
黄化茶树黄金菊的叶绿体结构透射电镜观察
2022-11-15 14:45江新凤曹挥华王治会江和源董春旺倪德江童忠飞
江西农业学报 2022年6期
江新凤,李 琛,曹挥华,王治会,江和源,董春旺,倪德江,童忠飞*
(1.江西省经济作物研究所/江西省茶叶质量与安全控制重点实验室,江西 南昌 330043;2.中国农业科学院 茶叶研究所,浙江 杭州 310008;3.华中农业大学 园艺林学学院,湖北 武汉 430070)
黄金菊是江西省选育的特异性茶树资源[1],来源于修水县宁州群体,与已经鉴定的国家良种宁州2号同属一类,为宁州群体种中表现优异的单株选育而来。尚未经过国家审(鉴、认)定,也未进行品种登记,该品种在江西种植面积较大[2],许多茶树新品系的选育均以该品系为母本或父本,该资源特色鲜明,但在全国范围内受到的关注很少,目前对于这一品种新梢黄化形成的机理尚未完全清楚[3]。江新凤等[4]在2021年对茶树资源黄金菊开展了植物学特性、适应性、适制性等方面的研究,探究了黄金菊茶树新梢黄化的生理、生化和分子机理,为江西茶树种质资源的选育提供了重要的数据资料,为光敏黄化品种栽培、种植等提供了理论基础。前人对其他作物研究表明,作物芽叶黄化通常与叶绿素合成受阻有关[5-10]。也有研究发现茶树白化性状中叶绿体结构变化较大[11-13](图1)。茶树黄金菊在正常光照情况下,待嫩梢成熟,叶色与宁州2号叶色无异,那么,黄金菊黄化的机理到底怎么样?其萌发过程中哪些亚细胞结构发生了变化?关于这些问题的研究鲜见报道。本文以宁州2号为对照,对黄化茶树黄金菊进行80%遮荫处理,待黄化茶树转为接近对照品种叶片叶色时,对各试验处理进行了叶绿素、SPAD值以及亚细胞透视电镜的观察,以期进一步揭示黄化茶树黄金菊的黄化机理。

图1 黄金菊芽叶萌发
1 材料与方法
1.1 试验地
试验茶树种植区域位于115°59′45.3″E,28°22′3.4″N,海拔50~55 m。该区域茶园土壤为红壤土,2017年测得茶园土壤pH值为4.06,有机质19.70 g/kg,全氮1137.00 mg/kg,全磷0.63 g/kg,全钾14.22 g/kg,水解性氮100.20 mg/kg,有效磷50.50 mg/kg,速效钾169.00 mg/kg,交换性镁0.42 mmol/kg,交换性钙1.20 mmol/kg。
1.2 试验茶树及取样
以黄金菊为研究材料,宁州2号为对照品种,试验茶树树龄均为20周年,长势良好。2018年4月10日开始对黄金菊茶树进行遮荫处理,遮荫措施:于茶行边每隔3 m驻直径20 mm钢管,离地面高1.8 m,处理长度15 m,宽6 m,于顶部及周边垂直到地面覆盖遮光率80%的遮荫网(用2层4针黑色遮阳网覆盖),定期观测黄金菊自然光照处理(NS)、遮荫处理(S)和对照品种宁州2号(CK)的表型性状。2018年6月10日,各试验处理间产生明显
表型性状变化,按茶树组织超微结构取样要求,采摘茶树品种自然光照黄金菊、遮荫处理黄金菊、对照品种宁州2号的第2叶、第4叶,送至中国科学院武汉病毒研究所进行TEM电镜检测,同时采用叶绿素仪分别测定各处理SPAD值(n=100),并测定各处理的叶绿素a、叶绿素b等成分含量。
1.3 测定方法
1.3.1 叶绿素含量测定 称取0.2 g茶树鲜叶(去梗去主脉),剪碎放入研钵中,加少量石英砂和碳酸钙粉,3 mL 5%乙醇,充分研磨成匀浆,再加入10 mL乙醇,充分研磨直至组织变白。避光,静置5 min后过滤到25 mL棕色容量瓶中,用乙醇定容至25 mL,摇匀。以95%乙醇为空白,分别在波长663、645、470 nm下测定吸光度[13]。
叶绿素a含量(Chla)、叶绿素b含量(Chlb)、类胡萝卜素含量(Car)的计算公式为:

式中,m为叶片质量(g)。
1.3.2 透射电镜观察 样品处理参照Wei等[14]的研究方法。待样品包埋好后,用EM UC7(德国Leica公司生产)超薄切片机切成60~100 nm的切片,铅和铀双染色后在Tecnai G220 TWIN (美国FEI公司生产)电镜下面观察。
2 结果与分析
2.1 黄金菊遮荫前后表型变化及叶绿素相对含量比较
相比较于宁州2号,黄金菊春季茶芽逐渐萌发,芽为橙红色,第1叶为浅紫色,第2叶呈现部分黄色。盛夏阶段,随着光照强度的加强,黄金菊的黄白色芽叶表型充分发展,这种现象在以往对黄金菊研究中已有报道[1-4],但是由于光照太强,有些幼龄茶树嫩梢容易出现灼伤等现象,成龄茶树或试验茶树及绿叶、对照茶树、遮荫茶树均会出现如此现象。至秋冬季节,黄金菊茶树呈现返绿现象。随着嫩梢成熟,先从嫩梢下部叶片开始返绿,自下而上叶片逐步由浅绿色返回到深绿色。通过在高于茶树树冠30~40 cm处覆盖80%黑色遮荫网,从而实现对茶树黄金菊的遮光处理,遮荫后黄金菊的表型性状的变化(图1)。结果表明,遮荫后黄金菊茶树新梢不会呈现自然光照下颜色变化的3个阶段,该试验也能证明,黄金菊茶树也属于光照敏感性茶树资源。

图2 遮荫处理后茶树的芽、叶等生长情况
由表1可知,遮荫后黄金菊第2叶的Chla含量、Chlb含量、类胡萝卜素含量、SPAD值分别为0.89 mg/g、0.23 mg/g、475.51 mg/g、31.24,比自然生长黄金菊的各指标分别提高了102.27%、155.56%、38.01%、58.90%,遮荫后色素含量呈现出显著性变化趋势(P<0.05),SPAD值提高了59.08%。遮荫处理的Chla含量、Chlb含量、类胡萝卜素、SPAD值与对照品种宁州2号的差异显著(P<0.05)。对照品种宁州2号的Chla含量、Chlb含量、类胡萝卜含量、SPAD值分别比遮荫处理提高了71.91%、130.43%、68.34%、48.43%。可见遮荫后黄金菊第2叶复绿过程中,叶绿素等物质代谢明显,但未达到宁州2号的绿化水平。

表1 试验茶树第2叶色素含量变化情况
由表2可知,遮荫处理第4叶的Chla含量、Chlb含 量、Car含 量、SPAD值 分 别 为1.99 mg/g、1.04 mg/g、936.12 mg/g、54.32,比自然生长黄金菊的各指标分别提高了36.30%、33.33%、11.74%、31.91%。遮荫处理的Car含量、SPAD值与自然光照处理的有显著性差异(P<0.05),Chla、Chlb含量在遮荫后有提高,但差异不明显。宁州2号的Chla含量、Chlb含量、Car含量、SPAD值分别与遮荫处理的相差不大,且组间无显著性差异。可见在遮荫后黄金菊第4叶复绿过程中,叶绿素等物质代谢迅速,与宁州2号第4叶绿化水平无异。
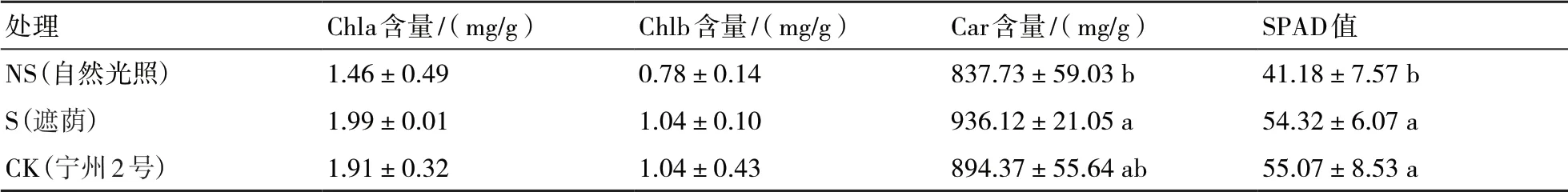
表2 试验茶树第4叶色素含量变化情况
2.2 遮荫对黄金菊叶绿体结构的影响
生长环境尤其是光照强度对新梢芽叶的颜色起到决定性的作用,当对自然生长的茶树黄金菊进行遮荫处理后,其新梢伸展发育成熟,叶片黄化程度逐渐减轻,绿色逐渐加重。取黄金菊成熟叶片(不同叶位)去叶脉的植物组织,经制样后置于TecnaiG220 TWIN透射电镜下观察叶片组织的叶绿体结构,结果如图3、图4所示。对照品种宁州2号、遮荫处理的黄金菊的第2叶、第4叶有典型细胞、线粒体和叶绿体结构,近椭圆形或者圆形的叶绿体贴细胞壁生长于细胞内,叶绿体内具有类囊体、嗜锇颗粒以及紧密堆叠而成的基粒;而自然光照条件下黄金菊的第2叶、第4叶中叶绿体具有类囊体、嗜锇颗粒和基粒,但是紧密堆叠的基粒片数量较少,叶绿体和细胞有膨胀趋势而且体积较大,叶绿体内充满了气泡,细胞内可见叶绿体数量少,类囊体紧密度不及遮荫处理的黄金菊和对照品种宁州2号的。进一步观察发现,遮荫处理的第2叶、第4叶叶绿体数量、片层堆叠明显多于自然光照条件,至第4叶时基本接近CK,甚至叶绿体的数量要多于对照品种宁州2号的。这也说明,黄金菊黄化可能主要与叶绿体数量、叶绿体的片层结构变化有关。
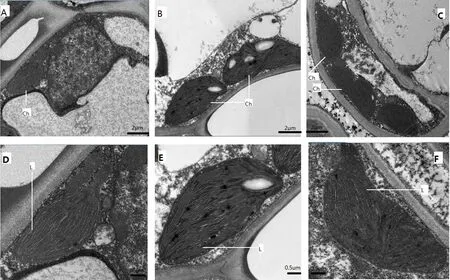
图3 自然光照和遮荫处理茶树第2叶电镜图
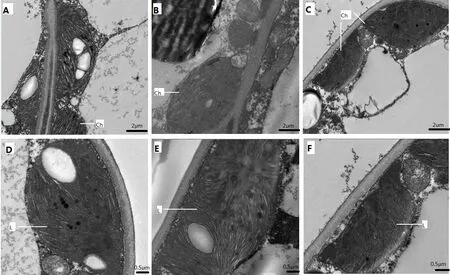
图4 自然光照和遮荫处理茶树第4叶电镜图
3 讨论与结论
一般情况下,黄化或者白化茶树品种的氨基酸含量高于普通绿茶,而酚类物质含量较低[13],这也是使得黄化或者白化茶树滋味更加鲜爽,口感为更多消费者所接受的原因之一。Wei等[14]通过采集不同地区的绿茶春季样品来研究气候条件和叶绿素相对含量对茶叶品质的影响,推测适度低温、较低的叶绿素含量会形成较好的绿茶品质,说明培育低叶绿素品种可以作为改良茶叶品质的一条途径,但叶绿素相对含量过低也会对茶树生长造成影响,完全无法合成叶绿素的茶树在苗期就可能死亡[15]。过去对白化茶树品种白叶1号、白鸡冠研究较多[16],如中国农业科学院茶叶研究所茶树育种团队研究了白叶1号茶树白化—返绿过程中色素蛋白复合体的改变情况[17],结果表明,白叶1号白化期间叶片中保留在捕光叶绿素蛋白复合体中的色素很少。显微镜扫描白叶1号成熟叶片发现,白化期内膜系统逐渐解体[18]。白叶1号全白时期,所有叶肉细胞中叶绿体结构完全解体[19]。吴金全[20]研究了白鸡冠白化前和返绿后的变化情况,结果表明其叶绿体结构和正常品种的差异不明显;由于短期低温环境(20 ℃左右)的影响,温度敏感型茶树叶绿体片层结构受到破坏,叶绿体正常代谢发育、生物合成受阻,进而产生白化植株。
在本试验中,作者以宁州2号为对照,采用了透射电镜技术比较了黄金菊遮荫前后第2叶、第4叶色素含量、亚细胞结构、叶绿体变化的差异,结果发现:遮荫后黄金菊色素含量增加迅速,叶绿体数量增加、叶绿体结构完整,没有出现叶绿体解体及大量囊泡,但叶绿体中基粒片层堆叠明显少于宁州2号。通过比较说明,黄金菊茶树黄化与叶绿体中基粒片层合成减弱有关,但叶绿体损失不及白叶1号、白鸡冠严重[19-20]。这解释了黄金菊黄化而不白化,同时氨基酸含量不够高、酚类物质不低的原因[13,16]。本研究发现黄金菊黄化可能与光胁迫情况下叶绿体数量的减少、叶绿体基粒片层堆叠不紧密或减少有关系。
本文以黄金菊为研究材料,选择宁州2号为对照品种。通过比较黄金菊茶树遮荫前后表型性状、色素含量、SPAD值以及叶绿体超微结构的变化,可以判断黄金菊属于光敏型黄化茶树资源,正常生长的黄金菊茶树由于受到光胁迫以致细胞中的叶绿体数量少、发育不完全,甚至叶绿体基粒片层堆叠不紧密等原因,是导致该茶树黄化的直接原因。相关现象的产生对黄金菊物质代谢、品质性状是否有调控作用,其机制是什么还需进一步分析。本研究为后续江西省深入研究黄色或白色茶树产生,研究该变化的机理打下了坚实的基础。
猜你喜欢
课程教育研究·学法教法研究(2019年18期)2019-10-08
少儿科技(2019年9期)2019-09-10
安徽农学通报(2017年18期)2017-10-18
世界知识(2017年12期)2017-08-01
现代农业科技(2017年11期)2017-07-14
江苏农业科学(2016年2期)2016-04-11
热带农业科学(2014年6期)2014-08-07
科学大众(中学)(2009年6期)2009-07-24
中学生物学(2008年6期)2008-08-29
