
3个茶树品种WOX基因家族的进化及密码子偏好性比较
2022-11-30 01:02王占军吴子琦王朝霞欧祖兰蔡倩文徐忠东张照亮
南京林业大学学报(自然科学版) 2022年2期
王占军,吴子琦,王朝霞,欧祖兰,李 杰,蔡倩文,徐忠东,张照亮
(1.合肥师范学院生命科学学院,安徽 合肥 230601;2.安徽农业大学茶树生物学与资源利用国家重点实验室,安徽 合肥 230036)
随着近年来测序技术飞速发展,茶树(Camelliasinensis)‘云抗10号’(Camelliasinensisvar.assamicacv.Yunkang 10,简称CSA)[1]、‘舒茶早’(Camelliasinensisvar.sinensiscv.Shuchazao,简称CSS)[2]和‘云南野生古茶DASZ’(云南省发现的野生茶树地方品种,简称DASZ)[3]等品种的高质量全基因组相继公开,为茶树研究提供了大量基因信息,然而,这些基因却因缺少茶树转基因体系、难以通过生物技术育种改良茶树品质[4]。组织培养是植物转基因中常用的技术,但是,茶树组培体系却受限于“有限的品种”“经验式的探索”和“体系不稳定”等诸多因素,极大地制约了优良品质或遗传背景清晰的茶树品种组培体系的建立,进而限制了通过组培体系构建茶树转基因体系的研究。
WUSCHEL-related homeobox(WOX)蛋白是一类调控植物生长发育的重要转录因子,包括WUS和WOX 1~14共15个成员[5-6];其中,WUS基因是模式植物愈伤组织向不定芽分化的关键驱动器,增加WUS基因的表达量能有效促进拟南芥愈伤组织分化为不定芽[7];而WUS缺失突变体即使被置于不定芽诱导培养基中,也无法实现其愈伤组织向不定芽分化,WOX5和WOX11也参与模式植物组培过程中不定芽分化的调控[8]。值得关注的是,茶树组培是否存在模式植物WOX基因的调控机制,能否通过茶树WOX基因调控目标茶树品种的组培,进而实现构建茶树转基因体系的理想目标。然而,关于茶树WOX基因的研究,仅有‘舒茶早’全基因组中鉴定WOX基因家族并揭示其适应环境的表达特征[9],其基因功能研究仍进展缓慢。
密码子中,编码同一氨基酸的不同密码子互称同义密码子;同义密码子编码同种蛋白质时使用频率不同的现象即密码子使用偏好性[10]。转基因时,如外源基因的密码子与受体基因组的密码子偏好性不同,会诱发甲基化造成所转基因沉默或表达量低[11],因此,针对目标基因的偏好性分析及其为适应宿主密码子偏好性的优化与改造,成为转基因研究的重要前提[12]。茶树密码子偏好性研究多集中于CsICE1[13]、CsCBF1[14]和CsSAD[15]等单基因,而茶树基因家族的密码子偏好性研究却鲜见报道。本研究基于上述3个茶树品种全基因组数据,运用生物信息学方法鉴定3个茶树品种的WOX基因家族序列,对它们进行系统发育分析与密码子的使用模式、使用频率及偏好性变异来源比较研究,以期为利用茶树WOX基因构建茶树转基因体系提供重要参考信息。
1 材料与方法
1.1 数据来源
从茶树基因组数据库(www.plantkingdomgdb.com/tea_tree/)、TPIA数据库(http://pcsb.ahau.edu.cn:8080/CSS/)和Figshare数据库(https://figshare.com/search?q=DASZ)分别获得‘云抗10号’(CSA)[2]、‘舒茶早’(CSS)[3]和‘云南野生古茶树DASZ’(DASZ)[4]的CDS及氨基酸序列。
1.2 茶树WOX基因成员鉴定及生信分析方法
下载Homeodomain(PF00046)的隐马尔科夫模型[16],使用HMMER3.0软件对3个茶树品种的氨基酸序列进行检索(E值<1×10-10)[17]。从TAIR数据库下载拟南芥WOX基因的氨基酸序列;以拟南芥WOX蛋白为源序列,分别在3个茶树品种数据库中进行BLASTP检索(E值<1×10-10)[17]。删除上述方法收集的冗余序列,使用CDD、SMART和Pfam验证候选序列结构域的完整性,最终获得目标序列。运用ExPASy鉴定序列理化性质[18];利用Cell-PLoc 2.0进行亚细胞定位预测[19]。运用ClustalX 2.1比对3个茶树品种及拟南芥WOX蛋白全长序列,基于比对结果使用MEGA X软件以邻接法构建系统进化树(P-distance模式,Bootstrap检验1 000次)。综合进化分析和BLASTP比对结果命名3个茶树品种的WOX基因成员。
1.3 密码子偏好性比较分析方法
1.3.1 碱基组成分析
使用TBtools工具分别从上述茶树数据库中提取茶树WOX基因ID号对应的CDS序列;利用Perl语言编写的脚本计算CDS序列密码子的GC1(表示同义密码子中第一下标数字位的相应碱基含量。下同。)、GC2和GC3含量及3个位置的平均GC含量。
1.3.2 同义密码子相对使用度(RSCU)和同义密码子相对使用频率(RFSC)计算
同义密码子相对使用度(relative synonymous codon usage,RSCU)是指某一密码子的实际使用频率与无偏好影响下预期使用频率的比值,它能直接反映密码子的偏好性程度。由CodonW 1.4.2程序分析获得RSCU值(URSCU),若URSCU>1,则实际使用频率高于其他同义密码子;若URSCU=1,则实际使用频率与其他同义密码子相等[20]。同义密码子相对使用频率(relative frequency of synonymous codon,RFSC)是某一密码子与该密码子对应的氨基酸在样本中的实际观察值的比值。用CodonW 1.4.2程序计算RFSC值(fRFSC),当某密码子fRFSC大于同义密码子平均使用频率的1.5倍或60%时,表明该密码子偏好性较强、视为高频密码子[21]。
1.3.3 密码子使用频率比较
密码子使用频率是指某密码子使用个数占该生物编码序列总密码子个数的千分比[22]。将3个茶树品种WOX基因分别与CSA、CSS和4种模式植物[拟南芥(Arabidopsisthaliana)、烟草(Nicotianatabacum)、毛果杨(Populustrichocarpa)、水稻(Oryzasativa)]基因组进行密码子使用频率比较,探究密码子使用是否有差异;当使用频率比值介于0.5~2.0时,说明两者的密码子偏好性相似度较高,宜选为基因表达受体;反之,不宜选为基因表达受体[23]。
1.3.4 密码子使用模式影响因素分析
有效密码子数(effective number of codons,ENc)是某密码子与其同义密码子平均使用频率间的差异,其值越低密码子使用偏好越强[24],GC3s表示同义密码子中第3位的G+C含量。ENc-GC3s图以GC3s和ENc值为横纵坐标,分析碱基组成与密码子偏好性的关系。当突变压是影响密码子使用模式主因时,ENc值位于或围绕预期曲线位置;反之,若自然选择等因素决定密码子使用模式,ENc值则远离预期曲线。
奇偶偏好性分析(parity rule 2 plot,PR2-plot)是以G3/(G3+C3)和A3/(A3+T3)作横纵坐标绘制散点图,G/(G+C)表示PR2偏好类型中GC-偏倚的值,A/(A+T)表示AT-偏倚的值,图的中心代表序列内核苷酸组成跟随PR2的点,与中心的距离和方向代表PR2偏差的程度与方向,离中心越远偏倚程度越强[25]。根据A、T、G、C在图中的占比,推测碱基突变在碱基变异中的影响程度。若密码子使用仅受突变压影响,理论上密码子第3位碱基A/T和G/C的使用频率相等;否则,密码子偏好受自然选择及其他因素影响。
由此可见,概念分析法可帮助译者准确理解法律术语原语的含义,而比较法则可用于检验目标语译文含义是否与源语含义一致。若采用贴近适用者的法律术语翻译策略,译者应特别注意在翻译时除遵循内涵对等的原则外,更应注意外延含义的对等。
中性分析(neutrality plot)能定量分析突变压和自然选择等其他因素对密码子偏好的影响程度[26]。以GC12值(GC1和GC2的平均值)为纵坐标,以GC3为横坐标制作散点图,每个点代表了特定物种的每个WOX基因,绘制回归曲线。回归方程的斜率表示突变压影响密码子使用偏好的程度。回归曲线斜率为0时,表明GC12与GC3之间变异差异大,密码子偏好完全受自然选择影响;斜率为1时,说明GC12与GC3变异差异相似,突变压为影响密码子偏好的主因,自然选择的影响力缺失或很弱;且曲线斜率越靠近0或1,密码子偏好性的影响因素越靠近自然选择或突变压。
2 结果与分析
2.1 茶树WOX基因家族鉴定及进化分析
2.1.1 基因成员鉴定与序列分析
本研究最终从CSA、CSS和DASZ中分别鉴定出11、18、13个WOX成员,共计42个WOX成员。理化性质分析和亚细胞定位结果详见表1,蛋白质氨基酸长度在118~916之间,最长和最短蛋白都属于DASZ;预测等电点介于5.09~9.94,蛋白分子质量在13 333.89~102 942.02 ku之间;所有茶树WOX蛋白不稳定系数均大于40,属于不稳定蛋白;3个茶树品种的42个WOX蛋白亲水性指数均为负数,表明它们皆是亲水性蛋白;脂溶性系数均高于40,表示它们均属于热稳定蛋白。亚细胞定位结果显示,42个WOX蛋白均位于细胞核内。这与Chang等[27]揭示的12个核桃WOX蛋白亚细胞定位结果一致。

表1 3个茶树品种WOX基因家族信息Table 1 The information of WOX gene families in three Camellia sinensis cultivars
2.1.2 系统发育分析
拟南芥、CSA、CSS、DASZ的WOX蛋白的系统进化树见图1。按照Graaff等[28]分类规则,3个茶树品种的42个WOX蛋白可分为现代进化支(WUS clade)、古老进化支(ancient clade)和中间进化支(intermediate clade),该分类结果与拟南芥WOX蛋白进化研究结果相似[29]。现代进化支包括24个茶树WOX成员,其中CSA有7个,由拟南芥WOX蛋白的进化关系将其分为5个亚类,分别命名为CsaWUS、CsaWOX1、CsaWOX3、CsaWOX4、CsaWOX5;CSS包含10个,也分为5个亚类,分别命名为CssWUS、CssWOX1、CssWOX2、CssWOX3、CssWOX4;DASZ包含7个,可分为4个亚类,分别命名为DaszWOX1、DaszWOX2、DaszWOX4、DaszWOX5。中间进化支包含13个茶树WOX成员,其中CSA有4个,CSS有5个,DASZ有4个,均可分为WOX8、WOX9和WOX11这3个亚类。古老进化支含5个茶树WOX成员,CSS有3个,DASZ有2个,都属于WOX13亚类。由图1可知,同一进化支内CSA与DASZ的亲缘关系较近。

图1 拟南芥与3个茶树品种WOX基因的进化分析Fig.1 Phylogenetic analyses of WOX gene families in Arabidopsis thaliana and three Camellia sinensiscultivars
2.2 3个茶树品种密码子偏好性分析
2.2.1 碱基组成

表2 3个茶树品种WOX基因CDS序列的GC值Table 2 GC values of WOX gene families in three Camellia sinensis cultivars 单位:%
2.2.2 RSCU和RFSC结果分析与密码子使用频率比较
从3个茶树品种WOX基因密码子的URSCU与fRFSC(表3)可知,CSA、CSS、DASZ的WOX基因中URSCU>1的密码子分别有30、32、29个,其中密码子以A/T结尾的分别有22、23、24个,占偏好密码子总数的73.33%、71.88%、82.76%;3者共有的偏好密码子有25个,其中A/T结尾的密码子共20个。分析高频密码子发现,CSA中高频密码子有7个(TTG、GTT、CCT、CCA、AAG、AGA、AGG),其中A/T 结尾的有4个(GTT、CCT、CCA、AGA);CSS和DASZ各有10个高频密码子(TTG、GTG、CCT、CCA、TAA、CAC、CAA、AAG、AGA、AGG;TTG、GTT、TCT、CCT、CCA、ACT、GCT、AAG、AGA、AGG),A/T结尾的分别有5个(CCT、CCA、TAA、CAA、AGA)和7个(GTT、TCT、CCT、CCA、ACT、GCT、AGA)。3个茶树品种的高频密码子中A/T结尾密码子占比均≥50%;它们共有A/T结尾高频密码子3个(CCT、CCA和AGA),CSA与DASZ共有高频密码子7个(TTG、GTT、CCT、CCA、AAG、AGA、AGG)。在高频密码子使用模式上,CSA与DASZ相似度较高。综上所述,尽管3个茶树品种在密码子使用模式上有一定差异,但均偏好使用A/T结尾的密码子。
3个茶树品种WOX基因与6种植物间密码子使用频率比较结果(表4)可知,其中,CsaWOX与CSA和CSS的全基因组密码子相比均存在1个差异密码子(皆为编码Ala的GCG);CssWOX与CSA有1个差异密码子(编码TER的TAG),但与CSS存在1个差异密码子(编码TER的TAA),结合表3发现,TAA是CssWOX的高频密码子,因此对目标基因的表达效率影响大;DaszWOX与CSA和CSS比较均在编码TER的TAG密码子上存在明显差异;结果表明3个茶树品种WOX基因在选择CSA和CSS为转基因受体材料时,个别密码子需优化处理。
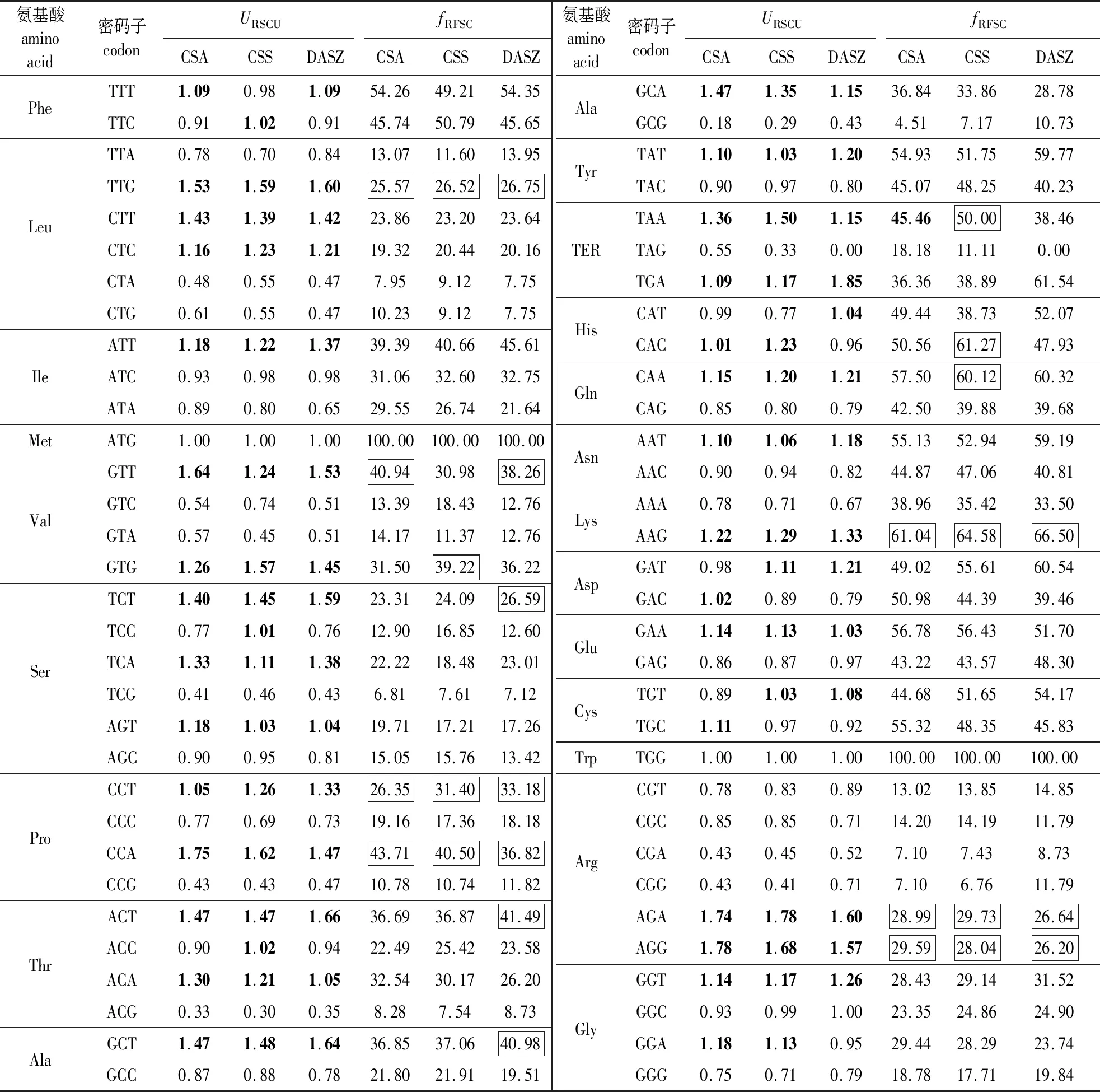
表3 3个茶树品种WOX基因密码子的URSCU和fRFSCTable 3 URSCU and fRFSC of codons of WOX gene families in three Camellia sinensis cultivars
3个茶树品种WOX基因与拟南芥密码子使用频率相比,比值≤0.5或≥2.0分别有7、4和2个;与烟草相比,密码子使用模式差异较大的分别有4、2和2个;与毛果杨相比,3个茶树品种均有6个密码子与其使用模式差异大。与水稻相比,密码子使用模式差异较大分别有13、12和12个。由此可知,4种模式植物中,烟草与3个茶树品种的密码子使用频率比较的差异最小,水稻的差异最大。3个茶树品种WOX基因与毛果杨比较,编码Leu的CTA、CTG,编码Pro的CCC和编码TER的TAA都是差异较大的密码子。在与水稻的比较中,编码Leu的CTG,编码Val的GTC,编码Pro的CCG,编码Thr的ACT、ACG,编码Ala的GCC、GCG和编码Gln的CAA均为差异较大的密码子。综上,烟草是3个茶树品种WOX基因的最佳异源表达受体,水稻最不适宜作为其基因异源表达受体。

表4 3个茶树品种WOX基因与6种植物的密码子使用频率比较Table 4 A comparison of codon usage frequency between WOX gene families in three C. sinensis cultivars and six plants

表4(续)
A.拟南芥Arabidopsisthaliana;N.烟草Nicotianatabacum;P.毛果杨Populustrichocarpa;O.水稻Oryzasativa。

A-C.ENc-GC3s图;D-F.PR2-plot作图;G-I.中性分析图 neutrality plot。图2 3个茶树品种WOX基因密码子变异来源分析图Fig.2 An analysis of the source of codon variations in WOX gene families in three Camellia sinensis cultivars
2.2.3 密码子变异来源分析
用ENc和GC3s相关性图探究影响密码子使用偏好的主要因素,其中标准曲线反映了无选择压力时二者关系(图2)。由图2A-2C可知,3个茶树品种WOX基因的ENc-GC3s图较相似,其中,CSS与DASZ的基因分布情况最相似,均多位于预期曲线下,而CSA中有2个基因落点高于预期曲线,其余9个基因落在曲线下。故3个茶树品种的大部分基因位于预期曲线下。表明影响3个茶树品种密码子使用偏好模式的主因是自然选择等其他因素,碱基突变压对密码子使用偏好存在一定影响,但影响力远小于自然选择等其他因素。
PR2偏倚分析通过探究密码子第3位上A/T与G/C之间的使用情况来确定影响密码子使用偏好模式的主因。观察图2D-2F发现,DASZ中多数基因分布于右下区域,说明在DASZ的CDS序列第3位密码子上T的使用频率高于A,G的使用频率高于C;CSS和CSA中大部分基因分布在左下,表明它们密码子第3位上T的使用频率高于A,C的使用频率高于G。由此可见,3个茶树品种A/T和G/C在密码子第3位上的使用频率存在差异,说明其密码子偏好性不仅受碱基突变压的影响,还受自然选择和其他因素影响。
GC12和GC3的中性分析可定量评估突变压和自然选择对密码子使用模式的影响程度(图2G-2I)。3个茶树品种WOX基因中性分析回归直线的斜率分别为0.065 6、0.013 7和-0.029 1,表明碱基突变压占3个茶树品种密码子使用模式6.56%、1.37%和2.91%,自然选择等因素分别占93.44%、98.63%和97.09%;表示自然选择等其他因素在3个茶树品种密码子使用偏好模式中占主导作用,而碱基突变压对密码子使用偏好的影响较小,这与ENc-GC3s和PR2-plot分析结果高度一致。
3 讨 论
本研究利用生物信息学方法在3个茶树品种中共鉴定出42个WOX基因成员(CSA11个,CSS18个,DASZ13个);其中CSS中鉴定结果与Wang等[9]研究结果一致。系统发育分析发现3个茶树品种WOX基因可分为3个大支,且每个大支都包含3个茶树品种WOX成员,表明WOX家族在进化过程中保守性较强;现代进化支包括WUS和WOX1-5,中间进化支包含WOX8、WOX9、WOX11,古老进化支只存在WOX13一个亚类。研究结果与毛白杨(Populustomentosa)[30]和番茄(Solanumlycopersicum)[31]的进化结果相似。但是,并未在3个茶树品种中发现WOX6、WOX10、WOX12和WOX14这4个亚家族,说明茶树与拟南芥的WOX基因在进化过程中存在差异。
尽管3个茶树品种在品种上存在差异,但研究结果表明它们的密码子使用模式相似性极高,即均存在密码子使用偏倚。碱基组成分析结果显示3个茶树品种WOX基因的GC含量均低于A/T含量,且都偏好A/T结尾。这与茶树CsICE1[13]、CsCBF1[14]、CsSAD[15]3个单基因的密码子偏好性和两种茶树全基因组[32](CSA和CSS)的密码子使用偏好研究结果均一致;Wang等[33]对3种麻疯树全基因组的密码子使用偏好分析也出现类似结果。从系统进化、GC含量和高频密码子使用模式的角度分析,3个茶树品种中CSA与DASZ的相似度较高,推测它们的亲缘关系更近。此外,在密码子变异来源研究中,PR2-plot分析结果表明3个茶树品种WOX基因密码子偏好性不仅受碱基突变压影响,还受自然选择和其他因素共同影响,且在苹果(Maluspumila)的奇偶偏好性分析中也出现相似结果[34];ENc-GC3s绘图定性分析了影响3个茶树品种WOX基因密码子使用偏好模式的主要因素,即自然选择等其他因素,碱基突变压对密码子使用偏好影响力小于自然选择等其他因素,Song等[35]对蒺藜苜蓿(Medicagotruncatula)研究也有类似观点;中性绘图定量分析结果显示自然选择等其他因素在3个茶树品种WOX基因密码子使用偏好模式中占主导作用,这与油棕(Elaeisguineensis)的中性分析结果[36]吻合。密码子变异来源分析结果表明,多种因素影响3个茶树品种WOX基因的密码子使用偏好模式,其中自然选择等其他因素起决定作用。同时,3个茶树品种WOX基因与CSA、CSS及4种模式生物的密码子使用频率比较发现,3个茶树品种WOX基因转CSA和CSS时个别密码子需优化,烟草与3个茶树品种的密码子使用频率比较的差异最小,即烟草是3个茶树品种WOX基因的最佳异源表达受体。而在烟草WOX基因中,NtWOX5在根尖和根特异性表达,对维持干细胞的稳定性起着重要作用,NtWOX3基因在根和幼嫩叶片中表达量较高[37]。
茶树在我国具有重要的经济价值,本研究鉴定出3个茶树品种WOX基因成员,发现3个茶树品种WOX在进化过程中存在较强保守性,揭示茶树WOX基因密码子的使用特征,借此可提高某些外源基因在茶树中的表达效率,并探究了它们的密码子偏好模式及其影响因素。研究结果为利用茶树WOX基因构建茶树转基因体系提供重要参考信息,以期利用转基因技术对茶树中特殊重要性状基因进行分子实验,从而对茶树的遗传改良实施更深入的研究。
猜你喜欢
广西植物(2022年8期)2022-09-07
茶道(2022年3期)2022-04-27
云南画报(2021年10期)2021-11-24
发明与创新·中学生(2019年6期)2019-06-26
生物学教学(2018年2期)2018-08-07
安徽农业科学(2018年1期)2018-05-14
延河·绿色文学(2016年8期)2016-05-14
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
小学生导刊(中年级)(2014年4期)2014-05-09
