
整联蛋白依赖的Akt激活在内皮细胞铺展中的作用
2022-11-27 03:18周海燕游婧璨陈妮邓鑫李永杰王立群
西南医科大学学报 2022年6期
周海燕,游婧璨,陈妮,邓鑫,李永杰,王立群
1.西南医科大学药物研究中心(泸州646000);2.西南医科大学药学院心血管药理系(泸州 646000)
血管新生是指在原有血管结构上长出新血管并形成血管网的生物学行为,其在胚胎发育、伤口愈合、肿瘤生长与转移等多种生理和病理过程中发挥重要作用[1]。内皮细胞-细胞外基质(Extracellular matrix,ECM)的相互作用以及由此介导的细胞黏附和铺展是血管新生的始动环节[2-3]。而内皮细胞铺展的分子机制仍未完全阐明。
整联蛋白又称整合素,是表达于细胞表面的ECM受体,介导细胞与细胞外基质以及细胞与细胞之间的相互识别和粘附。磷脂酰肌醇3激酶(Phosphatidyl ino⁃sitol 3-kinase,PI3K)-Akt 是ECM-整联蛋白激活的重要信号通路之一,在细胞代谢、细胞生长、细胞存活和细胞运动等过程中发挥重要调节功能[4-5]。研究表明PI3K-Akt 信号通路在ECM-整联蛋白诱导的上皮细胞[6],成纤维细胞[7]以及癌细胞[8]铺展中均起关键作用。然而,PI3K-Akt激活对内皮细胞铺展的影响仍不清楚。
纤维连接蛋白(Fibronectin,FN)是ECM 的重要组成部分,也是维持血管结构完整性的关键蛋白[9]。大量研究表明,FN及其受体在胚胎发生和血管发育中起着重要作用,FN基因缺失的小鼠伴有严重的血管发育障碍并在胚胎期死亡[10]。基于FN稳定血管结构,保护血管的生理作用,其在细胞与ECM 的相互作用研究中被广泛应用。因此,本研究将人脐静脉内皮细胞(human umbilical vein endothelial cells,HUVECs)接种于FN上,以研究Akt信号通路对内皮细胞铺展的影响,从而揭示内皮细胞铺展的潜在分子机制,为临床上治疗血管新生障碍性疾病提供新的靶点与策略。
1 材料与方法
1.1 材料
HUVECs(批号:#8000)和内皮细胞培养基(批号:#1001)购自ScienCell 公司(美国);FN(批号:F2006)购自Sigma公司(美国);磷酸化Akt抗体(批号:#4060)和总Akt抗体(批号:#4691)购自Cell Signaling Technology公司(美国);辣根过氧化物酶-山羊抗兔IgG(批号:A0208)、PI3K-Akt通路抑制剂LY294002(批号:S1737)和渥曼青霉素(Wortmannin)(批号:#S1952)购自上海碧云天生物技术有限公司;RGDS和RGES由上海昕浩生物科技有限公司合成。
1.2 细胞培养
HUVECs 培养于含5%胎牛血清、100 IU/mL 青霉素、100 μg/mL 链霉素以及1%内皮生长因子添加剂的内皮细胞培养基中,第3~10代的细胞用于实验。
1.3 细胞铺展
将FN(10 μg/mL)或PBS 加入24 孔板中,4 ℃铺板过夜,PBS 清洗3 三次,5%牛血清白蛋白室温下封闭1 h,PBS 再次清洗3 次后,接种HUVECs,加入无血清培养基继续培养30、60、90、120、180和240 min;在有些实验中,分别利用LY294002(50 μM)和渥曼青霉素(1 μM)或RGDS(200 μM)和REGS(200 μM)预处理HU⁃VECs 30 min,再将HUVECs 接种到预先铺有FN 的24孔板中培养120 min,细胞拍照,使用ImageJ 软件计算细胞面积,以评价细胞铺展。
1.4 免疫印迹
将FN(10 μg/mL)或PBS加入6孔板中,4 ℃铺板过夜,PBS清洗3次,5%胎牛血清白蛋白室温下封闭1 h,PBS 再次清洗3 次后,接种HUVECs,加入无血清培养基继续培养30、60、90、120、180和240 min;在有些实验中,分别利用LY294002(50 μM)和渥曼青霉素(1 μM)或RGDS(200 μM)和REGS(200 μM)预处理30 min后,再将HUVECs接种到FN包被的6孔板中培养1 h,提取细胞总蛋白,蛋白样品通过十二烷基磺酸钠-聚丙烯酰胺凝胶电泳分离后,转移到聚偏二氟乙烯膜上,5%脱脂牛奶室温下封闭1 h,加入抗-磷酸化Akt 抗体(1∶1 000)或抗-总Akt 抗体(1∶1 000),4°C 孵育过夜,用含0.1%吐温20的磷酸盐缓冲液洗涤之后,加入辣根过氧化物酶-山羊抗兔IgG 室温下孵育1 h,进一步洗涤之后,加入ECL 化学发光液,曝光显影,使用ImageJ 软件分析条带灰度值。
1.5 统计学分析
采用SPSS19.0 软件进行分析,计量资料以均数±标准差()表示,采用单向方差分析(One-Way ANOVA)对多组间进行比较后,进行Post Hoc 检验,LSD 法比较两组间差异,以P <0.05 为差异有统计学意义。
2 结果
2.1 FN介导内皮细胞铺展
在本研究中,我们首先评价了FN对内皮细胞铺展的影响。结果显示:当HUVECs 接种于PBS上时,细胞的铺展面积在240 min内几乎无变化;而当HUVECs接种于FN上时,细胞的铺展面积以时间依赖的方式逐渐增加,在120 min 时达到峰值,并且在各个时间点都显著高于PBS组,P<0.01,差异有统计学意义(图1),表明FN显著促进内皮细胞铺展。

图1 FN对细胞铺展的影响Figure 1 The effects of FN on cell spreading
2.2 FN 通过激活PI3K-Akt 信号通路介导内皮细胞铺展
为了探究内皮细胞铺展的潜在分子机制,我们检测了FN 对Akt 磷酸化水平的影响。结果显示:当HU⁃VECs接种于PBS上,Akt的磷酸化水平在120 min内基本无变化;而当HUVECs 接种于FN 上15 min,Akt 的磷酸化水平与对照组相比增加了93.8%,并呈现时间依赖性的升高,60 min时达到峰值,在各个时间点都显著高于PBS组,P<0.05,差异有统计学意义(图2),表明FN介导Akt 激活。随后,我们分别利用PI3K-Akt 信号通路抑制剂LY294002 和渥曼青霉素预处理HUVECs 30 min,再将其接种于FN 包被的24 孔板中,Western blot的结果显示:LY294002 和渥曼青霉素都显著抑制了Akt的磷酸化,P<0.05,差异有统计学意义(图3A,B)。而计算细胞铺展面积,发现与FN组相比,LY294002和渥曼青霉素预处理组细胞铺展面积都显著降低,差异有统计学意义,P<0.05(图3C,D),表明FN通过PI3KAkt信号通路介导内皮细胞铺展。

图2 FN介导Akt激活Figure 2 FN induces Akt activation

图3 FN通过PI3K-Akt信号通路介导细胞铺展Figure 3 FN induces endothelial cell spreading through PI3K-Akt signaling pathway
2.3 内皮细胞铺展需要整联蛋白依赖的Akt激活
整联蛋白是表达于细胞表面的ECM 受体,ECM-整联蛋白的相互作用激活细胞内多种信号传导通路。为了明确FN 介导的Akt 激活是否需要整联蛋白,我们检测了整联蛋白抑制剂RGDS 短肽或对照短肽RGES对Akt 磷酸化水平的影响,结果发现:RGDS 显著抑制了FN介导的Akt磷酸化,RGDS组Akt磷酸化水平减少了66.1%,P <0.05,差异有统计学意义;而对照短肽RGES对FN介导的Akt激活无显著影响(图4),表明FN介导的Akt 激活是整联蛋白依赖的。为了进一步探究整联蛋白依赖的Akt激活对内皮细胞铺展的影响,我们检测了整联蛋白抑制剂RGDS 短肽或对照短肽RGES预处理后,细胞铺展面积的变化,结果显示:与FN组相比,RGDS预处理组,细胞铺展面积显著降低,差异有统计学意义,P<0.05(图5),表明整联蛋白依赖的Akt激活对内皮细胞铺展是必须的。
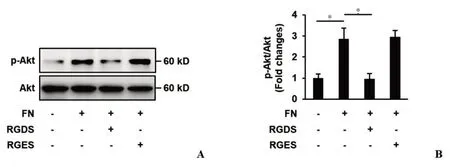
图4 整联蛋白抑制剂RGDS对FN介导Akt激活的影响Figure 4 The effects of RGDS(integrin inhibitor)on FN-induced Akt activation

图5 FN通过整联蛋白介导细胞铺展Figure 5 FN induces endothelial cell spreading through integrins
3 讨论
越来越多的证据表明,ECM 除了为细胞提供结构性支持外,在调控细胞粘附、铺展、迁移和存活等过程中也发挥关键作用[11]。FN 是细胞外基质中最早发现的非胶原糖蛋白,其功能之一是作为基底膜的组成部分调控血管网络的形成,因此对内皮细胞的生理和病理生理过程有重要影响[10]。本研究首先探讨了FN 对内皮细胞铺展的影响,结果发现FN显著促进内皮细胞铺展。之前的研究表明FN 可诱导胶质母细胞瘤细胞[12]、NIH3T3 细胞[13]和纤维母细胞瘤细胞[14]等多种细胞的铺展,这与我们的研究结果一致。并且有文献报道:细胞在FN 上铺展的早期步骤主要分为初步黏附期,细胞面积快速增长期以及缓慢铺展期三个阶段[15],结合我们的实验结果可以看出,对HUVECs而言,0~30 min为细胞初步粘附期,30~120 min为细胞面积快速增长期,120 min则进入缓慢铺展期。
Akt 是PI3K 级联反应中最经典的效应激酶之一,PI3K 诱导细胞膜磷脂酰肌醇的3'-OH 基团磷酸化,招募Akt到细胞膜上并介导其激活。有文献报道FN可以激活卵巢癌细胞[16]、脊髓星形胶质细胞[17]和人脑胶质瘤细胞[18]中的PI3K-Akt通路,而本研究发现FN以时间依赖的方式介导Akt 磷酸化,这与之前的研究结果一致。值得注意的是,FN 介导的Akt 磷酸化在60 min 时达到峰值,而FN 介导的细胞铺展在120 min 时到达平台期,这表明Akt的磷酸化与细胞铺展存在时间上的先后顺序,Akt 的激活早于细胞铺展;同时Akt 的激活在120 min时已经呈现下降的趋势,而细胞的完全铺展几乎在120 min 时完成,这表明Akt 激活和细胞铺展又存在时间上的相关性,提示FN介导内皮细胞铺展的潜在分子机制可能与PI3K-Akt激活有关。众所周知PI3KAkt 信号通路在调控内皮细胞迁移、增殖、存活以及凋亡中发挥重要作用[19-20],而我们应用PI3K-Akt 抑制剂进行干预的实验显示:LY294002和渥曼青霉素都显著抑制内皮细胞铺展,这表明FN诱导的内皮细胞铺展需要PI3K-Akt 信号的激活。尽管不同细胞的铺展可能需要Syk[21]、局部黏着斑激酶[22]以及PI3K-Akt[23-24]等一系列细胞内信号的协同作用,但使用FN作为细胞粘附底物的研究证实,PI3K-Akt 信号通路对成纤维细胞[23]和GD25 细胞[24]的铺展至关重要,这与本研究的结果一致。
整联蛋白通常由一个α亚基和一个β亚基组成,表达于细胞膜表面,作为ECM的受体与细胞外的ECM结合,不仅增强了细胞与基质的结合强度,还可以激活多种细胞内信号传导通路[25]。而对FN而言,其整联蛋白受体通常为αvβ3和α5β1,FN通过RGD 序列与细胞上的整联蛋白结合,因此细胞外给与RGDS 短肽可以抑制FN与其整联蛋白受体的结合[10,26]。我们关于RGDS对Akt激活影响的实验表明,使用RGDS短肽封闭整联蛋白可显著抑制FN诱导的Akt磷酸化,表明FN介导的PI3K-Akt信号通路激活依赖整合素。与我们的结果相一致,多篇文献证实PI3K-Akt 是FN 与整联蛋白结合后激活的信号转导通路之一[27-28]。随后,本研究进一步发现RGDS 显著抑制了FN 诱导的HUVECs 铺展,结合PI3K-Akt 抑制剂对细胞铺展影响的实验结果,表明FN通过整联蛋白依赖的Akt激活介导内皮细胞铺展。
4 结论
本研究探讨了内皮细胞铺展的潜在分子机制,发现整联蛋白依赖的Akt激活对FN介导的内皮细胞铺展至关重要。这一结果提示FN-整联蛋白-PI3K-Akt 信号通路可能是临床实践中治疗和预防血管新生疾病的潜在靶点。
(利益冲突:无)
猜你喜欢
河南畜牧兽医(2020年3期)2020-01-10
中国眼镜科技杂志(2019年9期)2019-11-11
祝您健康·文摘版(2019年10期)2019-10-14
天津医科大学学报(2019年6期)2019-08-13
分析化学(2017年12期)2017-12-25
文理导航·科普童话(2016年7期)2017-02-04
现代检验医学杂志(2016年4期)2016-11-15
中国病理生理杂志(2015年8期)2015-12-21
安徽医科大学学报(2015年9期)2015-12-16
云南中医学院学报(2014年5期)2014-07-31
