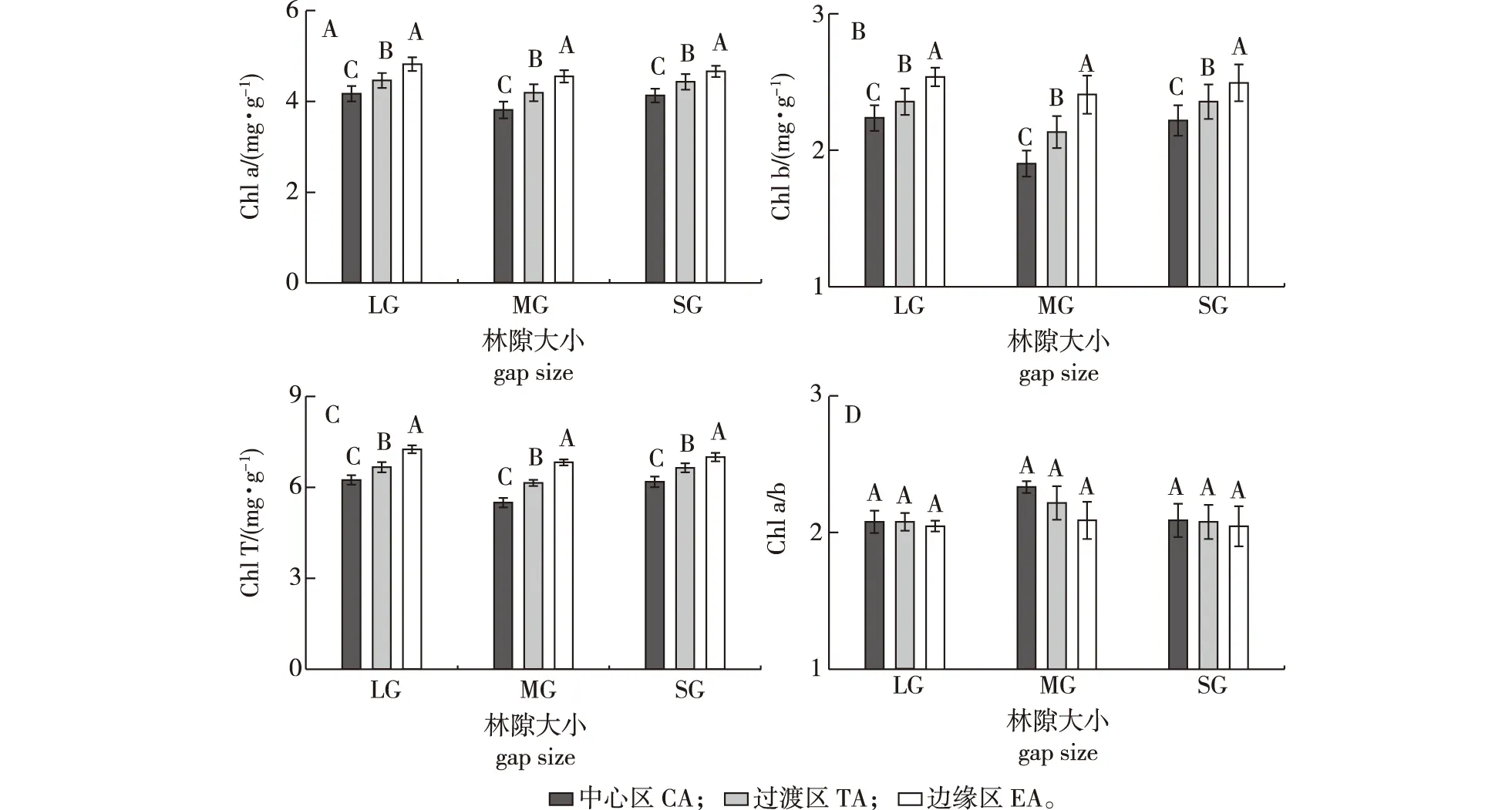
图5 小兴安岭蒙古栎林各林隙内不同位置红松幼树的叶绿素含量和叶绿素a/bFig.5 Chlorophyll content and radio of chlorophyll a to b of the Pinus koraiensis saplings at different positions of the Quercus mongolica forests gaps in Xiaoxing’an Mountains
3 讨 论
3.1 林隙大小对红松幼树光合能力的影响
光合特征参数能较好地反映植物的光合能力及其对不同光照环境的适应能力[40],本研究发现中林隙内红松幼树光合能力较强,与温带落叶林、热带雨林以及长白山原始阔叶红松林多个树种幼苗在林隙内表现出较强光合能力[11,15]的结论一致;但仅测定7月生长旺盛期红松光合速率[33]和测定整个生长季(5—10月)得到的结果不同。同时本研究证实了耐阴树种生长所需的最佳林隙林隙直径与林隙形成木树高比(D/H应小于1.0[41],但不同于伴生种蒙古栎萌生苗光合与生长需要大林隙(650~980 m2)[16],由于树种自身特性的差异,不耐阴树种蒙古栎对林隙大小的需求比具有一定耐阴性的红松更大。其原因在于林隙大小通过影响进入林隙的光量[5],改善林内光照、温度以及养分以满足植物更新与生长[1]。植物光合作用既受环境影响,也与自身条件有关[42]。Lsp、Lcp和AQY是光利用能力的重要指标,Lsp、AQY较高且Lcp较低的植物耐阴性较强[43],利用光的能力也较强[44],故红松针叶在中林隙内对强光及弱光的利用能力有所提高[31];且由于红松针叶长期适应不同大小林隙内的光照,使处于较强光照条件下的针叶表现出阳生叶片的光合特性[45],进而提高了中林隙内红松的光合能力。其次,林冠层疏开增加了直接辐射提高了林隙内气温[41],尽管不同大小林隙间气温无差异[8,46],但中林隙内较高的气温使林隙内红松幼树Tr与Gs显著增大,Gs增大最大限度地吸收光合反应原料CO2,有助于提升光合碳同化率[47],促进与外界的水汽交换,使Tr同步增强,进而光合速率提高[48-49],故红松在中林隙内光合能力最强。而大林隙并未对红松光合产生影响,原因在于林内光照过强,致使毛榛子等灌木迅速繁殖(灌丛盖度达到73.6%),灌丛呈集群分布与红松争夺养分,导致红松更新困难[50],抑制了红松光合作用。
此外,叶绿素是植物进行光合作用的主要色素,叶绿素含量增加有利于光能的吸收和转化[51-52]。红松幼树的叶绿素a、b和总叶绿素含量在林内较高,中林隙内最低,表明红松幼树主要通过增加叶绿素含量提高光能捕获能力以适应林内弱光环境,这与亚热带常绿阔叶林中赤栲(Castanopsiskawakamii)幼苗在林隙内叶绿素含量显著低于非林隙[18]一致;且衡量植物耐阴性的重要指标叶绿素a/b[53]在林内中心区最低,因为弱光条件加快了叶绿素a向叶绿素b转化[54],使叶绿素b含量增加,增强了光能捕获能力,提高其对弱光适应性。在中林隙内红松幼树的生长指标最大(胸径和树高),这与中林隙内红松幼树具有更强的光合能力相呼应,进一步证实林隙内幼苗生长与光合存在正相关[11]。
3.2 林隙内位置对红松幼树光合能力的影响
在各大小林隙内红松Pn,max沿中心区、过渡区至边缘区递减,这与阳性树种蒙古栎幼苗Pn,max在大林隙的中心位置最高[55]或在大、中林隙内随幼苗与中心距离减少而增加[16],以及中等耐阴树种胡桃楸(Juglansmandshurica)幼苗净光合速率在林隙中心附近达到峰值[56]类似,不同之处在于较耐阴树种红松同样在林隙中心区具有最高的Pn,max。其原因在于进入林隙的光量取决于林隙大小和林隙内位置[5,7],林隙内不同位置的微环境(光合有效辐射、空气与土壤的温度、湿度)存在较大差异性[57],特别是林隙中心区的光合有效辐射透光率高于边缘区[58-59],这与本研究结果一致,红松幼树长期适应这种光照异质性,其Lsp沿中心区、过渡区至边缘区随之递减,Lcp随之递增,导致红松Pn,max在各大小林隙内呈现中心区>过渡区>边缘区。
同时,在林隙中心区气温高于边缘区,相应地红松幼树的Tr与Gs在中心区也显著高于边缘区,使得红松针叶与外界的气体交换、传输能力以及光合反应原料CO2供给能力随之改变。Gs增大可以最大限度地吸收光合反应原料CO2,有助于提升光合碳同化率[47],促进与外界的水汽交换,Tr同步增强,光合速率随之提高[48-49]。可见,处于林隙内中心区的红松幼树不仅光能利用率相对较高,而且其气体交换、传输能力也较高,其Pn,max也显著提高。
此外,各大小林隙内红松幼树叶绿素a、b和总叶绿素含量沿边缘区、过渡区、中心区逐渐递减,但林隙内位置对蒙古栎萌生苗总叶绿素含量及胡桃楸和红皮云杉(Piceakoraiensis)的叶绿素含量[55]无显著影响,这可能源于林隙形成初期林隙内环境因子的变化对不同耐阴程度树种萌生叶的光合作用影响较小[54],而本研究林隙处于发育相对成熟阶段,耐阴较强的红松长期适应林隙内不同位置的光照条件,导致其叶绿素含量存在显著差异。特别是在光照较弱的林隙边缘区红松的叶绿素含量最高,这有助于吸收更多的光能来适应弱光,维持光合作用。
故在后续东北次生林恢复阔叶红松林的经营实践中,可根据林隙大小和林隙内位置合理栽植红松,以提高林隙空间利用率,例如在约为116.9 m2的林隙(D/H≈1.0)中心区群团状栽植红松或围绕已栽植的红松进行透光抚育(采伐蓄积比约为40%),更有利于红松幼树的生长与更新。
猜你喜欢
北华大学学报(自然科学版)(2022年5期)2022-11-21
格言·校园版(2022年17期)2022-07-06
小哥白尼(野生动物)(2021年9期)2022-01-17
阅读(科学探秘)(2020年8期)2020-11-06
绿色科技(2019年2期)2019-05-21
女性天地(2016年10期)2017-04-25
山西果树(2017年1期)2017-03-02
食品工业科技(2014年6期)2014-05-10
作文大王·低年级(2008年7期)2008-10-14
北京文学(2007年5期)2007-05-23
