
辨别性学习对条件性恐惧泛化的影响
2022-11-23 01:24赵绍晨岳洁如戎牧心杨子凡孟令怡郑希付
心理研究 2022年5期
赵绍晨 岳洁如 陈 伟 戎牧心 杨子凡 孟令怡 郑希付
(1 华南师范大学大脑、认知与教育科学教育部重点实验室,华南师范大学心理学院,华南师范大学心理应用研究中心,广东省心理健康与认知科学重点实验室,广州 510631;2 中国人民警察大学,广州 510663)
1 引言
恐惧泛化是指一个没有直接与厌恶刺激联系起来的线索, 由于它与已知威胁刺激的相似性或相关性, 引发了一种习得性恐惧反应 (Dymond et al.,2015)。 适当的恐惧泛化具有积极的进化意义,它使得个体可以迅速地对与危险刺激相似的潜在危险刺激做出反应,从而为“或战或逃”行为做好准备,避免潜在的伤害 (Dunsmoor et al., 2012)。 但是,过度的恐惧泛化被认为是焦虑障碍等心理问题的核心特征之一(Lissek et al., 2014)。
恐惧泛化一般采用辨别性条件恐惧范式, 该范式是目前研究恐惧习得、存储、泛化、提取、消退等过程的最重要的实验范式(Beckers et al., 2013; Lissek et al., 2008)。 在这一范式中,一般采用两个原本不具备引发个体情绪功能的中性刺激(条件刺激,Conditioned Stimulus, CS; 如色块或声音),以及一个厌恶刺激(非条件刺激, Unconditioned Stimulus,US; 如电击或恐惧图片)。 其中一个CS 后跟随US出现,称为CS+;一个 CS 后始终不跟随 US 出现,称为 CS-。 当 CS+多次与 US 匹配训练 (CS-US)后,CS+将逐渐习得恐惧效价, 个体将会对CS+这一原本为中性的刺激产生条件性恐惧反应,即CS+的单独呈现就可引起被试的恐惧反应(conditioned response, CR)。 在恐惧习得后,个体不仅对特定的CS+产生CR,也会对与CS+相似的中性刺激产生CR,这便是恐惧泛化,与CS+相似的中性刺激被称为泛化刺激 (generalization stimulus,GS)。
为了研究恐惧泛化的基本规律, 实验室研究通常采用简单的知觉刺激(如纯音、简单的几何图形、色调变化图形、面孔等)作为实验材料,运用条件反射范式进行恐惧泛化的研究 (Dunsmoor et al.,2017; Dunsmoor & Murphy, 2014; Dymond et al., 2015; Jasnow et al., 2017; Lissek et al.,2008)。 恐惧泛化的研究发现了峰对称梯度(Guttman & Kalish, 1956)、 峰漂移梯度(Hanson,1957)等现象,并就该类现象存在做出了许多可能的解 释 (Dunsmoor & LaBar, 2013; Struyf et al.,2015)。除峰梯度之外,被试还会表现出单调增加(如线性)的梯度,在与CS+同方向的维度终端刺激上响应 水 平 最 高 (Dunsmoor et al., 2009; Laberge,1961)。 这些结果表明,可能还存在其他影响恐惧泛化的加工机制。
恐惧泛化除了受刺激本身客观知觉特征的影响外,还受到个体知识经验(类别概念等)、认知信念等主观因素的影响。 Dunsmoor 和 Murphy(2014)研究表明,在恐惧条件反射中,泛化更可能从典型到非典型刺激,而不是从非典型到典型刺激(Dunsmoor &Murphy, 2014)。 其他研究表明,指导语操作影响参与者形成的规则,进而影响他们随后的泛化(Ahmed& Lovibond, 2015; Boddez et al., 2017; Vervliet et al., 2010)。 这些结果表明,知觉相似性只是许多可能影响泛化的因素之一。在焦虑障碍尤其是PTSD患者中, 一直存在对知觉上不相似的刺激产生恐惧反应的现象。 实验室研究也发现, 概念的相似性(Dunsmoor et al., 2011)、概念典型性(Dunsmoor &Murphy, 2014)和人工概念(Dymond et al., 2015)都对恐惧泛化产生不同程度的影响。
目前认为, 恐惧泛化是导致焦虑症发展的一种机制(Schiele et al., 2016),焦虑人格特征的个体容易通过恐惧泛化增加焦虑障碍的易感性和体验焦虑状态的频率(Raymond et al.,2017)。 研究发现恐惧泛化可能预测健康个体焦虑(或“焦虑状态”)的亚临床水平(Lenaert et al., 2014)。 更有趣的是,关于恐惧症的研究发现,与概念线索相比,知觉线索引发了更强的恐惧反应 (Shiban et al., 2016)。 本质上讲, 恐惧泛化可以看作是恐惧兴奋与恐惧抑制的平衡过程, 而焦虑障碍患者对知觉信息的快速加工可能意味着恐惧兴奋与抑制的失衡。
恐惧知觉泛化的模型理论认为恐惧泛化涉及皮层上的“快速通路”(high route)和皮层下的“慢速通路”(low route),泛化就是这两条神经通路共同作用的结果。在“快速通路”中,来自外周感受器的恐惧信息绕过感觉皮层的监控,从丘脑直接投射到杏仁核,杏仁核随即向脑岛、脑干等区域发放信号,引起植物神经系统的兴奋,迅速触发恐惧的生理反应。 在“慢速通路”中,恐惧信息被丘脑投射到高级感觉皮层,形成暂时的感觉表征(Lissek et al., 2014)。 恐惧泛化同时表现为快速的生理反应和慢速的认知判断。这对探究条件性恐惧中认知与情绪的作用机制有重要的启发意义。 LeDoux (2016)等又提出了关于恐惧产生的双系统回路,一条回路用于产生意识感觉,另一条回路用于控制对威胁的行为和生理反应。 第一种回路系统产生有意识的感觉, 而第二种回路系统在很大程度上无意识地运作 (LeDoux & Pine,2016)。恐惧情绪加工的双系统回路在一定程度上解释了主观预期与自动化的生理反应出现分离(Xu et al., 2016)。 基于此,本研究探究在正常个体的恐惧泛化中, 辨别性学习等高级认知活动对恐惧泛化的影响。
本研究采用阿拉伯数字作为条件性刺激材料,使用辨别性条件恐惧范式探究辨别学习在恐惧泛化中的作用。前人研究发现,在两位数字的数量控制加工任务中同时存在整体加工和局部加工 (陈兰等,2009)。 恐惧泛化的联结学习理论认为,在学习过程中,一些元素获得联结强度,而一些元素失去联结强度(Rescorla & Wagner,1972)。 条件反射过程中的差异性强化建立了特定刺激元素获得联结的价值。泛化是由与CS+所共有的元素决定的, 这些元素也有助于区分 CS+和 CS-(McLaren & Mackintosh,2002)。因此,本研究假设,辨别学习的过程对恐惧泛化会产生作用, 同时, 在主观预期和皮肤电导反应(skin conductance responses, SCR) 两个指标上可能存在不同的泛化反应模式。
2 方法
2.1 被试
43 名大学生(其中女性37 人),通过自愿报名的方式参加, 年龄在 18~26 周岁 (M=19.44,SD=1.96)之间,实验完成后给予一定报酬。 所有被试均为右利手, 视力正常或者矫正视力正常, 无听力障碍,无躯体疾病及精神障碍,且近期没有参加类似实验。 整个实验过程与被试招募标准均通过科研伦理委员会审核。 将被试随机分为2 组,其中符号组22人(女生 20 人),数值组 21 人(女生 17 人)。 两组被试的年龄无显著性差异,t(41)=1.14,p=0.26。实验开始前, 被试填写状态-特质焦虑量表 (State-Trait Anxiety Inventory, STAI), 两组被试在状态焦虑水平上无显著差异,t(41)=0.42,p=0.68,在特质焦虑水平上也无显著差异,t(41)=1.42,p=0.16。
2.2 实验材料
本实验中选取单位数 “4”“5”“6”; 两位数“44”“54”“55”“56”“66”;字母“AA”作为实验材料。 在符号组中,两位数“55”作为 CS+,字母“AA”作为 CS-,其余 6 个数字“4”“6”“44”“54”“56”“66”作为 GSs;在数值组中,两位数“55”作为 CS+,单位数“5”作为CS-,其余 6 个数字“4”“6”“44”“54”“56”“66”作为GSs(见表1)。 本实验的US 为施加于被试右手腕部的电击,电击强度在实验前由被试进行评定,其强度为“极端不舒服但还可以忍受”。

表1 实验材料
所有刺激材料均在电脑屏幕中间呈现。 首先在屏幕中间呈现 500ms 的注视点 “+”, 接着呈现8000ms 的 CS 或 GS。 同时在 CS 或 GS 的下方呈现提示文本,要求被试按1~9 的数字键判断数字后面跟随US 的可能性。 提示文本随着按键消失,CS 或GS 继续呈现(见图 1)。 习得阶段在 CS+呈现的最后200ms 可能伴随电击 (概率为75%),US 与CS 同时结束。 试次间的间隔 (Inter-Trial Intervals, ITI)为14~18s,平均间隔为 16s(Lissek et al., 2008)。
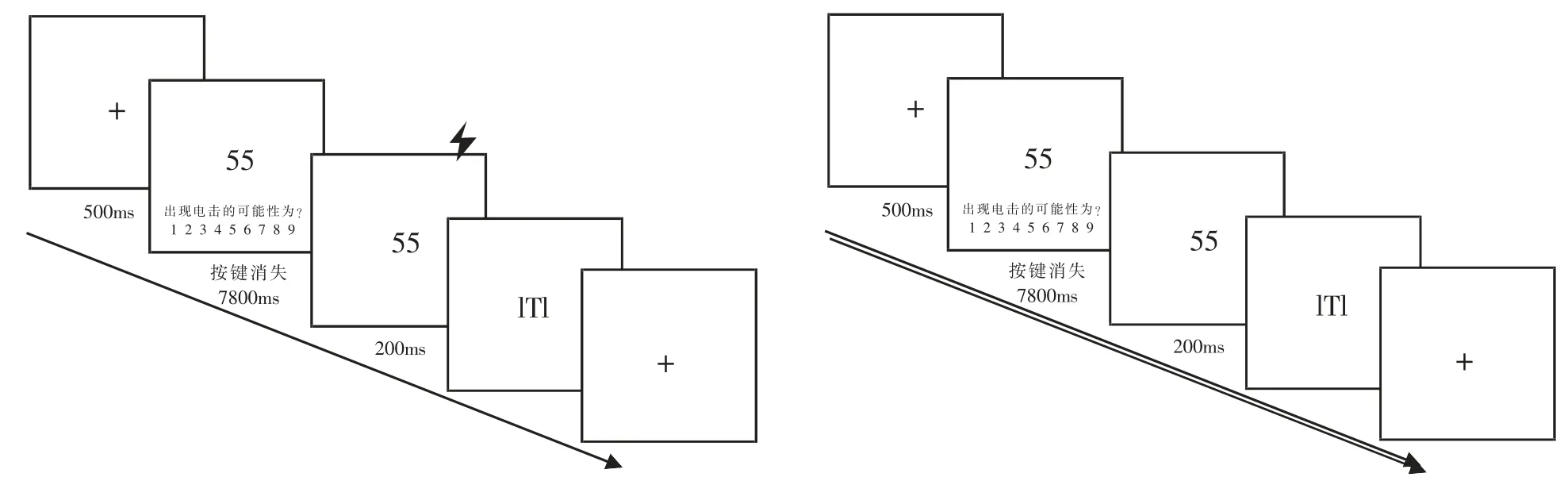
图1 刺激呈现流程图(左图为有US 呈现,右图为无US 呈现)
2.3 实验测量指标
2.3.1 US 主观预期值
在实验的各个阶段, 当CS 或GS 出现之后,下方将出现提示文本 “该刺激材料后出现电击的可能性”,对应下一行同时出现1~9 之间的9 个数字,并要求被试根据自己真实的判断用右手进行按键反应:其中1 代表完全不可能,5 代表不确定,9 代表完全可能。
2.3.2 皮电反应值
使用Biopac 16 通道生理记录仪 (型号MP150)记录被试的皮肤电。实验中将两个Ag/AgCl 电极分别缠在被试左手食指和中指的末端指腹上,电极连接在生理记录仪的EDA100C 模块上记录皮电, 采样率为1000Hz。 所采集的皮电反应数据通过Biopac 16 通道生理记录仪自带的AcqKnowledge 4.2 软件进行离线处理。对于每个被试,取CSs 或GSs 呈现后8000ms 时间窗内的最大值, 以CSs 或GSs 呈现前2000ms 时间窗内的平均值作为基线值,两者之差即为CS 或GS 诱发的原始皮电值(Milad et al., 2005)。 原始皮电值中的负数记为 0,并纳入分析(Soeter & Kindt, 2010)。基于各个被试原始皮电值中的最大值对其原始皮电值进行校准,随后进行开平方根计算以减少分布的偏态(Schultz et al., 2013)。
2.4 实验程序
参考Lissek 等人(2008)设计的经典条件性恐惧泛化研究范式,并在此基础上加入测试阶段,因此,本实验包括4 个阶段,分别为前习得阶段、习得阶段、泛化阶段和测试阶段。 程序采用E-prime 2.0 进行编程,各阶段条件刺激和泛化刺激的呈现试次情况详见表2。
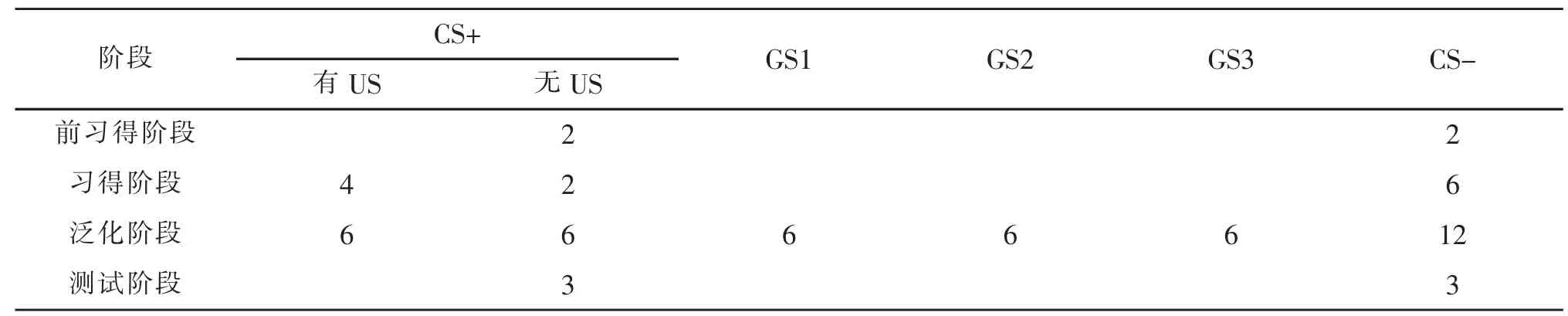
表2 各阶段条件刺激与泛化刺激的呈现次数
前习得阶段:本阶段刺激材料CS+,CS-各呈现2 次,共呈现4 次,刺激随机呈现,每个CS 不会连续两次以上重复出现。所有刺激后面都不跟随电击,让参与者熟悉操作规则。
习得阶段:本阶段CS+和CS-各呈现6 次,共呈现12 次。 其中CS+后面有75%的概率跟随电击,CS-后面始终不跟随电击。CSs 的呈现顺序以伪随机的方式呈现, 使每个刺激不会连续两次以上重复出现。此阶段目的是让被试习得条件性恐惧,若被试在测量指标上呈现对CS+与CS-反应的显著差异,则说明被试成功习得了恐惧。
泛化阶段: 本阶段采用block 设计, 分为6 个block。 每个 block 包含 10 个试次,其中 CS+,CS-各呈现 2 次,6 个 GS 各呈现 1 次。 刺激的呈现顺序以伪随机的方式呈现, 使每个刺激不会连续两次以上重复出现。 为了防止被试遗忘或习得消退, 每个block 中有 1 个 CS+后跟随 US。 而 CS-和 GSs 后面都不伴随电击。 Gene1、Gene2、Gene3、Gene4、Gene5、Gene6 分别代表被试在6 个泛化block 中对测试刺激的反应,以此研究泛化随时间的变化过程。
测试阶段: 本阶段刺激材料CS+,CS-各呈现3次,共呈现6 次,刺激以伪随机的形式呈现,泛化刺激GS 不呈现,本阶段刺激后面都不跟随电击。 以此探究与GS 相比,CS 在测量指标上的反应差异。
2.5 统计方法
对于正式实验中习得和泛化阶段所收集到的US 主观预期值和SCR 值的数据均采用重复测量方差分析。泛化阶段分别将同一类型的泛化刺激S1 和S2,S3 和 S4,S5 和 S6 进行合并,共得到三类泛化刺激 GS1,GS2,GS3。
3 结果
对被试的主观预期值和SCR 进行数据分析,以刺激类型(CS+/CS-或 CS/GSs)和刺激呈现时间分段(前期/后期)为被试内变量,以组别(符号组和数值组)为被试间变量,进行重复测量方差分析。
3.1 恐惧习得
3.1.1 主观预期值
在习得阶段,以刺激类型(CS+/CS-)、刺激呈现时间分段(前期/后期)和组别进行重复测量方差分析。 结果显示:刺激类型的主效应显著,F(1,41)=195.51,p<0.001,η2p=0.83;时间分段主效应不显著 F(1,41)=0.34,p=0.56,η2p=0.01;组别主效应不显著 F(1,41)=0.46,p=0.50,η2p=0.01;刺激类型、时间分段和组别交互作用不显著 F(1,41)=3.93,p=0.05,η2p=0.09;类型与时间分段交互作用不显著 F(1,41)=1.78,p=0.19,η2p=0.04,说明 CS+和 CS-的关系从习得前期到习得后期并没有发生显著变化, 被试在习得前期已习得了CS+和CS-的差异; 类型与组别交互作用不显著 F(1,41)=0.13,p=0.72,η2p=0.00,说明两组被试对CS+和CS-的恐惧反应不存在显著差异。 时间分段与组别交互作用不显著 F(1,41)=0.51,p=0.48,η2p=0.01,说明两组被试均在习得前期已习得 CS+和CS-的差异。 该阶段只有刺激类型的主效应显著,对CS+与 CS-进行配对样本 t 检验,结果显示,CS+显著大于 CS-,t(41)=13.98,d=2.13,p<0.001,说明被试都成功习得了恐惧,如图2 所示。
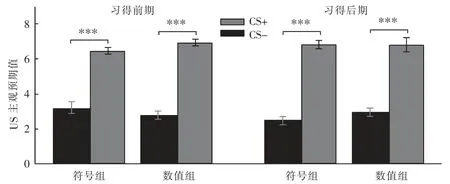
图2 习得前期、后期各组US 主观预期值
3.1.2 皮肤电导反应
在习得阶段,以刺激类型(CS+/CS-)、刺激呈现时间分段(前期/后期)和组别进行重复测量方差分析。 结果显示:刺激类型的主效应显著,F(1,41)=76.33,p<0.001,η2p=0.65; 时间分段主效应显著 F(1,41)=29.39,p<0.001,η2p=0.42;组别主效应不显著F(1,41)=2.80,p=0.10,η2p=0.06;刺激类型、时间分段和组别交互作用不显著 F(1,41)=0.12,p=0.73,η2p=0.00;类型与时间分段交互作用显著 F(1,41)=23.21,p<0.001,η2p=0.36,说明被试习得了 CS+和 CS-的差异,但从习得前期到习得后期发生显著变化; 类型与组别交互作用不显著 F(1,41)=0.01,p=0.93,η2p= 0.00,说明两组被试对CS+和CS-的恐惧反应不存在显著差异;时间分段与组别交互作用不显著 F(1,41)=0.55,p=0.46,η2p=0.01, 说明两组被试均在习得前期已习得CS+和CS-的差异。 事后检验发现,在符号组,在习得前期,CS+与 CS-差异显著,t(41)=7.02,p<0.001,在习得后期,CS+与 CS-差异显著,t(41)=3.42,p=0.03;在数值组,在习得前期,CS+与 CS-差异显著,t(41)=7.03,p<0.001, 在习得后期,CS+与 CS-差异边缘显著,t(41)=2.97,p=0.06, 说明在恐惧习得的过程中,SCR 减小,表现出了习惯化的倾向,如图3 所示。
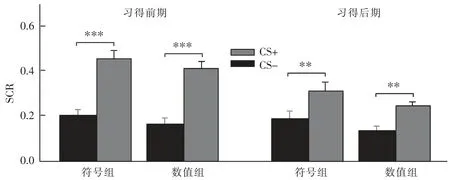
图3 习得前期、后期各组皮肤电导反应(SCR)
3.2 恐惧泛化
3.2.1 主观预期值
以刺激类型(CS+、CS-、GS1、GS2、GS3)和泛化时 间 进 程 (Gene1、Gene2、Gene3、Gene4、Gene5、Gene6)为被试内变量,组别(符号组/数值组)为被试间变量进行重复测量方差分析。
结果发现,刺激类型的主效应显著,F(4,164)=192.46,p<0.001,η2p=0.83。 事后检验表明,在刺激类型上,符号组主要表现为GS3 的泛化,而数值组主要表现为GS3 和GS2 的泛化,这表明符号组与数值组均通过威胁刺激与安全刺激的对比进行了泛化。时间进程主效应显著 F(5,205)=70.18,p<0.001,η2p=0.63。 事后检验表明,随着时间的推移,被试的泛化程度变弱,出现了泛化的消退。组别主效应不显著F(1,41)=0.00,p=0.95,η2p=0.00。 这表明,两组被试在泛化中并没有表现出差异反应。刺激类型、时间分段和组别交互作用显著 F(20,820)=4.11,p<0.001,η2p=0.09; 刺激类型与时间进程交互作用显著 F(20,820)=12.71,p<0.001,η2p=0.24;类型与组别交互作用不显著 F(4,164)=0.84,p=0.50,η2p=0.02;时间进程与组别交互作用不显著 F (5,205)=1.15,p=0.34,η2p=0.03。 简单效应分析表明,两组被试间并不存在显著性差异,分别表现为不同的泛化模式,具体表现为符号组对比CS+数值接近的泛化刺激GS3产生了泛化, 而数值组对与CS+符号特征类似的刺激GS2 和比CS+数量接近的泛化刺激GS 同时产生了泛化(图4A)。 在泛化的时间进程上,两组均表现出了泛化的消退效应(图4B)。
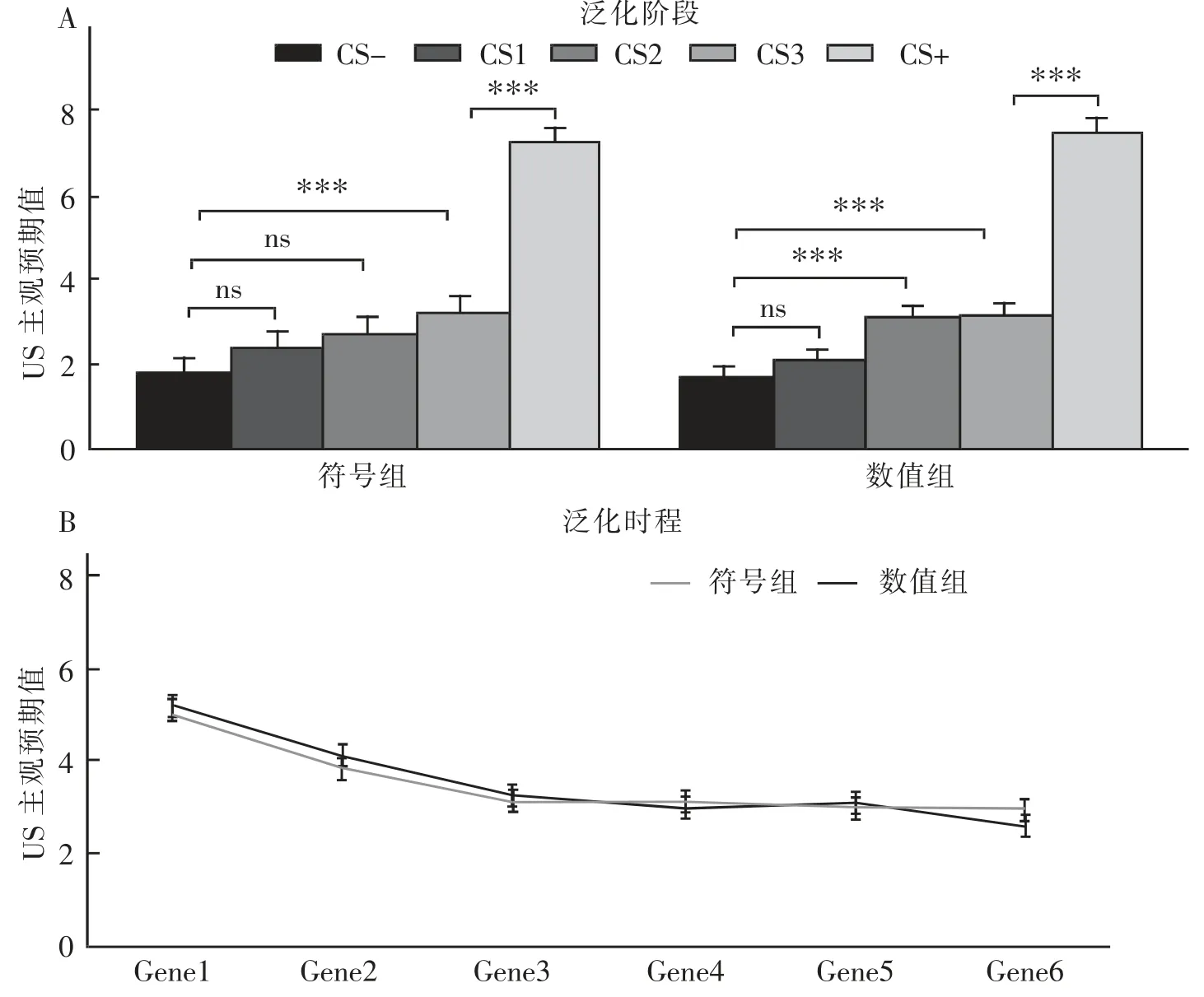
图4 A:泛化阶段刺激的主观预期值; B:主观预期值的泛化时程
3.2.2 皮肤电导反应
以刺激类型(CS+、CS-、GS1、GS2、GS3)和泛化时 间 进 程 (Gene1、Gene2、Gene3、Gene4、Gene5、Gene6)为被试内变量,组别(符号组/数值组)为被试间变量进行重复测量方差分析。
结果发现,刺激类型的主效应显著,F(4,164)=26.11,p<0.001,η2p=0.39。事后检验表明,刺激类型在SCR 上的差异主要表现为CS+与其他刺激的显著性差异, 这表明被试对诱发条件性恐惧的原始刺激CS+表现出了自动化的生理反应 (p 均小于0.001),而其他刺激与CS-的差异不显著, 并未表现出与恐惧类似的生理反应。时间进程主效应显著F(5,205)=6.31,p<0.001,η2p=0.13。 事后检验表明,Gene1 之后泛化反应消退迅速, 而其他的泛化block 之间并未出现显著差异。 组别主效应不显著 F(1,41)=0.71,p=0.41,η2p=0.02。 这表明,两组被试在泛化中并没有表现出差异反应。刺激类型、时间分段和组别交互作用显著 F(20,820)=1.61,p=0.04,η2p=0.04;刺激类型与时间进程交互作用显著 F(20,820)=1.56,p=0.06,η2p=0.04;类型与组别交互作用不显著 F(4,164)=0.85,p=0.50,η2p=0.02;时间进程与组别交互作用不显著F(5,205)=2.79,p=0.02,η2p=0.06。 简单效应分析表明,两组被试间并不存在显著性差异,并且两组内的测试刺激也并未出现泛化(图5A)。在泛化的时间进程上,两组均表现出了泛化的消退效应(图5B)。
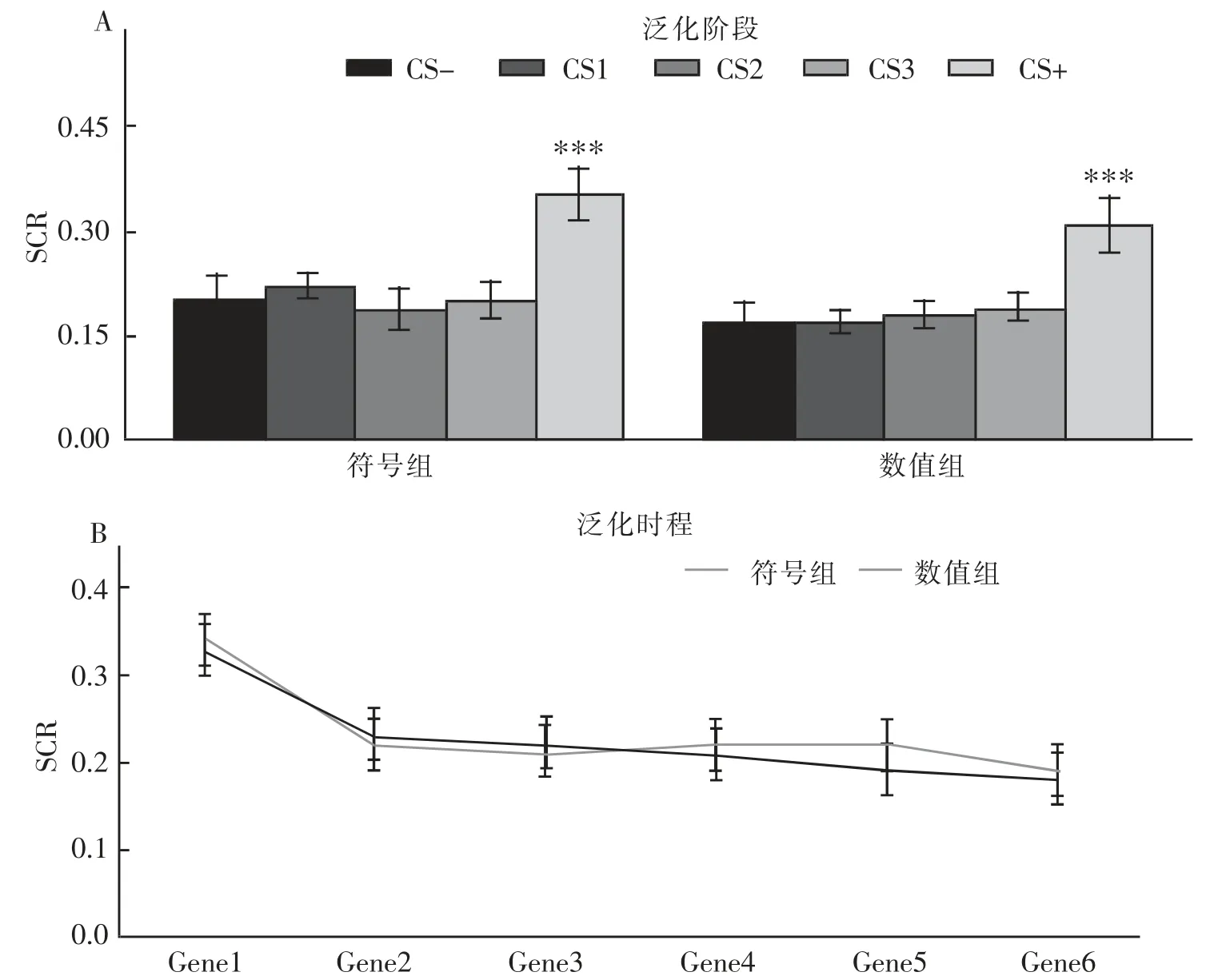
图5 A:泛化阶段刺激的SCR;B:SCR 的泛化时程
3.3 恐惧测试
3.3.1 主观预期值
在测试阶段,对刺激类型(CS+/CS-)和组别进行重复测量方差分析。结果显示,刺激类型的主效应显著,F(1,41)=182.39,p<0.001,η2p=0.82。 事后分析发现,CS+显著大于 CS-,t (41)=13.51,p<0.001,d=2.06, 这表明泛化刺激的消退并未对原始条件性恐惧刺激产生消退效果。 组别主效应不显著F(1,41)=0.26,p=0.62,η2p=0.01;刺激类型与组别交互作用不显著 F(1,41)=42.45,p<0.001,η2p=0.51,如图 6A 所示。
3.3.2 皮肤电导反应
在测试阶段,对刺激类型(CS+/CS-)和组别进行重复测量方差分析。结果显示,刺激类型的主效应显著,F(1,41)=54.92,p<0.001,η2p=0.57。事后分析发现,CS+显著大于 CS-,t(41)=7.41,p<0.001,d=1.13,这表明泛化刺激的消退并未对原始条件性恐惧刺激产生消退效果。 组别主效应不显著 F(1,41)=1.66,p=0.21,η2p=0.04;刺激类型与组别交互作用不显著 F(1,41)=1.36,p=0.25,η2p=0.03,如图 6B 所示。
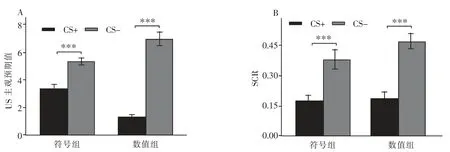
图6 A:测试阶段的主观预期值;B:测试阶段的SCR
4 讨论
本研究采用阿拉伯数字作为条件刺激, 借鉴Lissek 等人(2008)研究恐惧泛化的范式,通过操纵辨别学习中的安全刺激材料, 比较辨别学习在恐惧泛化中的作用。 结果发现,在主观预期指标上,两组被试在同等程度地习得恐惧的前提下, 符号组对数值与CS+接近的刺激产生了泛化; 而数值组对与CS+数值接近和符号特征相似的刺激均产生了泛化。 在SCR 指标上,两组被试只对CS+产生了恐惧反应,其他刺激均未引发相应的恐惧反应。
该研究结果与以往条件性恐惧中的特征学习对恐惧泛化的影响相一致(Vervliet & Geens, 2014)。恐惧的学习并非简单地与条件刺激本身有关, 还受恐惧习得个体知识经验、 认知信念和推理等主观因素的影响(Lee et al., 2019; Vervliet et al., 2010;Wong et al., 2020; Wong & Lovibond, 2017)。 本研究中,辨别性学习的差异产生了不同的泛化模式,这可能与恐惧习得过程中威胁刺激与安全刺激的对比学习有关。 前人通过辨别性条件恐惧范式操纵特征学习,选用黄色三角形作为CS+,在颜色相关组选用红色三角形作为CS-, 而在形状相关组选用黄色圆形作为CS-, 蓝色三角形和黄色的正方形作为泛化刺激。结果发现,颜色相关组对黄色正方形产生泛化, 而形状相关组对蓝色三角形产生泛化(Vervliet& Geens, 2014)。 辨别性条件恐惧习得过程中,安全刺激决定了恐惧泛化的模式。
辨别性恐惧学习中, 恐惧泛化的模式可以用刺激泛化的元素模型来解释。 巴甫洛夫条件性恐惧的元素模型认为,条件刺激由大量元素构成,这些元素能与 US 形成独立的联结(Vogel et al., 2019)。 在条件化的过程中, 不同的元素通过对US 的预期来获得或失去联结强度。 当一个从未条件化的刺激出现时, 与条件刺激类似的元素产生了US 的联结强度, 这个刺激就会引发泛化 (McLaren & Mackintosh, 2002)。值得注意的是,在 CS+与 CS-的辨别性强化过程中,只有与CS+相同的元素才获得联结值,而那些与CS-相同的元素则失去联结值。因此,不同的CS-会产生不一样的泛化模式。在当前研究中,符号组的安全刺激与威胁刺激存在共同的符号特征,使符号特征失去了与US 的联结值,因此,只有GS3引发了泛化反应; 而在数值组的安全刺激与威胁刺激并没有共同的符号特征,符号特征刺激获得与US的联结值,因此,GS2 和GS3 都引发了泛化反应。
本研究发现了在泛化阶段中主观预期值和SCR 上的分离。 恐惧泛化的双通路模型认为,个体能通过快速通路和慢速通路来习得和表达恐惧。 在“快速通路”中,来自外周感受器的恐惧信息绕过感觉皮层的监控,从丘脑直接投射到杏仁核,杏仁核随即向脑岛、脑干等区域发放信号,引起植物神经的兴奋,迅速触发恐惧的生理反应。 在“慢速通路”中,恐惧信息被丘脑投射到高级感觉皮层, 形成暂时的感觉表征, 通过比较泛化刺激与条件刺激的相似性进行泛化(Lissek et al., 2014)。 恐惧泛化同时表现为快速的生理反应和慢速的认知判断。 在恐惧习得的过程中,条件刺激直接与厌恶刺激产生联结,快速产生生理反应的同时, 也形成了对恐惧信息的认知加工,因此,恐惧表达在主观认知和客观生理反应上表现是一致的。 有意思的是,在本研究中,泛化阶段只在主观预期上对相关刺激表现出了恐惧判断,而SCR 指标上并未表现出恐惧相关的生理反应。 这表明泛化刺激并未引发相应的防御生存回路。 条件性恐惧的学习过程中存在着外显学习和内隐学习两种不同的学习系统,主观预期值体现了对CS-US 联结的外显学习,而SCR 则是个体内隐学习过程的表现(Balderston & Helmstetter, 2010; Schultz &Helmstetter, 2010)。 在以往关于知觉泛化的研究中, 泛化刺激在主观判断和客观生理反应上都引发了相关的恐惧反应。 这可能与知觉相似性刺激本身具备了与条件刺激的内隐联结有关。而在本研究中,泛化刺激并非与条件刺激存在直接的知觉相关性,因而只表现出了认知泛化。这意味着,恐惧泛化涉及不同的系统:生理活动、主观报告和显性行为(Lang,1968)。虽然实际的脑回路比双系统回路所暗示的要复杂得多, 但双系统回路对焦虑障碍的治疗有很好的启发意义。
在临床上, 暴露疗法是治疗焦虑障碍的最有效的方法之一 (Carpenter et al., 2018; Hofmann &Smits, 2008), 但暴露疗法的高复发率是实际存在的一个问题。一般来讲,暴露疗法是通过把病人重复暴露于诱发恐惧的刺激或环境中, 却不会出现任何厌恶的结果, 从而使病人关于这些恐惧相关刺激的预期发生认知冲突,达到消除恐惧的效果。这个过程只是消退了关于恐惧的认知而并未消退关于恐惧的生理反应。恐惧的网络理论模型认为,恐惧由刺激情境、刺激和反应以及刺激反应之间的意义构成。这些恐惧线索都可能重新诱发相关的恐惧反应。总之,如果仅外显或仅内隐系统被“治疗”,那么未被“治疗”的系统可以重新激发恐惧。因此,治疗失败的原因是认知防御、自主唤醒、情绪状态和错误想法对目标恐惧结构的改造产生干扰造成的。在临床上,可以尝试在暴露程序中将内隐和外显过程分开处理。
本研究在前人研究的基础上, 通过辨别性条件恐惧范式, 验证了高级认知加工在恐惧泛化中的作用, 证明了主观认知加工和客观生理反应在恐惧泛化中的不同加工模式。 但本研究尚未明确产生不同加工模式的内在机制是什么, 双通路加工模型在高级认知加工的过程中的回路尚不清楚。 关于恐惧泛化的神经机制需要未来进一步的研究。同时,如何把双通路研究的成果应用在临床治疗中也需要更多更深入的研究。
猜你喜欢
中共云南省委党校学报(2022年1期)2022-04-26
小学生优秀作文(低年级)(2020年4期)2020-07-24
今传媒(2019年10期)2019-11-11
银幕内外(2019年4期)2019-09-10
当代陕西(2019年12期)2019-07-12
现代经济信息(2019年7期)2019-06-23
特别健康(2018年9期)2018-09-26
读友·少年文学(清雅版)(2018年2期)2018-09-10
特别健康(2018年2期)2018-06-29
语言与翻译(2015年1期)2015-07-18
