
克氏针茅物候对气候变暖和水分变化的响应及其光合生理生态机制
2022-11-12 03:29顾文杰周广胜吕晓敏周梦子汲玉河周怀林
生态学报 2022年20期
顾文杰,周广胜,2,3,4,*,吕晓敏,2,4,周梦子,2,4,周 莉,2,4,汲玉河,2,4,周怀林,4
1 中国气象科学研究院, 北京 100081 2 郑州大学地球系统科学与技术研究院生态气象联合实验室, 郑州 450001 3 南京信息工程大学气象灾害预报预警与评估协同创新中心, 南京 210044 4 河北固城农业气象国家野外科学观测研究站, 保定 072656
植物物候是植物长期适应温度、降水等环境条件季节性变化所形成的彼此相适应的生长发育节律[1],它不仅是植物生长发育状况的直观反映[2],也是指示气候变化的重要指标[3—4],已经广泛应用于指导农事活动及防灾减灾[5]。同时,植物物候作为影响生态系统结构与功能的重要参数[6—7],在调节陆地生物化学循环中发挥着重要作用[8],是陆面过程模型及植物生产力模型的重要参数[9]。因此,研究植物物候变化规律及其调控机制对物候的准确预测及科学应对气候变化均具有重要意义。
已有研究表明,全球植物春季物候整体呈提前趋势,而秋季物候呈推迟趋势[10—11],且这种现象在北半球高纬度地区更为显著[12]。然而,研究发现内蒙古草原植物物候对气候变化的响应存在显著的空间差异性,南部地区植物物候呈现出返青期提前,枯黄期推迟的趋势;而中北部地区则呈现出相反的变化趋势[13]。区域气候的差异及植物-气候相互作用的复杂性导致植物物候研究仍存在很大的差异性与不确定性[14],亟需开展植物物候对气候变化响应的生理生态机制研究。温度[15]、降水[16]、光照[17]、空气湿度[18]、CO2浓度[19]等环境因子是影响植物物候的重要因素,尤其是温度和水分。温度被认为是影响植物物候最重要的环境因子[20],植物只有在一定温度环境中才能生长发育,且需要一定的积温才能完成其生活周期[21]。水分不足限制植物对光、热条件的利用效率[16],被认为是调节干旱半干旱地区植被活动的关键因子[22]。并且,温度与降水之间的相互作用通过改变环境因子的可利用性[23]或植物对环境因子的敏感性[24]显著影响植物物候变化。然而,现有研究大多关注物候变化与环境因子的关系,关于植物物候变化的生理生态机制仍然不清楚[25]。近来研究表明,物候变化与其净光合速率、蒸腾速率、气孔导度等光合生理生态因子密切相关[26],但仍缺乏植物物候对水热环境变化响应的光合生理生态机制研究。
中国温带草原是世界第三大草原区[27],克氏针茅草原是该地区特有的草原群系,是典型草原的代表类型之一,在畜牧业生产中占有重要的地位[28]。并且,克氏针茅草原生态系统脆弱,植物物候已经受到气候变化的显著影响[29]。因此,本研究试图以克氏针茅为研究对象,利用增温与控水相结合的原位控制试验资料,分析研究克氏针茅物候对气候变暖和水分变化的响应及其光合生理生态机制,以增进物候对环境变化响应的理解,并为物候模型发展提供依据。
1 研究区域与研究方法
1.1 研究区概况
试验于2020年在中国气象局内蒙古锡林浩特国家气候观象台(44°08′03″ N、116°19′43″ E,海拔990 m)开展。该地区位于内蒙古典型草原中部,是典型的温带半干旱大陆性气候区,1955—2015年间该地区年均气温为2.5 ℃,年均降水量为283.6 mm[30],冬季寒冷干燥、夏季温暖湿润,太阳辐射较强。试验样地地势平坦开阔,土壤类型主要为淡栗钙土,土壤腐殖质层较薄。优势植物有克氏针茅(Stipakrylovii)和羊草(Leymuschinensis),重要伴生种包括细叶葱(Alliumtenuissimum)、糙隐子草(Cleistogenessquarrosa)、冷蒿(Artemisiafrigida)、矮葱(Alliumamsopodium)、木地肤(Kochiaprostrata)、黄蒿(Artemisiascoparia)、阿尔泰狗娃花(Heteropappusaltaicus)等。
1.2 试验材料与设计
试验以克氏针茅草原作为试验对象,采用红外线辐射增温与控水相结合的原位控制实验方法。该地区气温呈显著上升趋势,但降水年际间波动大,总体趋势不明显[31],未来暖湿化与暖干化现象均有可能出现。因此,以该地区的自然温度和降水为对照,设置5个温度和降水处理:对照(T0W0)、轻度干热(T+1.5W-50%)、重度干热(T+2W-50%)、轻度湿热(T+1.5W+50%)、重度湿热(T+2W+50%) (表1)。每个处理均设置4个重复小区,共20个小区。小区面积为4 m2(2 m×2 m),相邻小区间隔2 m。小区上方安装有长3 m、宽3 m的遮雨棚。小区四周用高0.3 m、深1 m的隔水铁板围起,以减小地表和地下水分流动的影响。
采用不同功率的红外线辐射增温装置对小区进行增温处理。选用的红外辐射灯管长为1 m,平行地面悬挂于小区中心,离地高度为2 m。利用不同遮雨量的遮雨棚装置和灌溉进行水分处理。遮雨板材质为透光率95%以上透光PC板,分为带孔遮雨板和不带孔遮雨板两种类型。通过改变遮雨棚带孔遮雨板和不带孔遮雨板的比例调控遮雨棚的遮雨量。每次降水时还需收集减水处理小区(T+1.5W-50%、T+2W-50%)遮雨棚截留的降水,并将其均匀灌溉到增水处理小区(T+1.5W+50%、T+2W+50%) (表1)。
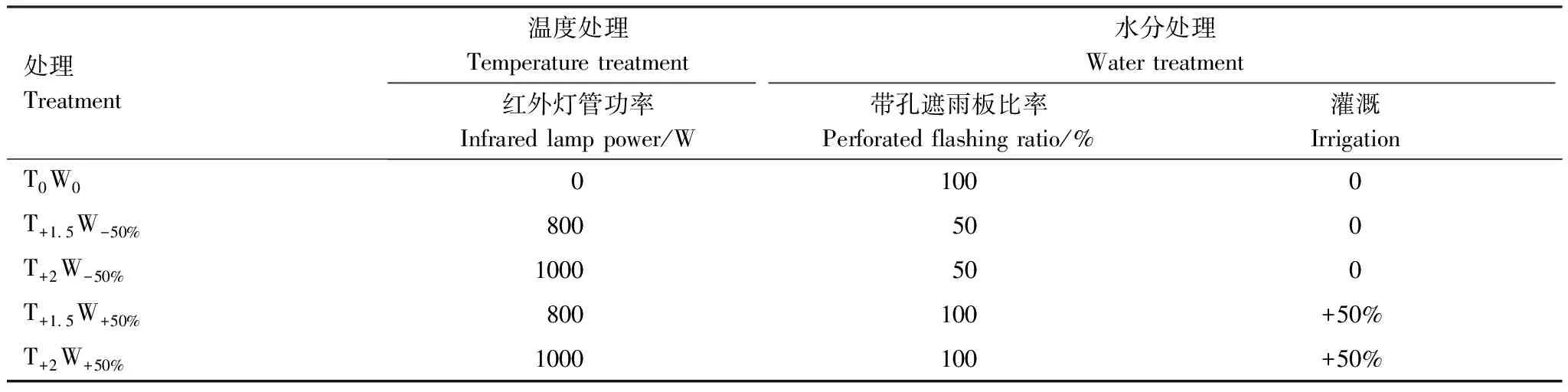
表1 不同小区的水热处理方案
1.3 测定项目与方法
1.3.1土壤温湿度
利用ECH2O土壤温湿度检测系统(METER, Pullman, WA, USA)测定各试验小区的土壤温度(Tsoil, ℃)和湿度(RHsoil,%)。该系统观测频率为30 min/次,利用热敏电阻测定各小区0—10 cm的土壤温度,并通过电容/频域技术测量0—30 cm每10 cm一层的土壤分层体积含水量。
1.3.2物候观测
依据《农业气象观测规范》[32]对克氏针茅物候进行人工观测。每日上午和下午分别对试验样地小区进行巡视,记录各小区克氏针茅到达各关键物候期的日期,并将日期记录转换为同年1月1日开始计算的日序。当观测小区50%克氏针茅的叶片恢复弹性,由黄转青,则认为达到返青期;当小区50%克氏针茅的针茅从叶鞘顶端或侧端露出,记录到达抽穗期;当小区50%克氏针茅的地上器官约有三分之二枯萎变色,记录为枯黄期。
1.3.3叶片光合生理生态参数
为研究气候变暖和水分变化情景下克氏针茅光合生理生态特征的变化,并进一步分析植物物候响应气候变化的光合生理生态机制,利用Li- 6400便携式光合作用系统(LI-COR, Lincoln, NE, USA)测定克氏针茅各关键物候期的光合生理生态参数,测定内容包括叶片净光合速率(Pn, μmol m-2s-1)、气孔导度(Gs, mol m-2s-1)和蒸腾速率(Tr, mmol m-2s-1)等。当克氏针茅到达返青期、抽穗期和枯黄期时,于晴朗无风天气的9:30—11:30进行观测。测定时,每个小区选取1株具有代表性并到达该物候期的克氏针茅,将叶片平展且不重叠地铺满叶室,测定部位为叶片中部,依据仪器使用手册将空气流速设定为300 mmol/s;叶室CO2浓度控制为大气CO2浓度,即400 μmol/mol。光照强度的设定因物候期而不同,通过测定克氏针茅叶片的光响应曲线确定,返青期为1200 μmol m-2s-1,抽穗期为1500 μmol m-2s-1,枯黄期为2000 μmol m-2s-1。叶片水分利用效率(WUE, mmol/mol)计算如下[33]:
WUE=Pn/Tr
(1)
1.4 数据处理
利用Microsoft Excel 2019和SPSS 21.0进行数据整理、汇总与分析。采用配对样本t检验方法,分析不同水热处理下小区土壤温湿度的差异;利用双因素方差分析方法,分析增温和水分对克氏针茅物候的影响;温度、水分和物候期对克氏针茅光合生理生态特征的影响则采用三因素方差分析方法,并用Duncan多重比较方法分析不同处理间物候及光合生理生态特征差异。为阐明光合生理生态参数和土壤温湿度对克氏针茅物候的直接或间接作用方式及影响,利用SPSS AMOS 21.0软件构建通径分析模型进行分析。
2 结果与分析
2.1 试验小区土壤温湿度变化特征
各试验小区土壤温度和湿度在克氏针茅生长季内的波动趋势较为一致,但是土壤温度和湿度在不同增温和控水处理之间存在显著差异(图1)。与对照相比,增温处理小区的土壤温度显著升高;增水处理(T+1.5W+50%、T+2W+50%)小区的土壤湿度显著高于对照,减水处理(T+1.5W-50%、T+2W-50%)小区的土壤湿度显著低于对照。并且,增温处理和水分处理间存在相互影响,在相同温度处理下增水处理小区的土壤温度显著低于减水处理小区;在相同水分处理下,增温2 ℃处理小区的土壤湿度显著低于增温1.5 ℃处理小区。
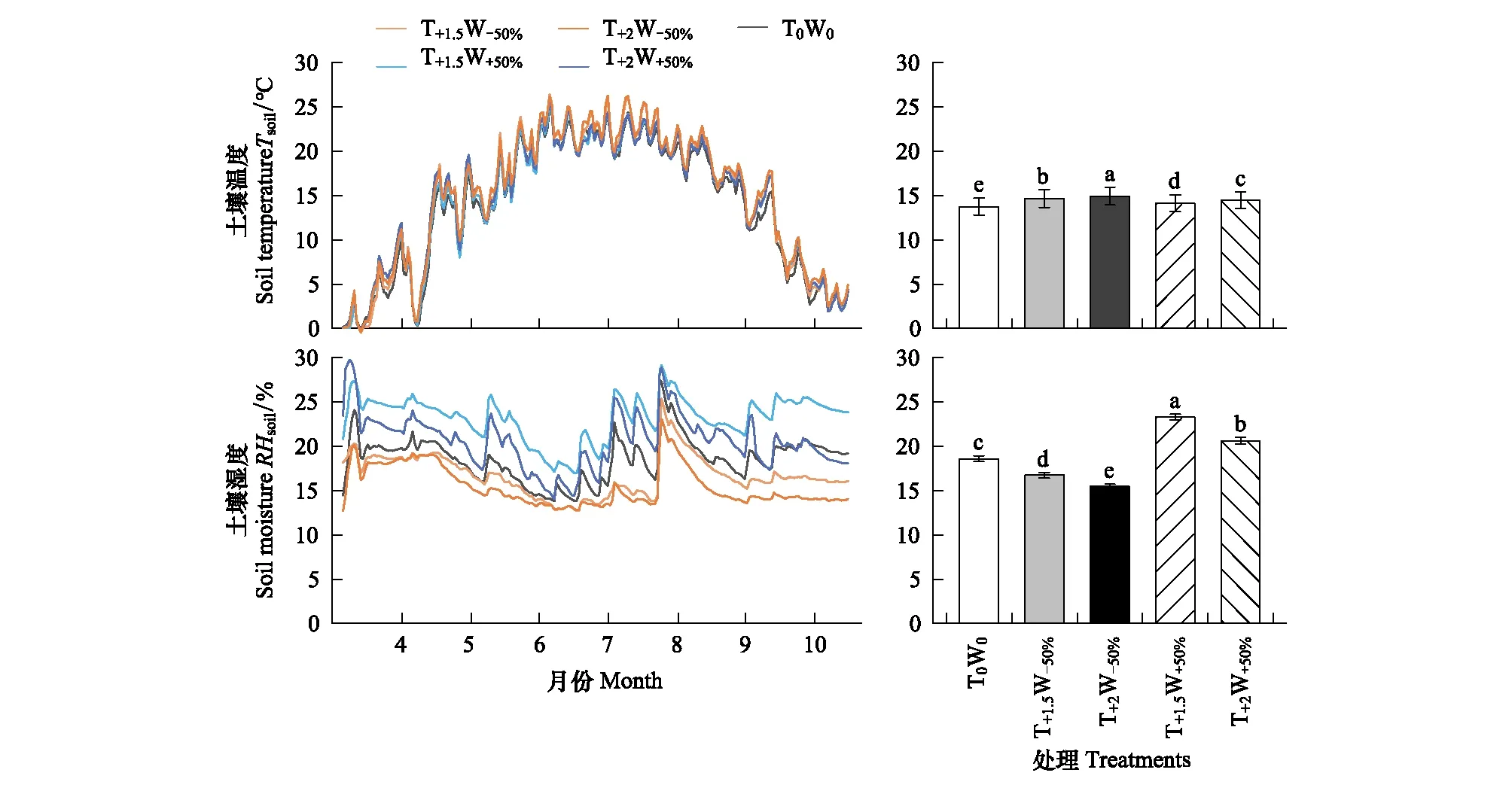
图1 不同增温和控水处理下土壤温度和湿度的动态变化及生长季平均土壤温度和湿度的变化Fig.1 Changes of soil temperature and moisture & average soil temperature and moisture under different warming and precipitation treatments不同小写字母表明不同处理间差异显著(P < 0.05)
2.2 克氏针茅物候期变化
不同增温和水分处理下,克氏针茅到达各物候期的时间存在显著差异(表2)。与对照相比,T+1.5W-50%、T+2W-50%、T+1.5W+50%、T+2W+50%处理的返青期分别提前2 d、3.7 d、2 d和3.5 d,抽穗期分别提前3.8 d、4.5 d、10.5 d和12.5 d,枯黄期分别推迟6.3 d、7.7 d、6 d和7d,导致生长季分别延长8.3 d、11.4 d、9 d和10.5 d。双因素方差分析表明,温度是影响克氏针茅返青期物候的重要因素,水分主要影响克氏针茅的抽穗期,温度和水分协同作用对各物候期的影响不显著(表3)。Duncan多重比较表明,不同水分处理间克氏针茅的抽穗期差异显著,增温使枯黄期显著推迟(表2)。
2.3 克氏针茅光合生理生态特征变化
不同水热处理下,克氏针茅各物候期的光合生理生态特征差异明显(图2)。在克氏针茅的返青期,增温增水处理(T+1.5W+50%、T+2W+50%)的净光合速率、气孔导度和蒸腾速率均显著高于对照(P< 0.05),水分利用效率低于对照,但未达到显著水平;增温减水处理(T+1.5W-50%、T+2W-50%)的净光合速率略高于对照,但蒸腾速率低于对照。减水处理使克氏针茅的水分利用效率提高,且在重度干热处理(T+2W-50%)更为显著(P<0.05)。
与返青期相比,不同水热处理下克氏针茅抽穗期的净光合速率、气孔导度和蒸腾速率均明显下降。增温增水处理(T+1.5W+50%、T+2W+50%)的净光合速率、气孔导度和蒸腾速率均高于对照和增温减水处理(T+1.5W-50%、T+2W-50%),且在增温和水分协同作用下T+1.5W+50%处理的净光合速率、气孔导度和蒸腾速率显著高于T+2W+50%处理(P<0.05)。水分利用效率对增温增水处理的响应不显著,但增温减水处理使克氏针茅的水分利用效率显著升高(P< 0.05)。在枯黄期,克氏针茅的光合能力进一步降低,不同水热处理间的克氏针茅光合生理生态特征差异减小。增温处理下,克氏针茅的净光合速率及水分利用效率显著高于对照(P< 0.05),表明枯黄期的推迟与克氏针茅的光合能力提高密切相关。
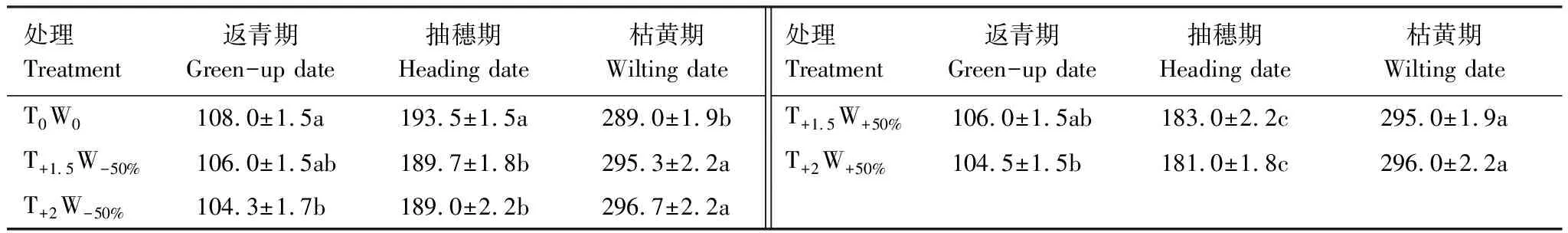
表2 不同增温和控水处理下克氏针茅物候的变化
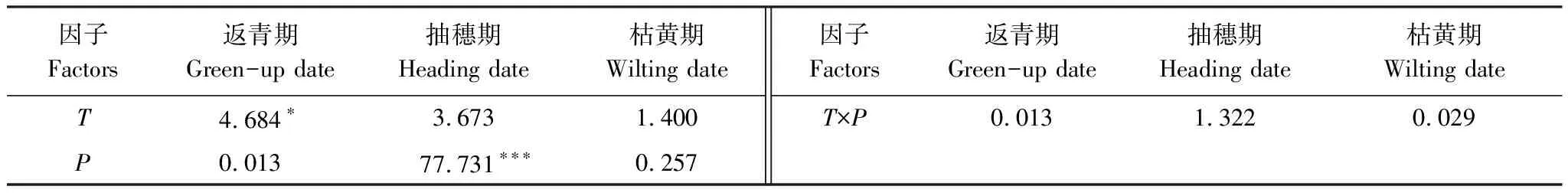
表3 不同增温和控水处理下克氏针茅物候的双因素方差分析 (F值)
三因素方差分析表明,不同物候期的温度和水分协同作用使得克氏针茅的光合生理生态特征变化显著(表4)。不同物候期的克氏针茅的净光合速率、蒸腾速率、气孔导度和水分利用效率差异显著,水分变化显著影响克氏针茅的蒸腾速率、气孔导度、净光合速率和水分利用效率,但温度更多与物候协同作用影响克氏针茅的光合生理生态特征。
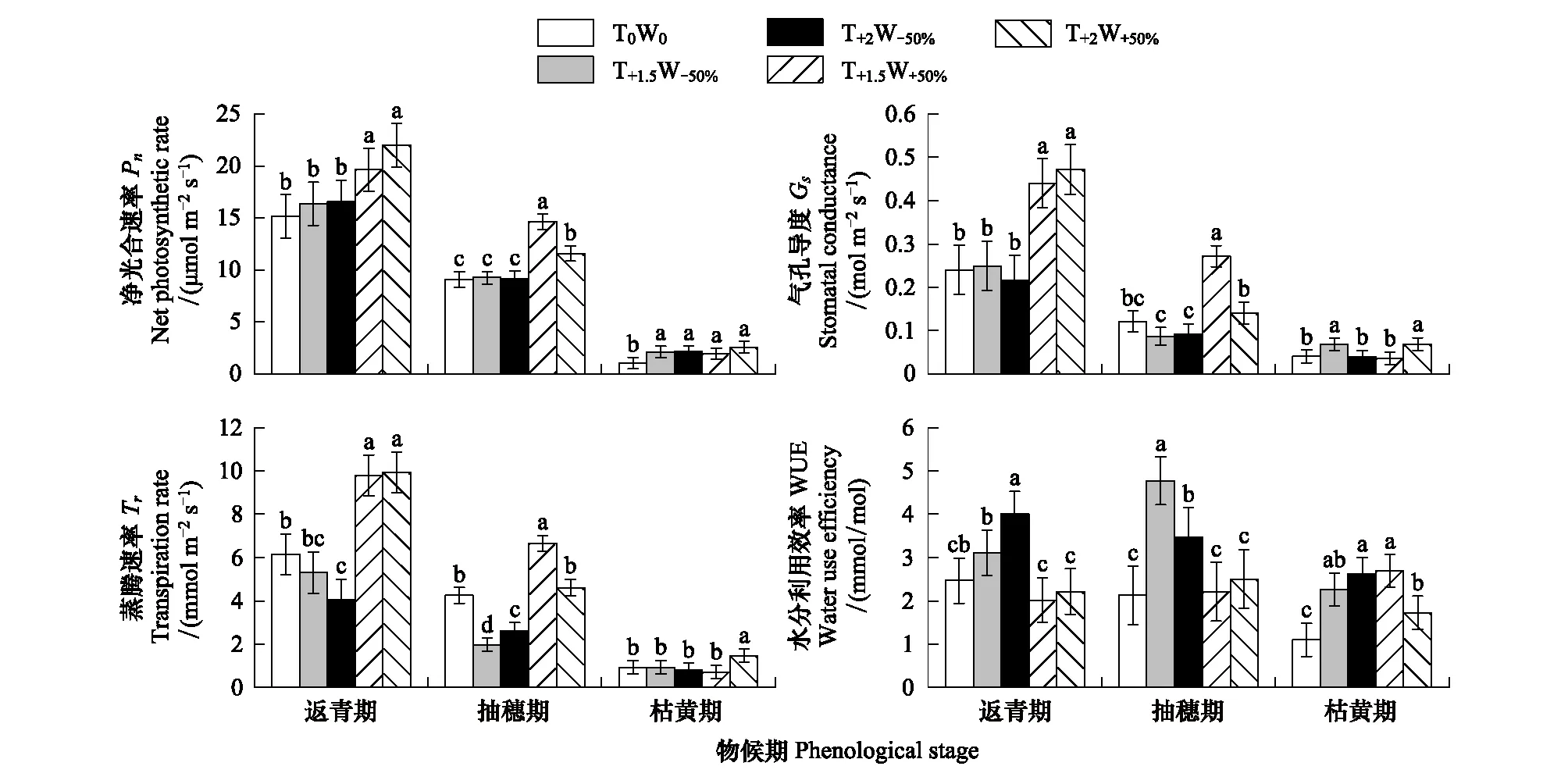
图2 不同处理下克氏针茅主要物候期的净光合速率、气孔导度、蒸腾速率和水分利用效率的变化Fig.2 Changes in net photosynthetic rate, stomatal conductance, transpiration rate and water use efficiency of main phenological stages of Stipa krylovii under different treatmentsGS:返青期 Green-up stage;HS:抽穗期 Heading stage;WS:枯黄期 Wilting stage;不同小写字母表明不同处理间差异显著(P<0.05)

表4 不同处理下克氏针茅光合生理生态特征的三因素方差分析结果 (F值)
2.4 克氏针茅物候变化的光合生理生态机制
通径分析(图3)表明,光合生理生态参数、土壤温度和土壤湿度对克氏针茅的返青期、抽穗期和枯黄期的解释率分别为95%、58%和62%,且对克氏针茅物候期直接作用最大的因子均为净光合速率,说明净光合速率是导致克氏针茅物候变化的主要光合生理生态因子。返青期和抽穗期的净光合速率对克氏针茅物候的直接效应分别为-0.961和-0.864,表明净光合速率的增加促进返青期和抽穗期的提前;枯黄期的净光合速率对克氏针茅物候的直接效应为0.790,表明净光合速率的增加导致枯黄期推迟。返青期的土壤温度对物候的间接效应主要通过水分利用效率影响蒸腾作用及气孔导度,进而影响净光合速率,温度升高促使克氏针茅的返青期提前(-0.191)。抽穗期的土壤温度对物候的间接效应(-0.181),主要通过水分利用效率影响蒸腾作用,而土壤湿度对物候的间接效应(-0.750),主要是通过气孔导度影响净光合速率。枯黄期的土壤温度通过影响净光合速率间接影响植物物候(0.256),土壤温度对枯黄期的总效应为正值,表明温度升高导致枯黄期物候推迟(表5)。
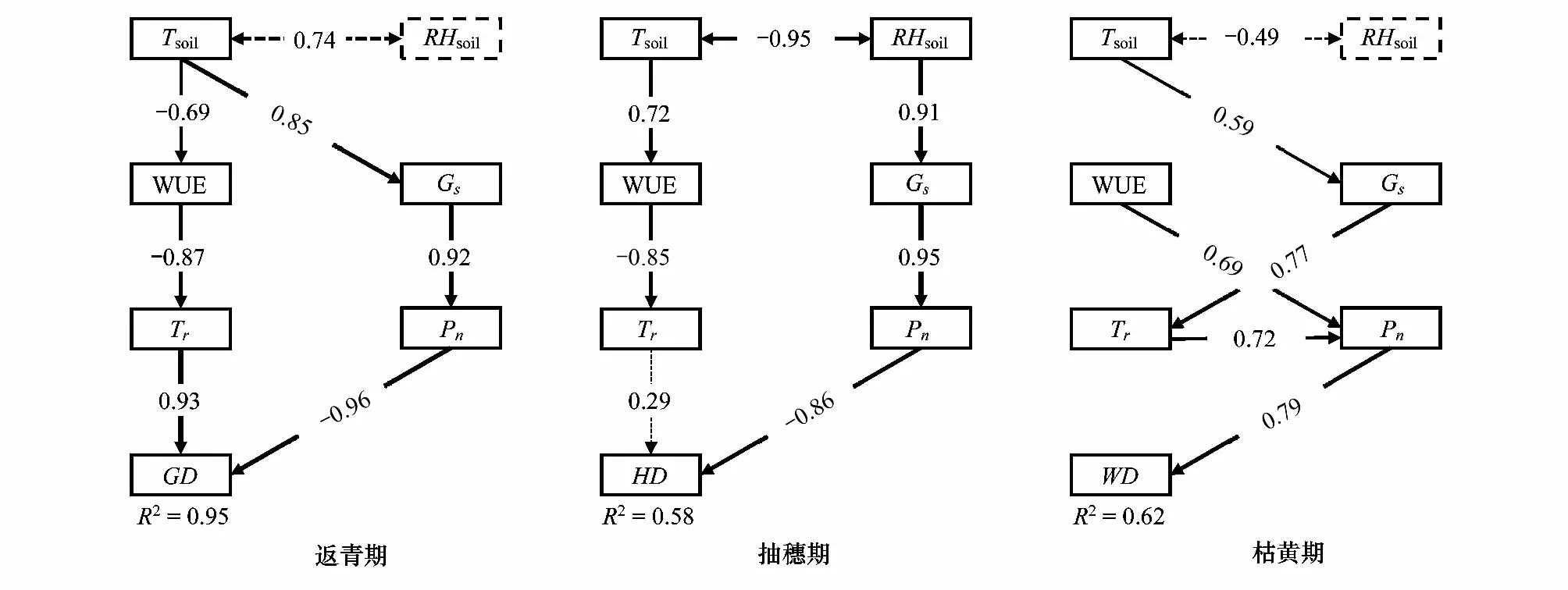
图3 克氏针茅物候和光合生理生态因子的通径分析Fig.3 Path analysis of phenology and photosynthetic ecophysiological characteristics of Stipa kryloviiTsoil:土壤温度 Soil temperature;RHsoil:土壤湿度 Soil moisture;WUE:水分利用效率 Water use efficiency;Gs:气孔导度 Stomatal conductance;Tr:蒸腾速率 Transpiration rate;Pn:净光合速率 Net photosynthetic rate;GD:返青期 Green-up date;HD:抽穗期 Heading date;WD:枯黄期 Wilting date;----:P < 0.05;:P > 0.05
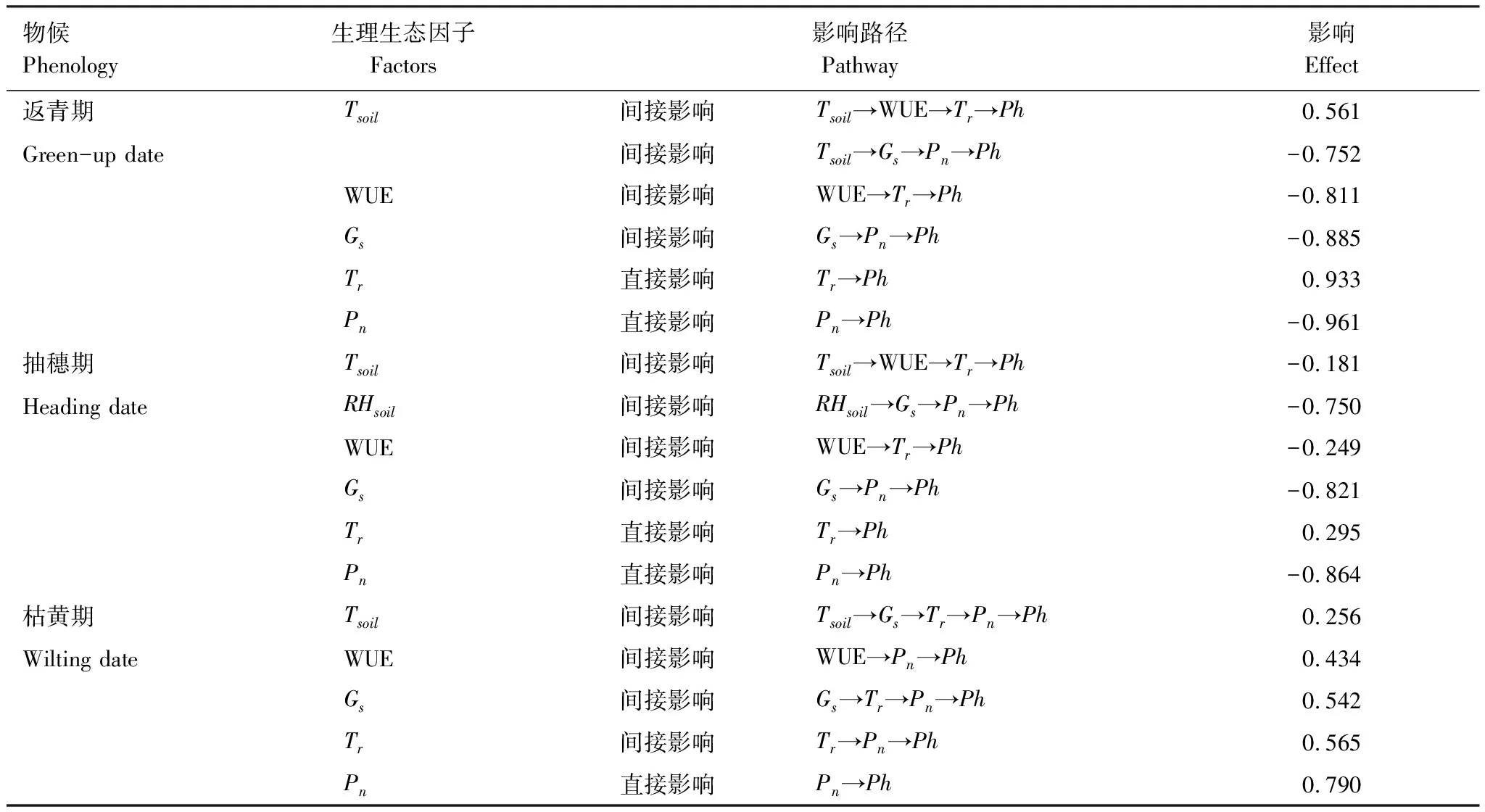
表5 克氏针茅物候和光合生理生态因子的通径分析
3 讨论与结论
以气候变暖为主要特征的全球气候变化已经成为事实[34],已经影响土壤理化性质和植物生理生态特征,进而影响土壤-植物系统[35]。本研究基于红外线辐射增温与控水相结合的原位模拟试验发现,土壤温度和湿度之间存在相互影响。已有研究表明,气候变暖会导致土壤温度升高,对土壤湿度的影响则表现出显著的季节差异[36]。夏季增温导致蒸散加强,土壤湿度降低[37];冬季增温则增大土壤与大气间的温差,使水汽更易液化渗入土壤,导致土壤湿度升高[35]。
气候变化已经导致草原植物物候发生显著改变。1965—2013年,青藏高原地区牧草返青期、抽穗期和开花期平均每10年分别提前7.4 d、6.2 d和6.4 d,而枯黄期则呈现推迟趋势(2.1 d/10 a),温度升高被认为是导致物候变化的主要原因[38]。但受气候变暖影响,内蒙古通辽地区气候暖干化,水分胁迫成为导致该地区羊草、冰草(Agropyroncristatum)和委陵菜(Potentillachinensis)返青期推迟,枯黄提前的重要原因[39]。生存分析模型表明,气温升高2 ℃将导致草原植物返青期提前1—4 d[40];开顶式箱(OTC)增温模拟表明,增温2.2 ℃使钉柱委陵菜(Potentillasaundersiana)的生殖期提前3—5 d,但使高山嵩草(Kobresiapygmaea)的生殖期推迟5—7 d,增温导致的水分胁迫被认为是造成高山嵩草物候期推迟的原因[41]。本研究发现,增温处理下克氏针茅植株的返青期提前2—4 d,抽穗期提前3—13 d,枯黄期推迟6—8 d,物候期变化趋势与已有研究结果一致[42]。研究还发现,克氏针茅在抽穗期对水分变化敏感,增温增水处理使抽穗期提前,较增温减水处理提前6—9 d。
环境因子是影响植物生长发育的重要因素,也是植物生理生态变化的诱因。植物光合作用本质上是一系列酶促反应,温度升高显著提高植物光合作用的重要环节RuBP的羧化和再生的最大速率[43]。本研究也发现,增温处理下克氏针茅的净光合速率升高。但有研究表明,植物净光合速率、气孔导度和蒸腾速率对升温的响应存在物种间差异,增温促进了发草(Deschampsiacaespitosa)的光合作用,但抑制了遏蓝菜(Thalaspiarvense)的光合作用[44]。水分是限制植物叶片光合作用的关键因素,降水增加将导致植物光合速率、气孔导度和蒸腾速率升高,而水分胁迫将导致植物气孔导度和蒸腾速率下降,水分利用效率提高[45],本研究也观测到一致的现象。
植物物候是植物对温度、降水等环境条件季节性变化的适应结果,是植物可利用气候资源的综合体现。植物的净光合速率体现了植物利用气候资源将CO2和水转化为有机物的快慢[46]。本研究通过原位控制试验,将环境因子、光合生理生态因子和植物物候联系起来,并分析了克氏针茅物候对气候变暖和水分变化的响应及其光合生理生态机制,得到以下结论:(1)温度升高导致克氏针茅的返青期和抽穗期提前、枯黄期推迟,水热协同作用对克氏针茅物候的影响主要体现在抽穗期。(2)在返青期和抽穗期,增温增水导致克氏针茅的净光合速率、气孔导度和蒸腾速率显著升高,而增温减水则导致水分利用效率显著升高。对于枯黄期,增温使净光合速率和水分利用效率升高,不同水分处理间光合生理生态特征无显著差异。(3)不同物候期,影响克氏针茅物候的最重要直接因子均为净光合速率,净光合速率升高导致克氏针茅返青期、抽穗期提前,枯黄期推迟。(4)克氏针茅物候期与环境因子及其光合生理生态特征密切相关。净光合速率是植物物候变化的决策因子,温度和降水则是植物物候变化的限制因子。
致谢:感谢中国气象局内蒙古锡林浩特国家气候观象台对试验提供的大力支持。
猜你喜欢
实用手外科杂志(2022年2期)2022-08-31
实用手外科杂志(2022年2期)2022-08-31
中国农业气象(2022年8期)2022-08-24
当代水产(2022年6期)2022-06-29
中国典型病例大全(2022年13期)2022-05-10
农业工程学报(2022年4期)2022-04-24
土壤(2022年1期)2022-03-16
农业与技术(2020年12期)2020-07-04
江苏农业科学(2017年21期)2017-12-13
新课程·下旬(2017年7期)2017-08-14
