
Salvia miltiorrhiza extract may exert an anti-obesity effect in rats with high-fat diet-induced obesity by modulating gut microbiome and lipid metabolism
2022-11-25 08:16:04ZiLiAiXianZhangWeiGeYouBaoZhongHaiYanWangZhengYunZuoDuanYongLiu
World Journal of Gastroenterology 2022年43期
Zi-Li Ai, Xian Zhang, Wei Ge, You-Bao Zhong, Hai-Yan Wang, Zheng-Yun Zuo, Duan-Yong Liu
Abstract
Key Words: Salvia miltiorrhiza extract; Obesity; Gut microbiota; Lipid metabolism; High fat diet
INTRODUCTION
Obesity, a condition whose incidence increases yearly, can lead to several chronic metabolic syndromes such as diabetes, hyperlipidemia, and atherosclerosis. According to 2016 epidemiological statistics, more than 1.9 billion adults worldwide suffer from obesity, and its prevalence reaches 70% in the United States[1]. According to the WHO, adults with a body mass index (BMI) > 25 are considered overweight,while adults with a BMI > 30 are considered obese[2].
Gut microbiota (GM) is among the key regulators of metabolism[3]. Dysregulation of the GM is closely connected with obesity and its complications[4]. Under physiological circumstances, there is a symbiotic relationship between the GM and the host, which keeps the intestinal dynamic equilibrium of the body and metabolism[5]. On the other hand, the GM-host imbalance[6]has been associated with the occurrence of many diseases, such as obesity, enteritis, and colitis. Recent data indicated that there are significant differences in the structural composition of the GM of obese patients and that of non-obese people. For example, fewerBacteroides(beneficial bacteria which metabolize oligosaccharides and polysaccharides, providing nutrition to the host) and morePachydermawere found in the intestinal tract of obese patients[7]. Consistent findings suggest that obesity is associated with decreased abundance in some taxa, such asBifidobacterium,Christensenella, andAckermannia,which are considered beneficial microbes. Moreover, as a key regulator of host metabolism, the GM can influence lipid metabolism and blood and tissue lipid levels in humans and rats[8]. The GM has a major role in the fermentation of carbohydrates, fermenting carbohydrates and producing short-chain fatty acids (SCFAs), such as acetic acid, propionic acid, and butyric acid, to prevent and treat obesity and its complications. As a result,mounting evidence suggests treating obesity or obesity-related disease by improving the structure of the GM balance to regulate metabolism, particularly lipid metabolism.
Currently, bioactive substances of natural drugs are becoming increasingly popular as a new safe and effective approach to prevent and treat obesity.Salvia miltiorrhizaextract (Sal) is a traditional Chinese medicine that includes water-soluble components such as salyanolic acid, tanshinaldehyde, and comfrey acid that can inhibit early adipogenesis[9]and alleviate lipid metabolism disorders[10].Moreover, its fat-soluble components, such as dihydrodanhinone I, tanshinone IIA and IIB, and cryptosanthoxylinone can reduce glycerol release[11], promote adipocyte differentiation, and reduce the contents of triglycerides (TG) and cholesterol[12]to treat fatty liver or coronary atherosclerosis[13]. The main pharmacological effects of Sal include restraining the activation of IκB-α and NF-κB, inhibiting the oxidation of low-density lipoprotein (LDL-C) from regulating lipid metabolism processes and antioxidant effects, and improving the body’s sensitivity to insulin by activating the AMPK pathway[14]. Sal is also used to treat atherosclerosis[15], hyperlipidemia, obesity, and other dyslipidemia-related diseases. Furthermore, Wanget al[16]found that the effect of Sal in regulating hepatic steatosis may be related to the intestinal flora. Hence, this study further investigated the anti-obesity effects of Sal in rats with high-fat diet (HFD)-induced obesity and explored the underlying mechanism by focusing on the GM and lipid metabolism.
MATERIALS AND METHODS
Animals
Male Sprague-Dawley rats (aged 7-8 wk, weighing 160 g ± 20 g) were provided by the Hunan Silaike Jingda Experimental Animal Co. Ltd. (Changsha, China; Animal Certificate Number: SCXK (Xiang)2019-0004) and housed in specific pathogen-free conditions (23.0 °C ± 2.0 °C ambient temperature, 50%-60% relative humidity, and 12/12 h light/dark cycle) in Laboratory Animal Science and Technology Center of Jiangxi University of Traditional Chinese Medicine (Animal use license SYXK 2021-0007). The rats had free access to food and water throughout the experiment. The protocol (Permit Number:JZLLSC2021-236) was approved by the Jiangxi University of Chinese Medicine Animal Care and Use Committee and performed according to the guidelines prescribed by the committee. Experimental manipulation was performed after 7 d of acclimatization.
Drug
Salvia miltiorrhiza(batch number: 200701) was supplied by Baishixin Chinese Herbal Pieces Co., Ltd.(Millizhou, China). Preparation ofSalvia miltiorrhizaextract was prepared as follows: The radixSalvia miltiorrhizabunge was crushed into powder, dried at 58 °C, placed in 5000 mL round bottom flask,mixed with 75% ethanol (material-liquid ratio 1:10), soaked for 18 h, and placed in a water bath temperature 90 °C. A reflux extraction device was used to heat the reflux for 3 h, after which the filtrate was collected with four layers of gauze and left for 18 h after hot filtration.
The prepared Sal was freeze-dried with a vacuum freeze-dryer (Scientz-100F) and then smashed with zirconia beads using a mixer mill (MM 400, Retsch) at 30 Hz for 1.5 min. Then, 100 mg was dissolved with 1.2 mL of 70% methanol solution and vortexed for 30 s every 30 min 6 times, and the samples were placed in a refrigerator at 4 °C overnight. After centrifugation at 15984gfor 10 min, the extracts were filtered and then analyzed by ultra-performance liquid-chromatography tandem mass spectrometry(UPLC-MS/MS). Analytical conditions and mass spectrometry data were based on Wanget al[17]and Chenet al[18], respectively. Linear ion trap and triple quadrupole scans were acquired on a triple quadrupole-linear ion trap mass spectrometer (QTRAP) with the AB4500 QTRAP UPLC/MS/MS System operating in positive and negative ion mode (Figure 1A and B). For each period, a particular set of MRM transitions was observed by the metabolites eluted during this period (Figure 1C and D).Table 1 shows some of the metabolites identified in this study along with their metabolite numbers,integral values, and names.
Experimental design
According to previous studies[19-23], an HFD with purified ingredients and a total caloric value of 475 Kcal/100 g, with lard as the main source of fat (D12451, Research Diets Inc.) was used to induce obesity.The experiment scheme for the modeling of HFD-induced obesity in rats and drug administration is shown in Figure 2. At the initial phase, all the rats were divided into either a control group (given normal diet;n= 8) or an HFD group (given HFD;n= 40) for 7 wk and housed at an ambient temperature of 22.0 °C ± 1.0 °C. Then, the HFD groups were randomly subdivided into five groups:Control (HFD + normal saline), Sal_L (HFD + 0.675 g/kg/d Sal), Sal_M (HFD + 1.35 g/kg/d Sal), Sal_H(HFD + 2.70 g/kg/d Sal), and orlistat (HFD + 32.4 mg/kg/d). All the treatments lasted for 8 wk. The body weight was measured every 3 d and intake of food was measured per cage daily. After deducting the residual food from the initially supplied, the food intake (g/rat/wk) was determined. Rats were randomly selected from multiple cages, and cages were changed every 2 wk to control for potential cageeffects and sex/age differences.
Macroscopic observation
On the final day of the trial, all rats were euthanized with pentobarbital sodium (40 mg/kg intraperitoneally) and weighed, and the liver and fat were quickly removed and weighed after blood was drawn from the abdominal aorta. The body fat index (BFI) was calculated as follows: BFI = Total weight of fat/Body weight of rat × 100%.
Histological analysis
Fresh liver and adipose tissues were fixed with 4% polyformaldehyde (PFA) at 4 °C, embedded in paraffin, and then cut into 4-μm thick slices. Samples were then stained with hematoxylin-eosin (H&E)and observed under a light microscope to examine the histopathology according to the published criteria by Yerianet al[24]and Liewet al[25].
Determination of serum biochemical parameters
The serum was centrifuged for 15 min at 999gat room temperature. Serum TG, total cholesterol (TC),high-density lipoprotein (HDL-C), LDL-C, glucose (GLU), and free fatty acids (FFAs) were measured with a Beckmann COULTERAU480 automatic biochemical analyzer.
Enzyme-linked immunosorbent assay (ELISA)
To detect the contents of cAMP in liver tissue, and hormone-sensitive lipase (HSL) and PKA in adipose tissue, parts of the liver and adipose tissue were homogenized under slow rotation (4 °C, 30 min) in 300µL of RIPA buffer. The supernatant was obtained by centrifugation at 18759gfor 30 min. The concentrations of cAMP, HSL, and PKA were determined with commercial ELISA kits (Thermo Fisher Scientific, Waltham city, MA, United States), and the absorbance was measured at 450 nm with a microplate reader (Thermo, Varioskan, MA, United States).
Microbial diversity analysis
Intestinal contents of all rats were collected in cryopreservation tubes and preserved at -80 °C. The microbial diversity analysis was conducted using I-sanger (Majorbio Bio-Pharm Technology Co. Ltd.,Shanghai, China; www.i-sanger.com). Microbial community genomic DNA was extracted from intestinal contents using the E.Z.N.A.®soil DNA Kit (Omega Bio-Tek, Norcross, GA, United States). The DNA extract was tested on a 1% agarose gel, and the concentration and purity of the DNA were determined using a NanoDrop 2000 UV-vis spectrophotometer (Thermo Scientific, Wilmington, United States). PCR reactions were run in triplicate.
PCR products were extracted from 2% agarose gel, purified using the AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, Union City, CA, United States) according to the manufacturer’s instructions,and quantified using a Quantus™ Fluorometer (Promega, United States). Purified amplicons were equimolar and end-to-end sequenced on the MiSeq PE300 platform/NovaSeq PE250 (United States)under the standard protocols of Majorbio Bio-Pharm Technology Co. Ltd. (Shanghai, China). The raw reads were uploaded to the NCBI Sequence Read Archive (SRA).
Multiplexing of raw 16S rRNA gene sequencing reads was performed, followed by quality filtering achieved with Fast version 0.20.0 and merging with FLASH version 1.2.7. UPARSE version 7.1 was used for clustering operational taxonomic units (OTUs) with a 97 percent similarity cutoff. The RDP classifier version 2.2 was used for classification analysis of the 16S rRNA database with a confidence threshold of 70%. Calculation of the Shannon index and Sobs diversity was performed, and principal coordinates analysis (PCoA) was used to assess alpha diversity using Mothur (version v.1.30.1). The Kruskal-Wallis H test and the Wilcoxon rank-sum test were used to identify taxa that significantly differed (biomarkers)between groups (P< 0.05). I-sanger was used to perform community bar plot and heatmap analysis,correlation analysis, and co-occurrence network analysis.
LC-MS/MS analysis
A total of 50 mg of rat feces were mixed with 80 µL of methanol and 400 µL of methyl-tert-butyl ether for lipid extraction. The mixtures were vortexed for 30 s, followed by sonication for 30 min, and then precipitated at -20 °C for 30 min. The organic phase was separated by centrifugation at 18759gfor 15 min at 4 °C. Then, 350 µL of lipid extracts contained in the upper phase were transferred to EP vials and dried in a vacuum concentrator. Consequently, the lipid extract was re-dissolved in 100 µL of isopropanol:acetonitrile (1:1, v/v) solution, followed by 2 min vortexing and 5 min ultrasonication on an ice water bath. Finally, 80 µL of supernatant was transferred carefully to sample vials for LC-MS/MS analysis.
Mass spectral data were collected using a Thermo Q-Exactive Mass Spectrometer equipped with an electrospray ionization[26]source that can operate in either positive or negative ion mode. The raw data from the LC-MS analyses were imported into Lipid Search (Thermo, CA, United States) for peak detection, alignment, and identification. MS/MS fragments were used to identify the lipids. At least 80%of the lipidomic features detected in any set of samples were retained. After filtering, minimum lipid values were performed for specific samples with lipid levels below the lower limit of quantification, and each lipid profile was summed and pooled. After pooling procedures and imputation, log-transformed data were statistically analyzed to determine significant differences in metabolite levels between comparable groups.
Statistical analysis
Most of the data were statistically analyzed and figures produced using GraphPad Prism 7.0 software(San Diego, CA, United States). One-way ANOVA was performed on multiple groups, followed by Duncan’s test to analyze statistical differences. For the data on the GM, we used the online platform of the Majorbio Cloud. The Wilcoxon rank-sum test was used to analyze alpha diversity. Weighted UniFrac distances were used to generate the PCoA plots. Kruskal-Wallis rank sum tests were used to analyze species differences between groups. All results are represented as the mean ± SE.P< 0.05 was considered statistically significant.
RESULTS
Effects of Sal on body weight gain and fat accumulation in obese rats
After 8 wk of feeding, the body weight of HFD-induced obese rats was significantly higher (P< 0.05)compared to the control group (n= 8) (Figure 3A). However, the body weight decreased significantly in the Sal_H group (n= 8) from the 9thwk, and in Sal_L (n= 8) and Sal_M (n= 8) groups from the 10thwk compared with the HFD group (P< 0.05 for all). On the last day, the final body weight (Figure 3B),waistline (Figure 3C), and body fat index (Figure 3D) in the HFD group were observably increased compared with those of the control group, while Sal supplementation significantly reduced these indices without dose dependence (P< 0.05 for all). No significant difference was found among these groups in food intake (Figure 3E) (P> 0.05). The above results suggest that Sal can reduce the indices of HFD-induced obesity without affecting appetite intake.
Effects of Sal on serum lipid profiles, liver tissue, and adipose histopathology in HFD group rats
A long-time HFD intake often results in abnormal lipid metabolism[27]. Serum lipid content can reflect the lipid metabolism in the body, among which TG, TC, LDL-C, and HDL-C are the most critical indicators to measure lipid metabolism. In our study, HFD feeding led to a significant increase in serum TG (Figure 4A), TC (Figure 4B), LDL-C (Figure 4C), GLU (Figure 4E), and FFAs (Figure 4F) levels and a significant decrease in the HDL-C (Figure 4D) level (P< 0.05 for all), while Sal_L, Sal_M, and orlistat significantly reduced TG and LDL-C levels without a dose-dependent relationship compared with the HFD group (P< 0.05 for all). However, Sal_H had no significant effect on TC or HDL-C (P> 0.05).
To assess the pathological injury in liver and fat tissues, pathological samples were prepared and stained using H&E. In the liver tissue in the control group, the hepatic lobule structure was ordered and tight (Figure 4G and H). By contrast, the sample in the HFD group exhibited pitting necrosis with balloon-like changes in hepatocytes, which revealed obvious accumulation of lipid droplets in the liver of HFD-induced obese rats. However, Sal and orlistat treatment alleviated these pathological changes.These data further suggest that the HFD intervention for 7 wk can lead to obesity, while Sal administration can effectively reduce dyslipidemia and hepatic lipid accumulation caused by HFD.
H&E-stained fat tissue was observed under a microscope, which showed that the outline of fat cells and volume became larger and the arrangement was loose in the HFD group (Figure 4I and J) compared with the control group. After treatment with Sal and orlistat, both the outline and volume of fat cells were reduced. The results showed that Sal alleviates hepatic steatosis and adipocyte hyperplasia in HFD-induced obese rats.
Sal increases the activities of cAMP, PKA, and HSL in HFD rats
The PKA and HSL signaling system is involved in promoting lipolysis and has a central role in regulating metabolism in all organ systems affected by obesity. In the present study, the levels of cAMP(Figure 5A) in liver tissue, and HSL (Figure 5B) and PKA (Figure 5C) in adipose tissue of the HFD group were significantly lower than those of the control group (P< 0.05 for all); contrarily, these levels were significantly increased in HFD-induced obese rats treated with Sal and orlistat (P< 0.05vsHFD group for all). The results indicate that Sal regulates lipid metabolism by enhancing lipolysis in obese rats induced with an HFD.
Sal improves the composition of gut microbiome in HFD rats
Many studies have shown that GM dysbiosis has an essential role in the pathogenetic process of human obesity and animal obesity induced by an HFD. Fecal samples from various groups were analyzed by 16S rRNA sequencing to investigate the regulatory effect of Sal on the GM composition in HFD-induced obese rats. Twenty-four samples yielded a total of 1224808 sequencing reads. The Shannon index curve(Figure 6A), reflecting the alpha diversity of the intestinal flora, shows an adequate amount of sample sequencing data; the Venn diagram (Figure 6B) shows the overlap among the six groups at the OTU level. A total of 856, 810, 862, 889, 853, and 738 OTUs were identified in the control, HFD, Sal_L, Sal_M,Sal_H and orlistat groups, respectively. Statistical analysis of bioinformatics for OTUs at 97% similar levels found that 502 OTUs overlapped among groups. Compared with the control group, the number of OTUs in the GM was reduced in the HFD group, while Sal reversed the change. We think the main reason for the invalid effect of orlistat on OTUs of GM is that there was an anomalous sample in the orlistat group.
To determine which bacteria were improved through Sal and thus intervened in the disease progression of obesity, we analyzed the composition of GM in different groups. The community barplot analysis at the phylum level (Figure 6C) showed that the relative abundances ofProteobacteriaandActinobacteriotadecreased in the HFD group, and that ofDesulfobacterotaincreased compared with those in the control, Sal, and orlistat groups. Compared with the control group at the genus level (Figure 6D),the relative abundances ofAerococcus,Dubosiella,Psychrobacter, andnorank_f_Lachnospiraceaewere significantly decreased, and those ofQuinellaandTuricibacterwere increased in the HFD group. The community heatmap analysis at the genus level (Figure 6E) showed that the relative abundance ofBlautiawas increased in the HFD group than in the control, Sal, and orlistat groups, while those ofFacklamia,Jeotgalicoccus,NK4A214_group, andCorynebacteriumwere decreased.
Furthermore, β-diversity analysis, including non-metric multidimensional scaling (NMDS) and partial least squares discrimination analysis (PLS-DA), was used to assess the diversity variance among these six groups. NMDS (stress = 0.15) (Figure 6F) revealed that the GM composition of the HFD group was completely separated from that of the control group. Although the aggregation of the Sal_L group was similar to that of the HFD group, the two groups were significantly separated, and the aggregation of the control group was significantly enhanced as the dose of Sal increased.
PLS-DA (Figure 6G) was further used to analyze the similarity and differences among the grouped samples. It demonstrated that the species distribution of the HFD group was separated from that of each dose group of Sal and the control and orlistat groups, while the distance between Sal_M and the control group was shorter than that between the HFD and control groups. These findings suggest that Sal effectively improves GM composition in obese rats.
To further discover the regulatory effect of Sal on specific GM in obese rats, we analyzed species differences among the control, HFD, Sal_L, Sal_M, Sal_H, and orlistat groups. The genus-level Kruskal-Wallis H test bar plot (Figure 7A) showed that Sal and orlistat treatment markedly decreased the relative abundances ofQuinellaandBlautiain obese rats and significantly increased the abundances ofFacklamia,Corynebacterium,Psychrobacte, andnorank_f_Ruminococcaceae. In addition, the Wilcoxon ranksum test at the genus level showed that compared with the control, Sal_L, Sal_M, Sal_H, and orlistat groups, the relative abundances of these species in the HFD group were significantly decreased,includingFacklamia,Jeotgalicoccus,Aerococcus, andDubosiella, while those ofLactobacillus,Turicibacter,andQuinellawere increased significantly (Figure 7B-F). These findings suggest that Sal is important in treating obesity by regulating the GM composition in obese rats.
Correlation analysis of gut microbiota
To further clarify the distribution between groups and species, we analyzed the correlation of species abundance information among different samples through network analysis to obtain the coexistence relationship of species in environmental factors. The association and model prediction by network analysis at the genus level (Figure 7G) showed that the top species were most closely related to these six groups according to the degree of weighting (Table 2):Romboutsia, Aerococcus, Turicibacter, Quinella,Facklamia,CorynebacteriumandDesulfobacterota. The evolution analysis by the phylogenetic tree at the genus level (Figure 7H) indicated that the top species of the closest consanguinity relation wereRomboutsia,Lactobacillus,unclassified_Lachnospiraceae,Aerococcus,Turicibacter, andQuinella,which is consistent with the conclusion of community bar plot analysis, indicating that Sal exerts an anti-obesity role by regulating the above bacterial flora structure.
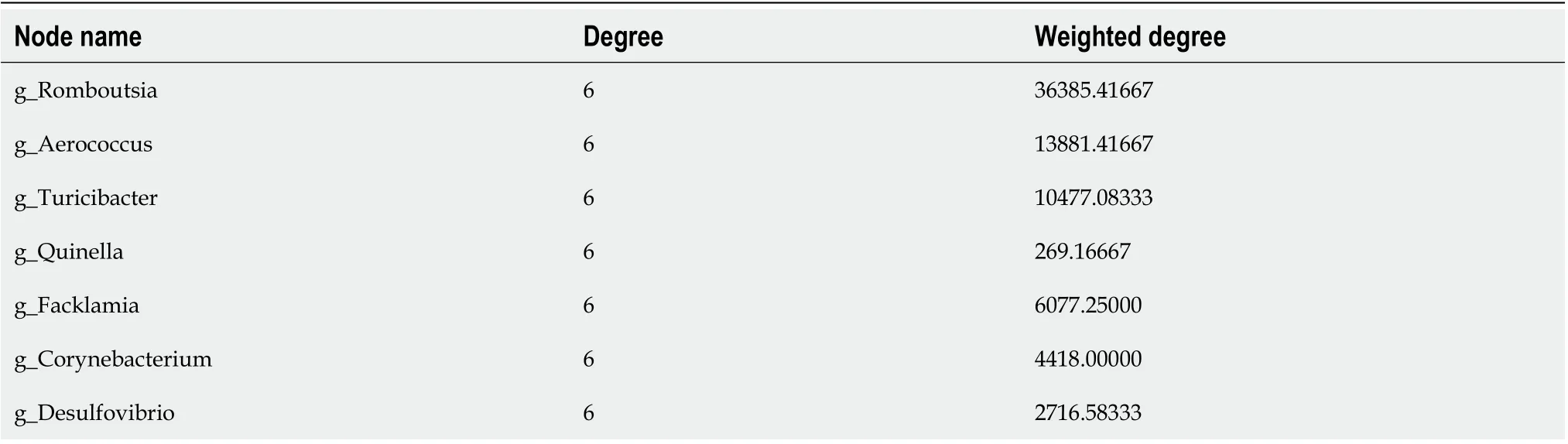
Table 2 Species nodes
Finally, the functional prediction analysis (Figure 7I) revealed that these bacteria were primarily concentrated in energy production and conversion, amino acid transport and metabolism, carbohydrate transport and metabolism, biosynthesis, transport, and catabolism of secondary metabolites, lipid transport, and lipid oxidation, according to COG and KEGG orthology information and abundance.These findings suggest that the mechanism of Sal in obesity treatment involves energy and lipid metabolism.
Sal regulates lipid metabolism in obese rats
Lipid metabolism disorders are closely related to obesity and metabolic syndrome[28]. Here, nontargeted lipidomic studies of intestinal contents in rats were conducted to explore the effects of Sal on lipid metabolism. The number of differential metabolites between the control and HFD groups and between the HFD and the Sal groups is visualized in Figure 8A. In a statistically significant analysis,different metabolites were identified whereP< 0.05 and VIP > 1 were used to identify the different metabolites. In order to understand the metabolic differences between the control, Sal, and HFD groups,the significant lipid data were analyzed using a heatmap of metabolite cluster analysis, which showed the variation of each lipid in each group directly and illustrated the relative increase (red) or decrease(blue) tendency in the HFD group compared with the control and Sal groups (Figure 8B). Interesting,the expression of the control group and the HFD group showed opposite trends; the Sal_M group was the closest to the control group trend in each Sal group, while the low and high dose groups of Sal showed the opposite trend when compared to the control group, which could be explained by the doseresponse curve. Therefore, the Sal_M group was selected to make further comparisons of differential metabolites. Relative contents of identified lipids and fold changes in metabolites in the control and Sal_M groups compared with those in the HFD group were calculated to further investigate the magnitude of change in the significant lipids (Table 3). Increased TGs (TG18:2/18:2/20:4,TG16:0/18:2/22:6, TG16:0/14:0/22:6), DGs (DG14:0/22:6, DG22:6/22:6), and CL (18:2/18:1/18:1/20:0)were observed in the HFD rats. In contrast, the lipids including ceramides (Cers; Cer d18:0/20:4, Cer d16:0/21:0, Cer d16:1/24:1), (O-acyl)-ω-hydroxy fatty acids (OAHFAs; OAHFA18:0/14:0), and Hex1Cers (Hex1Cer d18:0/16:0 + O, Hex1Cer d18:1/18:2 + 2O, Hex1Cer t18:0/16:0 + O, Hex1Cer t18:1/18:1 + 2O) showed a decreasing tendency in HFD-induced obese rats. These results identified the up-regulated and down-regulated lipids in the obesity rat induced by an HFD.
In order to further obtain the variation of the expression trend of the different metabolites among the control, HFD, and Sal_M groups, we performed VIP value analysis (Figure 8C and D). TG (16:0/14:0/22:6), TG (16:0/18:2/22:6), TG (18:2/18:2/20:4), Cer (d16:0/21:0), and DG (22:6/22:6) were the most significant metabolites among these groups. These metabolites are closely related to the development of obesity and might have an important role in the metabolism dysfunction in HFD-induced obesity.
In pathway analysis, matched metabolic pathways were displayed based on thePvalue and KEGG pathway enrichment analysis (Figure 8E). A total of 7 pathways were found withP< 0.05, five of which were closely associated with obesity, including regulation of lipolysis in adipocytes, insulin resistance,glycerolipid metabolism, fat digestion and absorption, and cholesterol metabolism. In general, these results demonstrate that Sal may exert an anti-obese effect by regulating lipid metabolism in obese rats.
The above results show that Sal can effectively regulate the blood lipid level and GM composition of obese rats. The distance-based redundancy analysis (db-RDA analysis) and Spearman correlation heatmap were used further to investigate the correlation between lipid metabolism and the GM. Three sets of environmental factors were selected to establish the correlation between lipid metabolism and GM, including the physiological and biochemical indexes (FFAs, weight, BFI, and GLU), the key factors of lipid metabolism (cAMP, PKA, and HSL), and the lipid molecules screened by lipidomics. Accordingto the db-RDA analysis, the levels of FFAs, weight, BFI, and GLU of obese rats in the HFD group were opposite to the GM abundance in the control, Sal_L, Sal_M, and Sal_H groups at the genus level(Figure 8G). Analysis of the Spearman-related heatmap (Figure 8F) at the genus level found that TGs,DGs, and Cers were positively correlated withQuinella,Desulfovibrio,Blautia, andTuricibacter, and negatively correlated withAerocuccus,Dubosiella,Facklamia, andJeotgalicoccus; cAMP, PKA, and HSL were positively correlated withCorynebacterium,Aerocuccus,Dubosiella, andFacklamiaand negatively correlated withDesulfovibrioandBlautia(Figure 8H), which agreed with our previous findings about the structure of the microbiome. The above analysis hints that Sal participates in the regulation of the microbial community structure and environmental characteristics, as well as the regulation of lipid metabolism, which helps to promote lipolysis and prevent adipogenesis.

Table 3 Differential metabolites in feces after Sal treatment
DISCUSSION
In the present study, the weight losing effect of Sal on HFD-induced obese rats is evident, which was proven by the decrease in weight and BFI without changing food intake compared to the HFD groups with increased weight, obesity indexes, and lipid profiles. Moreover, we observed that Sal significantly down-regulated the levels of TG, TC, LDL-C, GLU, and FFAs in the serum of obese rats. Additionally,H&E analysis revealed that the liver tissue from HFD-induced obese rats treated with Sal displayed reduced mean adipocyte size, increased smaller adipocytes, and alleviated hepatic steatosis. Sal can improve dyslipidemia and reduce obesity by preventing the buildup of excess lipid compositions in the liver that can cause fatty liver and dyslipidemia in obese people. Similarly, after Sal intervention,smaller adipocytes were observed in adipose tissue. These results suggest that Sal has promising antiobesity effects associated with reducing HFD-induced body weight independent of food intake.
Sal has been proven to have anti-inflammation[29]and antioxidation effects, which is widely used to treat cardiovascular disease[30], diabetes[31], fatty liver disease[32], and other dyslipidemia diseases[33]. Junget al[9]reported that tanshinone, an ingredient in the Sal, can inhibit HFD-induced obesity by preventing early adipogenesis and improving lipid metabolism. Importantly, we found that Sal exerts anti-HFD-induced obesity effects by improving blood lipid levels, and regulating GM and metabolites,thus further suggesting the potential of Sal for clinical treatment of obesity.
As an environmental factor, the GM interaction with the host[34]has an essential role in the occurrence and development of obesity[35]. Previous studies reported lower GM diversity in obese mice than in the control group, and obese mice showed a decrease inBacteroidesand an increase inFirmicutes[36,37]. Armougomet al[38]found moreMethanobacter,Bacteroidetes, andLactobacillusin anorexic patients. GM imbalance is one of the pathogeneses of obesity. In this study, we found that obese rats had a different gut flora composition than lean rats, with higher abundances ofDesulfobacterota,Quinella,andTuricibacterand lower abundances ofJeotgalicoccus,NK4A214,Aerococcus,Dubosiella,Psychrobactor,norank_f_Lachnospiraceae, andnorank_f_Erysipelotrichaceaeat the genus level andProteobacteriaandActinomycetesat the phylum level. Studies have suggested thatFirmicutes, includingLachnoaceae,Erysipelotrichaceae,Oscillospiraceae, andRuminococcaceae, can promote energy absorption in obese and diabetic rats[39]. In this study, we found that Sal decreased the abundance ofFirmicutes, includingnorank_f_LachnospiraceaeandDubosiella,in HFD-induced obese rats. Similar results were observed in HFD-induced obese rats treated with polyphenols and procyanidins[40,41].Bacteroidetes, a beneficial bacterium, have been reported to be negatively associated with obesity and hyperlipidemia. Previous studies have suggested that the proportion ofBacteroidetesandProteobacteriais slightly higher than that of the control population[27]. Interestingly, when HFD was used to successfully induce obesity in rats,we found an increased abundance ofBacteroidetesat the phylum level. Our results are consistent with Schwiertz’s evidence of a significant increase inBacteroidetesin overweight and obese subjects[42]. We believe that this is because factors, such as heredity, diet, and environment, may have similar or even opposite effects on the flora. Lipid factors involved in metabolism may also interfere with the structure of GM through various mechanisms. Studies have shown that FXR agonists regulate lipid metabolism and GM by increasing the presence ofAckermannand decreasing the presence ofDesulfuricansin obese mice[43], which is consistent with our findings.
One of the consequences of dyslipidemia is inefficient lipid metabolism, which in turn accelerates the development of obesity and dyslipidemia[44,45]. Lipid metabolism disorders provide the “first hit” in the progression of metabolic diseases such as obesity[46]. Hence, this study applied lipidomics to analyze the composition of lipid metabolites in obesity and observed the alteration of lipid metabolites by increasing the treatment of Sal to analyze the molecular mechanism of the anti-obesity and cholesterol-lowering effects of Sal, thus providing new insights into the treatment of obesity. A range of significant changes occurred in the lipidome with progressive obesity. Most notably, the contents of DGs, TGs, and CLs were increased in the HFD group compared to the control group, which was similar to previous studies[47]. Contrary, Sal decreased the levels of these lipid species in the lipidome profile,suggesting that Sal could improve the imbalance between HFD-induced lipid synthesis and catabolism.The concentration of FFAs in serum is related to lipid metabolism and glucose metabolism, and diseases such as diabetes and obesity can increase the concentration of FFAs. When energy metabolism is abnormal, FFAs will accelerate the accumulation of TG in hepatocytes or convert to lipid intermediates such as DGs, CLs, and Cer, depleting cellular functions and, in turn, leading to obesity and other metabolic disorder diseases[48]. Interestingly, we found that the supplementation of Sal effectively decreases the hepatic accumulation of TGs, DGs, and CLs. Evidence suggests that Cer is involved in obesity-induced metabolic disorders by various mechanisms, including inflammation, apoptosis, and autophagy. However, in our study, Cer was not entirely increased in the HFD-induced obese rats. We wondered whether Cer might interfere with obesity by participating in other pathways or whether the balance of sphingolipid (SP) metabolism, rather than Cer accumulation, is correlated with the development of obesity[49]. SP species are not only increased in obese patients but are also associated with hepatic oxidative stress, suggesting that these lipids may participate in the progression of obesity[50]. In general, TG accumulation is a sign of obesity and is positively correlated with hexosylceramide(HexCer); yet, in our experiment, Hex1Cer, which acts as a SP, was reduced in the HFD-induced obese rats and negatively correlated with TG. Part of that has to do with the small number of animals analyzed, and the role of Hex1Cer in obesity is not quite clear yet. Our results were consistent with Eisinger’s study[51]. Some researchers also believe that pharmacological blockage of glucosylceramide prevents obesity and liver steatosis[3,52,53]. In addition, we found that a few fatty acids, such as OAHFA, a novel lipid in structure with functions such as stimulating insulin secretion and improving glycolipid transportin vivo[54,55], were increased in HFD-induced obese rats treated with Sal. These lipids are structurally novel lipids that are provided. It is worth mentioning that some beneficial lipids are important to the development of obesity, such as phosphatidylcholines[56], which can regulate lipid and cholesterol metabolism[57]. Although it did not appear in our study, it is also one of the focuses of our future research.
Pathway enrichment analysis can assess the biochemical pathways involved in significantly different metabolites and their functional classification. Significant pathways enriched by KEGG, including lipolysis in adipocytes, insulin resistance, glycerolipid metabolism, fat digestion and absorption, and cholesterol metabolism, have been shown to be closely associated with the development of obesity,among which metabolism of cholesterol had the greatest significance, with aP-value of 0.0016.Moreover, the dysfunction of glycerolipid metabolism may disturb the energy metabolism of hepatocytes and adipocytes. Furthermore, obese patients are often accompanied by hormonal dysregulation, which aggravates lipid metabolic disorders. For example, as one of the anti-lipolysis hormones,insulin has an important role in reducing lipolysis by reducing cAMP concentration in adipocytes. The increase of cAMP catalyzes the hydrolysis of triacylglycerol to fatty acids and glycerol and then completes the lipolysis process, and PKA with HSL is involved[2]. Similarly, increases in FFAs can also cause insulin-mediated decreases in glucose oxidation and utilization, increase insulin resistance, and cause hyperinsulinemia and obesity. In our study, the levels of cAMP, PKA, and HSL were decreased in the HFD group compared to the control group, while GLU and FFA were increased. Spearman-related heatmap analysis further showed that cAMP, PKA, and HSL had a negative correlation with the structure of the GM in the HFD group, while FFAs, weight, BFI, and GLU were positively correlated with GM.
It is widely believed that the regulatory effect of intestinal microbiota on lipid metabolism is closely related to bile acids and short-chain fatty acids (SCFAs). According to Zhanget al[58], increasing the SCFA-producing generaBlautiaandAllobaculumcould enhance intestinal integrity and reduce the body weight in obese mice[59], thus rivaling obesity.Allobaculumis a member ofLachnospiraceae,Ruminococcaceae, andErysipelotrichaceae, a family that affects host metabolism[60]and protects body weight gain from HFD.Allobaculumcan produce butyric acid and propionic acid[61,62]. In our research, analysis of the Spearman-related heatmap showed that lipids including TGs, DGs, and CL that increased in the HFD group were inversely associated withAllobaculumand positively related toBlautia,which suggests that Sal may improveAllobaculumbut notBlautiato produce SCFAs and then regulate the GM and lipid metabolism. Acetate, propionate, and butyrate are examples of SCFAs produced by bacteria, and research showed that SCFA-induced activation of the PPAR pathway could modulate lipid metabolism by increasing energy consumption[63], reducing body weight, and decreasing liver TG accumulation[64]. It should be noted that the genusBlautia,as a member of theLachnospiraceaefamily, could produce SCFAs, which can regulate inflammation and metabolism[65]. Consequently, the observation in our research that Sal promoted a decrease inLachnospiraceaeandAllobaculumindicated that Sal likely confers beneficial effectsviamodulating the GM composition and host lipid metabolism with SCFAs, which is consistent with previous reports[66,67]. However, whetherLachnospiraceaehas a key role in the regulation of obesity and lipids still needs to be confirmed by accurate tests.
Desulfovibriohas been reported to be positively correlated with metabolism[68,69]and may increase after an HFD[70,71]. Similar to our findings,Desulfovibriowas reported to be decreased by the intervention of Sal. We also found that the improvement of gut microecology resulting in the increase of SCFAs may be responsible for the cholesterol-lowering effects of obesity. There is a reciprocal relationship between bile acids and cholesterol metabolism, which is closely related to intestinal flora.Bile acids are synthesized from cholesterol in the liver and have a role in maintaining cholesterol homeostasis and promoting lipid absorption[72]. Intestinal flora regulates lipid metabolism in the host by affecting bile acid composition. GM could influence bile acid metabolism by performing structural modifications, including oxidation, deconjugation, or hydroxylation[73]. It has been confirmed that bile acids and bile acid signaling pathways are involved in the control of plasma lipid and lipoprotein levels and that hepatic bile acid synthesis has an important role in the regulation of plasma TG levels in obese individuals[74], which is consistent with the present study that the cholesterol metabolism was most concentrated in KEGG pathway with the elevated cholesterol and TG in the serum and the significant elevation of TGs in lipidomics, while Sal may regulate the relationship between bile acids and cholesterol metabolism by reducing cholesterol accumulation and regulating lipid metabolism.
The above experimental results suggest that HFD-induced obesity causes dyslipidemia and dysregulation of GM and metabolites, accompanied by weight gain, hepatic steatosis, and abdominal fat accumulation. Importantly, we found that Sal effectively improved blood lipids and reshaped the balance of GM and lipid metabolism in obese rats (Figure 9), reversing weight gain and fat accumulation. Correlation analysis further demonstrated that Sal exerted anti-obesity effects through lipid metabolites of intestinal flora, which laid a good foundation for the subsequent study. In the following study, the key components of Sal’s anti-obesity effects will be further investigated by pharmacokinetic and high-performance liquid chromatography techniques. The key flora and metabolites of Sal’s antiobesity effects will be further demonstrated by intestinal flora transplantation and lipid metabolite supplementation, and the key targets of Sal’s anti-obesity effects will be further explored by molecular docking or protein interaction and gene enrichment and editing techniques, which will become a promising way to find new targets for obesity.
CONCLUSION
Our results showed that Sal reduces body weight, body fat index, serum lipid level, hepatic lipid accumulation, and adipocyte vacuolation in HFD-induced obese rats, which may be associated with enhanced gut integrity and improved lipid metabolism. 16s RNA sequence analysis revealed that Sal could reverse HFD-induced dysbacteriosis while LC-MS/MS analysis indicated that Sal could improve the lipid composition of HFD rats, which provides research basis and evidence to study the mechanism of Sal in the treatment of obesity.
ARTICLE HIGHLIGHTS
Research background
Obesity is a world health problem. A growing number of studies have suggested that gut microbiota is an important regulator of host metabolism, and the dysregulation or imbalance of gut microbiota (GM)is closely related to obesity and its complications. Mounting evidence suggests that improving the structure of the GM balance to regulate metabolism, particularly lipid metabolism, is a viable strategy for treating obesity or obesity-related disease.
Research motivation
Salvia miltiorrhizaextract (Sal) has shown good efficacy in experimental obese rats induced by a high-fat diet (HFD). Also, disturbances in gut microbiota have been observed in various diseases, including metabolic disease. However, few studies have explored the role of Sal on gut microbiota and lipid metabolism when treating obesity.
Research objectives
To investigate whether Sal can alleviate obesity induced by an HFD by regulating gut microbiome and lipid metabolism.
Research methods
Rats were given an HFD (with purified ingredients and a total caloric value of 475 Kcal/100 g, with lard as the main source of fat) for 7 wk, while Sal (0.675 g/1.35 g/2.70 g/kg/d) was administered for 8 wk.Serum lipid test, liver and fat tissue histopathologic examination, ELISA, 16S RNA sequencing, and LCMS/MS analysis were used to evaluate the efficacy of Sal on obesity.
Research results
Sal effectively improved blood lipids and reshaped the balance of gut microbiota and lipid metabolism in obese rats, reversing weight gain and fat accumulation caused by HFD. Correlation analysis further demonstrated that Sal exerted anti-obesity effects through lipid metabolites of intestinal flora, which laid a good foundation for the subsequent study.
Research conclusions
Sal may exert an anti-obesity effect in HFD-induced obese rats by modulating the gut microbiome and lipid metabolism.
Research perspectives
This study addressed an important topic of the development of obesity,i.e., the role of gut microbiota and lipid metabolism in the development of obesity, using an extract from a Chinese herb that has been found to have anti-obesity effects in various diseases.
FOOTNOTES
Author contributions:Ai ZL and Zhang X contributed equally to this work; Zhong YB and Ge W performed the experiments; Liu DY, Wang HY, and Zuo ZY contributed reagents/materials/analytical tools; Liu DY and Ge W analyzed the data; Ai ZL and Liu DY wrote the paper; Zuo ZY and Wang HY conceived and designed the experiments.
Supported bythe National Natural Science Foundation of China, No. 82060836; Jiangxi Province Graduate Student Innovation Special Fund Project, No. YC2021-B146; and Jiangxi University of Chinese Medicine Science and Technology Innovation Team Development Program, No. CXTD22008.
Institutional review board statement:The study was reviewed and approved by the Jiangxi University of Chinese Medicine (Formula-Pattern Research Center).
Institutional animal care and use committee statement:All procedures involving animals were reviewed and approved by the Jiangxi University of Chinese Medicine Animal Care and Use Committee (Approval No.JZLLSC2021-236).
Conflict-of-interest statement:There are no conflicts of interest to report.
Data sharing statement:No additional unpublished data are available.
ARRIVE guidelines statement:The authors have read the ARRIVE Guidelines, and the manuscript was prepared and revised according to the ARRIVE Guidelines.
Open-Access:This article is an open-access article that was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution NonCommercial (CC BYNC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is noncommercial. See: https://creativecommons.org/Licenses/by-nc/4.0/
Country/Territory of origin:China
ORCID number:Zi-Li Ai 0000-0001-5630-5960; Hai-Yan Wang 0000-0003-1089-0687; Duan-Yong Liu 0000-0003-2855-2811.
S-Editor:Chen YL
L-Editor:Wang TQ
P-Editor:Yu HG
 World Journal of Gastroenterology2022年43期
World Journal of Gastroenterology2022年43期
- World Journal of Gastroenterology的其它文章
- Integrity of the editing and publishing process is the basis for improving an academic journal’s Impact Factor
- Upper gastrointestinal endoscopic findings in celiac disease at diagnosis: A multicenter international retrospective study
- Kyoto classification of gastritis: Advances and future perspectives in endoscopic diagnosis of gastritis
- Current status of minimally invasive liver surgery for cancers
- Correction to “MicroRNA-596 acts as a tumor suppressor in gastric cancer and is upregulated by promotor demethylation”
- Differential analysis of intestinal microbiota and metabolites in mice with dextran sulfate sodium-induced colitis