
基于RAPD分子标记的藜蒿种质资源遗传多样性分析
2022-11-11 03:44:32王宇航魏清莹薛天源何思晓陈伟达董元火曾长立
湖南生态科学学报 2022年4期
王宇航,魏清莹,薛天源,何思晓,陈伟达,董元火,曾长立
(江汉大学 生命科学学院/湖北省汉江流域特色生物资源保护开发与利用工程技术研究中心,湖北 武汉 430056)
藜蒿(Artemisiaselengensis),又名蒌蒿、芦蒿、水蒿,是菊科蒿属的一种多年生草本植物。蒿属(Artemisia Linn)全属约300余种,常见植物有黄花蒿、青蒿、艾蒿和冷蒿等,主要分布于亚洲、欧洲、北美洲的温带和亚热带地区等。其中,我国蒿属植物有180余种,主要分布在东北、华北、西北和西南[1]。藜蒿嫩茎为青绿色或紫红色;嫩叶多为青绿色,老叶为深绿色;具有大量侧根和纤维状须根,有匍匐的地下茎,整株直立生长[2]。藜蒿含有多种药理活性物质如黄酮、绿原酸和多酚等,具有抗氧化[3]、抗肿瘤[4]等作用;同时也富含粗纤维、多糖和维生素等营养物质,作为蔬菜的同时兼具保健功能。藜蒿种质资源分布范围广泛、分化复杂且物种间差异小,形态学等常规方法难以有效鉴定和区分不同种质资源。目前,已知的藜蒿基因组信息较少且不够完整,不便于采用基因或基因组学方法加以鉴别和区分。
采用DNA分子标记进行物种的亲缘关系和遗传多样性分析、遗传图谱构建等工作,与传统形态学标记、细胞学标记相比,DNA分子标记不受季节、环境限制,在揭示物种多样性的变化程度和分布方面具有无可比拟的优势[5]。张淑艳[6]利用ISSR和RAPD分子标记对3个近缘种沙蒿的15个种群进行了遗传多样性和亲缘关系分析,两种标记结果均显示3种蒿大部分遗传变异存在于种群内,3个近缘种种群间遗传分化较小。Pandey等[7]通过形态学表征和RAPD标记进行建树聚类,并开发出与青蒿素含量连锁的标记,为盐渍地种植青蒿变种提供了可能。Younsi等[8]对8个野生种群共80个白蒿个体进行RAPD和ISSR标记分析,结果显示其群体内存在高水平的遗传多样性,对于该药用物种的资源保护具有重要意义。Nganthoi等[9]利用RAPD、ISSR、IRAP三种分子标记评估了4个蒿属分类群之间的遗传关系,观察到了高水平多态性,并通过进化树的分析将4个分类群归为2个组群。Huong Thi Nguyen[10]利用RAPD和ISSR标记对9份艾草资源进行了遗传多样性分析,结果证实艾草种内变异性很大。Elmeer等[11]利用RAPD和SRAP对3个不同海拔的18个白蒿个体的遗传变异进行了研究,系统发育关系显示其可分为2个主要类群,结果表明遗传关系与植物的地理位置分布相关;Oyundelger等[12]利用全基因组测序技术开发了新的物种特异性简单序列重复(SSR)标记,对沿蒙古大气候梯度分布的11个冷蒿种群分析发现,整个蒙古草原的冷蒿种群中存在着大量的基因流。
随机扩增多态性DNA标记(RAPD)是一种基于聚合酶链式反应(PCR)的分子标记技术,操作简单、快速、成本低,被广泛用于物种分类、种质资源亲缘关系鉴定和遗传多样性分析[13]。目前,鲜有应用分子标记技术对藜蒿种质资源进行分类鉴定和遗传多样性分析的研究报道。因此,本研究采用RAPD分子标记技术对国内收集到的102份藜蒿种质资源进行系统分类和遗传多样性分析,以明晰不同品系藜蒿资源的亲缘关系,为藜蒿种质资源的遗传多样性保护和遗传改良提供科学依据。
1 材料与方法
1.1 试验材料
供试的102份藜蒿材料收集于全国6个省份,后种植于湖北省汉江流域特色生物资源保护开发与利用工程技术研究中心汉南基地藜蒿种质资源圃。因试验材料均为未登记品种,所以本文以序号代表(表1)。试验所用材料均为健康生长且无病虫害污染的新鲜植物叶片。
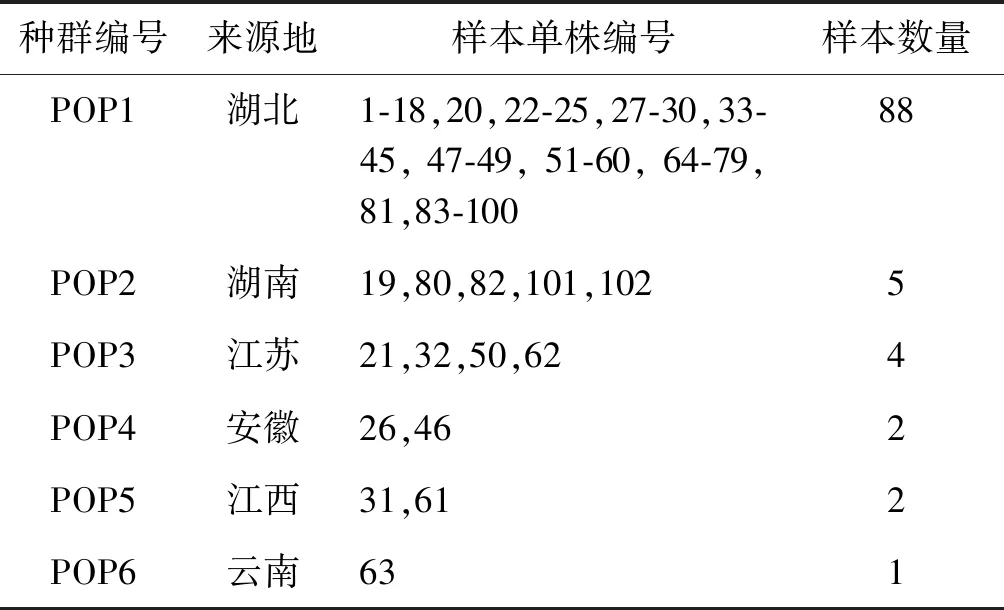
表1 102份藜蒿材料信息Table 1 Information of 102 Artemisia selengensis
1.2 藜蒿基因组DNA提取与检测
剪取各品系幼嫩叶片用液氮研磨成粉末,采用改良的CTAB方法提取基因组DNA。用1%琼脂糖凝胶电泳检测DNA提取质量;使用超微量分光光度计测定各样品DNA浓度,然后用ddH2O稀释至30 ng/μL,-20 ℃保存备用。
1.3 RAPD引物合成及筛选
通过查阅相关文献[14-18],最终选取18条RAPD引物交由北京擎科生物科技有限公司武汉分公司进行合成(表2)。首先,从各种群中分别选取1个代表性样品DNA进行引物初筛,再根据扩增结果进行复筛,对于条带清晰、多态性高、重复性好和扩增效率高的引物,将用于后续实验。
1.4 RAPD扩增体系及程序
反应总体积(20 μL)为:模板DNA 1.5 μL(25 ng/μL);引物0.8 μL(35 ng/μL);2×Es Taq Master Mix(含有Es Taq DNA Polymerase,3 mM MgCl2和400 μM每种dNTP)9 μL;ddH2O补足至20 μL。
扩增程序为:94 ℃预变性5 min;94 ℃变性30 s,34 ℃退火30 s,72 ℃延伸45 s,35个循环;72 ℃延伸5 min,最后4 ℃保存。PCR扩增产物使用1%琼脂糖凝胶进行电泳,电泳结果在凝胶成像系统中观察并拍照。
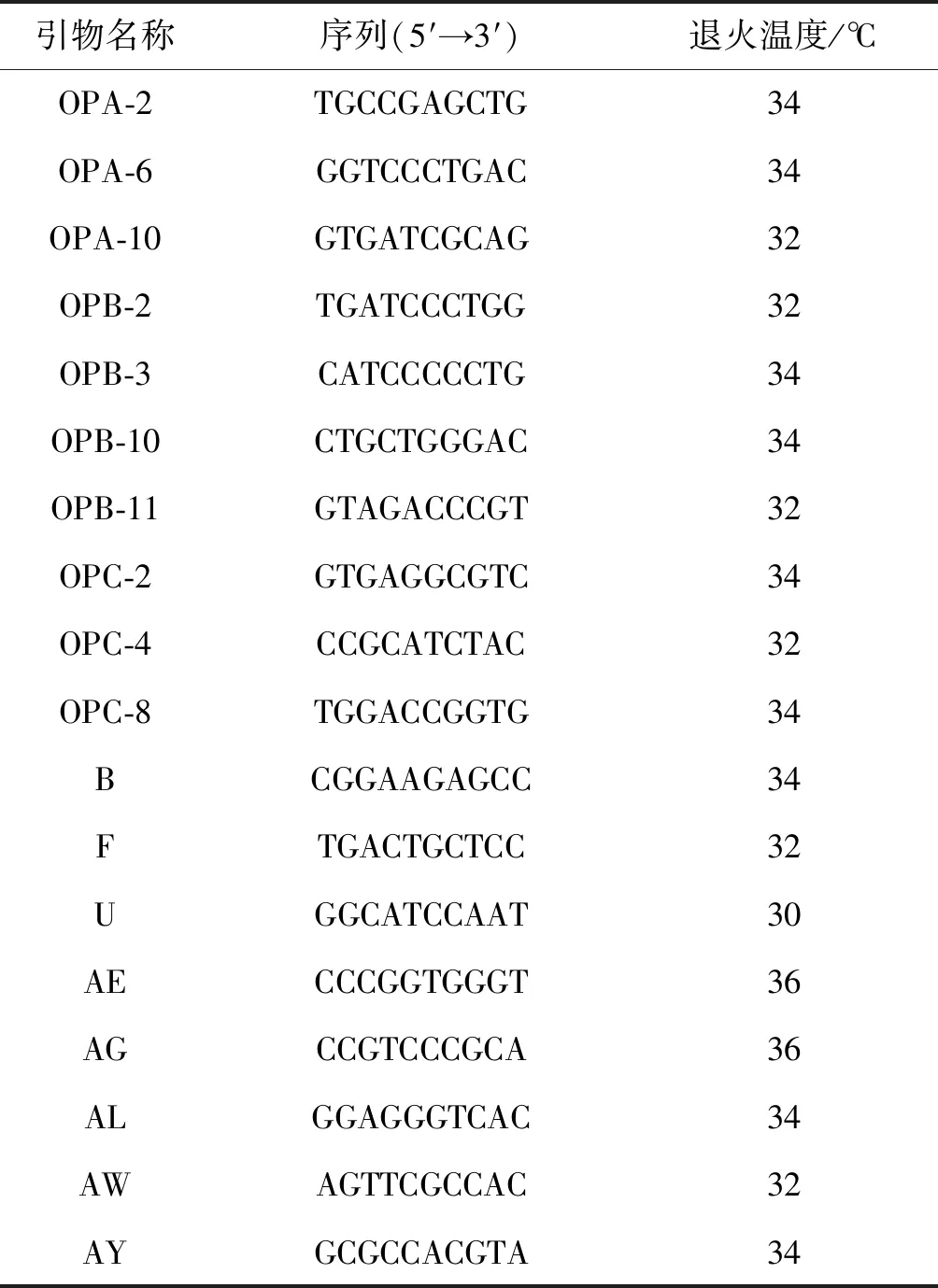
表2 RAPD-PCR引物列表Table 2 List of Primers for RAPD-PCR
1.5 数据处理与分析
选择扩增结果清晰、可重复的条带进行统计,在同一位点出现条带的记为“1”,条带不清晰或无条带则记为“0”,使用Excel表格记录统计结果,最后生成二元的“0”、“1”矩阵,利用R软件进行遗传距离计算和进化树构建,使用POPGENE32软件进行遗传多样性分析,利用STRUCTURE v2.3.4软件进行群体结构分析。
2 结果与分析
2.1 藜蒿样本基因组DNA提取结果
供试材料的DNA使用1%琼脂糖凝胶电泳检测,条带清晰完整(图1),使用超微量分光光度计测定基因组DNA浓度在200~600 ng/μL,OD260/OD280值在1.8~2.0左右,可以用于后续的RAPD-PCR试验分析。
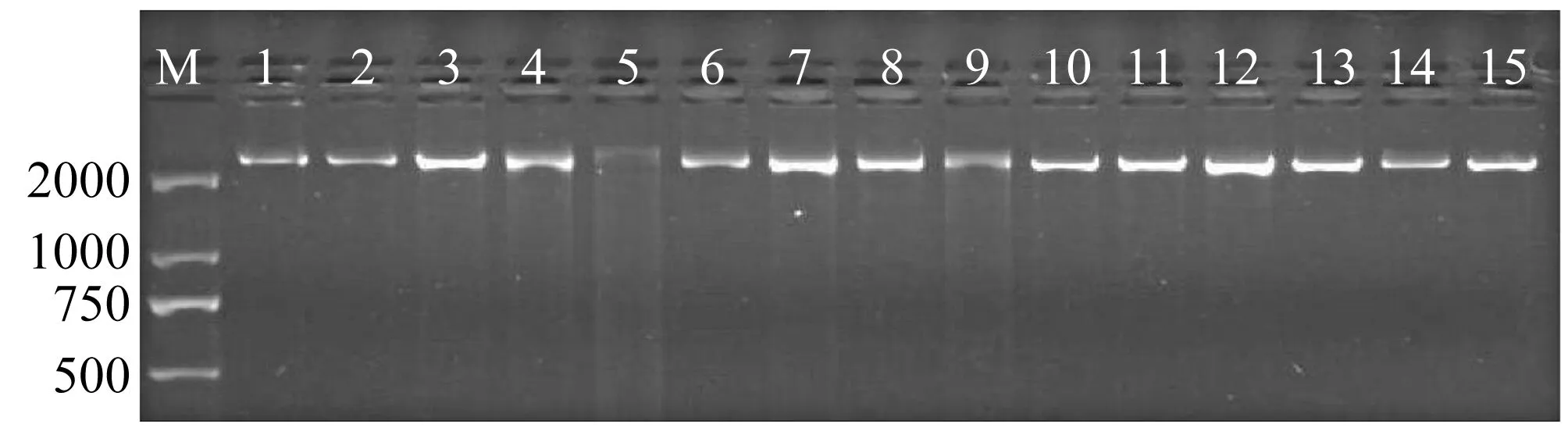
图1 藜蒿DNA检测电泳图Figure 1 Electrophoresis of Artemisia selengensis DNA detection注:1~15表示供试材料1~15号,M为2 000 bp DNA marker
2.2 藜蒿扩增多态性分析
从18条设计的RAPD引物中共筛选出11条扩增条带清晰、多态性高和重复性好的引物(表3),对6个省份的102个藜蒿样本进行RAPD-PCR扩增,引物AG扩增结果如(图2)。结果共获得95个条带,每条引物平均扩增出8.6个条带,扩增条带最多的引物为OPC-8,扩增出13条,引物AL扩增条带最少,仅7条,所有引物扩增的片段大小范围在200~2 000 bp,其中多态性位点93个,多态性位点比率为97.89%,表明本研究所用的RAPD引物具有高的多态性,种群间Nei遗传多样性指数(H)标准差为0.152 3,均值为0.288 3,Shannon遗传多样性指数(I)标准差为0.196 9,均值为0.444 5,表明收集的藜蒿种质资源具有丰富的的遗传多样性。(表4)。
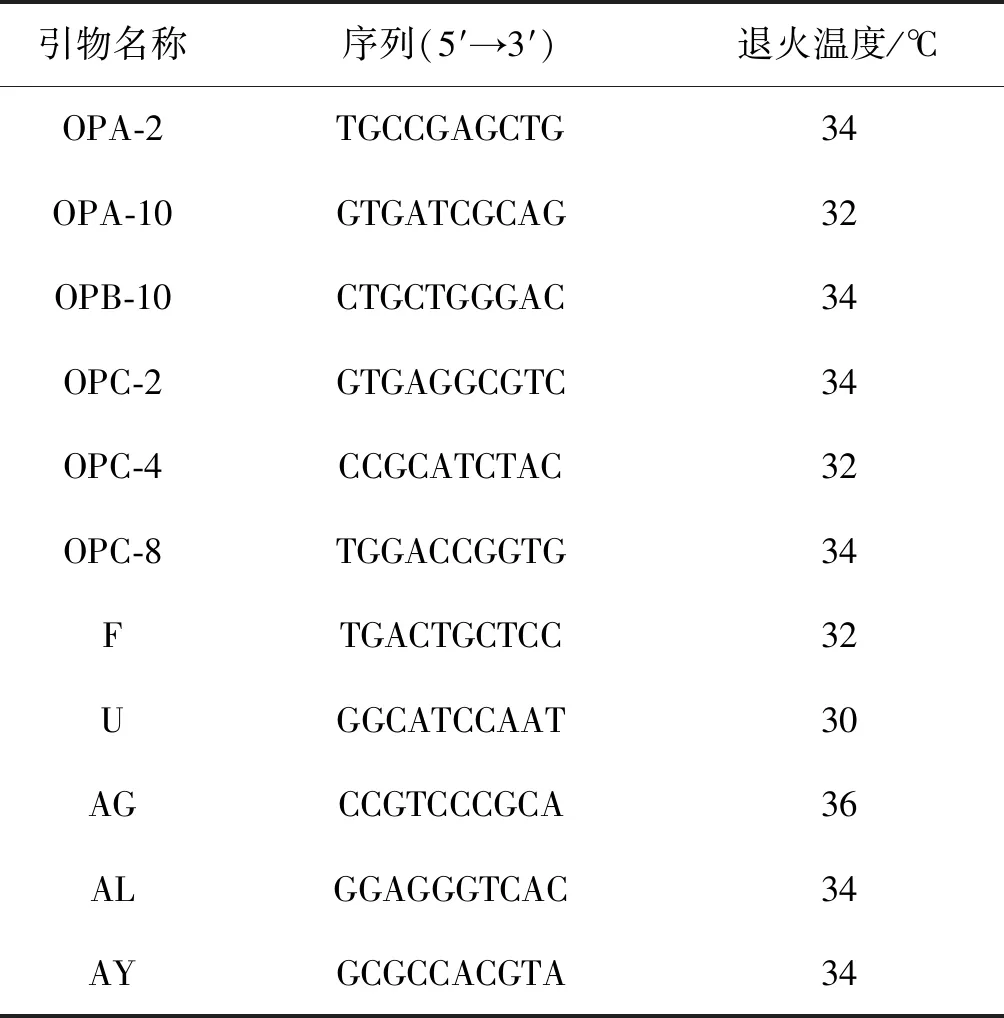
表3 试验所用的RAPD引物Table 3 RAPD primers used in the experiment
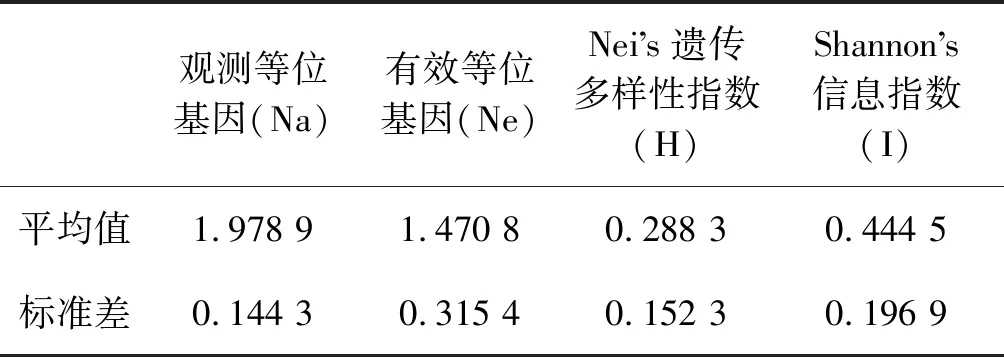
表4 102份藜蒿样品遗传多样性信息Table 4 102 genetic diversity information of Artemisia selengensis samples
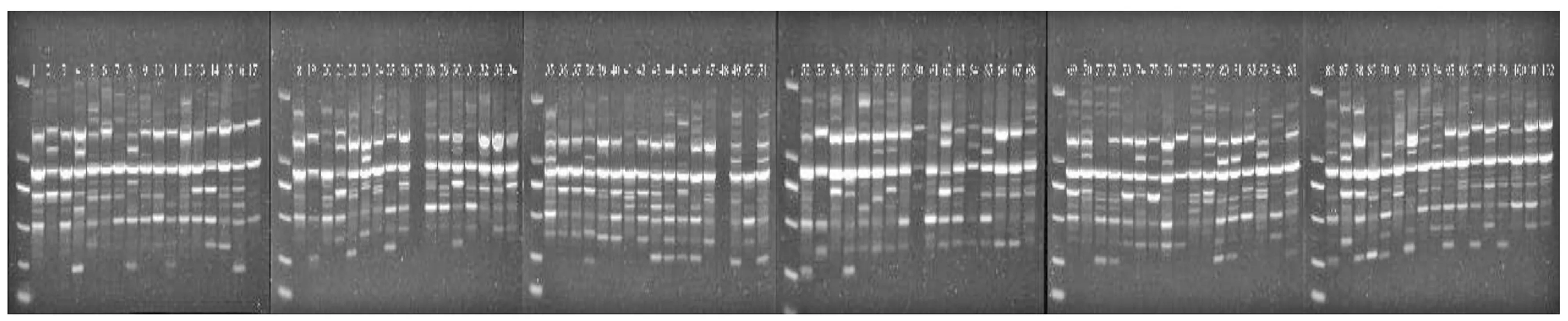
图2 引物AG对102份藜蒿材料的扩增产物电泳图Figure 2 Electrophoresis of the amplification products of primers AG to 102 samples of Artemisia selengensis
2.3 遗传分化分析
对藜蒿6个种群的总基因多样性(Ht)和种群内基因多样性(Hs)分析发现,Ht值为0.249 3,Hs值为0.146 9表明遗传变异主要是由于种群内部变异引起的;藜蒿种群间基因分化系数为41.09%,种群内为58.91%,基因流(Nm)为0.717 0,表明种群之间基因交流程度低(表5)。
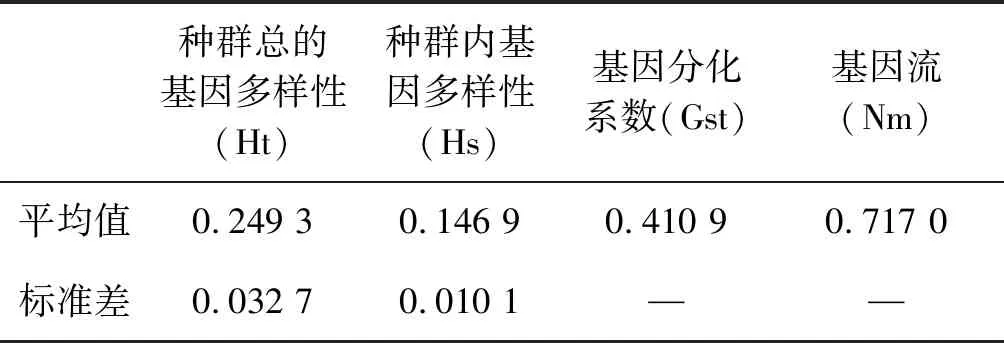
表5 藜蒿遗传分化分析
2.4 藜蒿种群间遗传相似系数和遗传距离分析
利用POPGENE32软件对藜蒿6个种群的遗传距离和遗传相似系数进行计算,结果表明,POP1和POP5、POP2的遗传距离显著小于其它种群,其中POP1和POP5最近,为0.044 0,表明这2个种群之间存在频繁的基因交流,具有相似的遗传信息。POP1与POP2遗传距离为0.061 5,表明POP1与POP5、POP2有着密切的亲缘关系;而POP4和POP6两个种群之间的遗传距离最远,为0.274 9,表明这2个种群之间基因流动较少(表6)。遗传相似系数分析结果和遗传距离分析结果趋于一致,6个种群聚类如(图3),在遗传距离为0.1时,分为两个组,第一组中湖北先和江西聚在一起,然后依次与湖南、江苏和安徽聚为一支,种群之间存在基因交流,第二组云南单独为一支,与其他种群遗传距离较远。
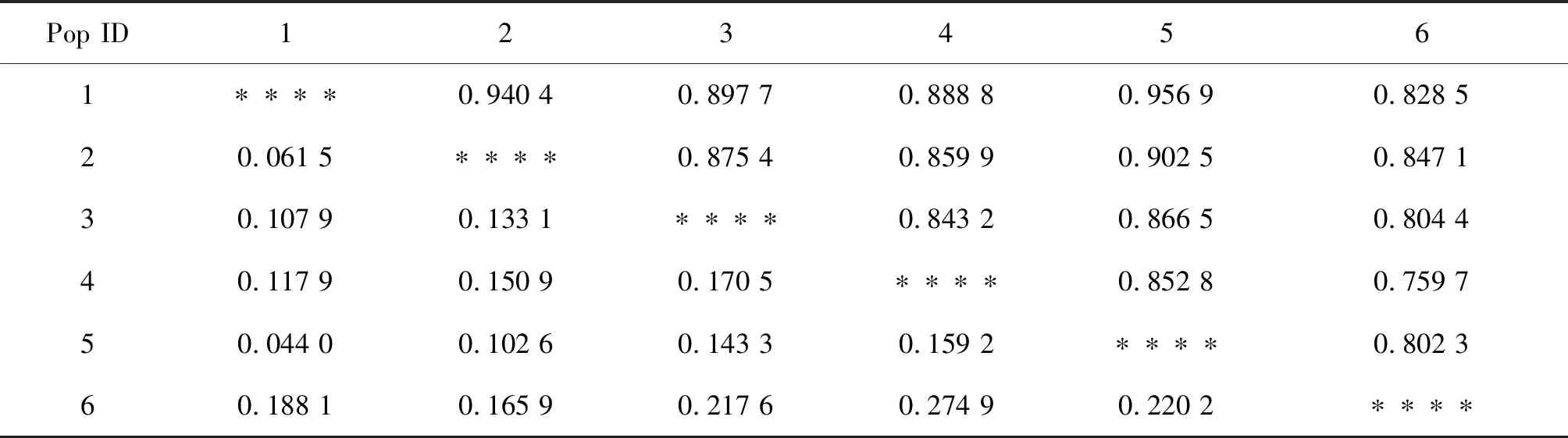
表6 藜蒿6个种群遗传相似系数和遗传距离Table 6 Genetic similarity coefficient and genetic distance of 6 populations of Artemisia selengensis
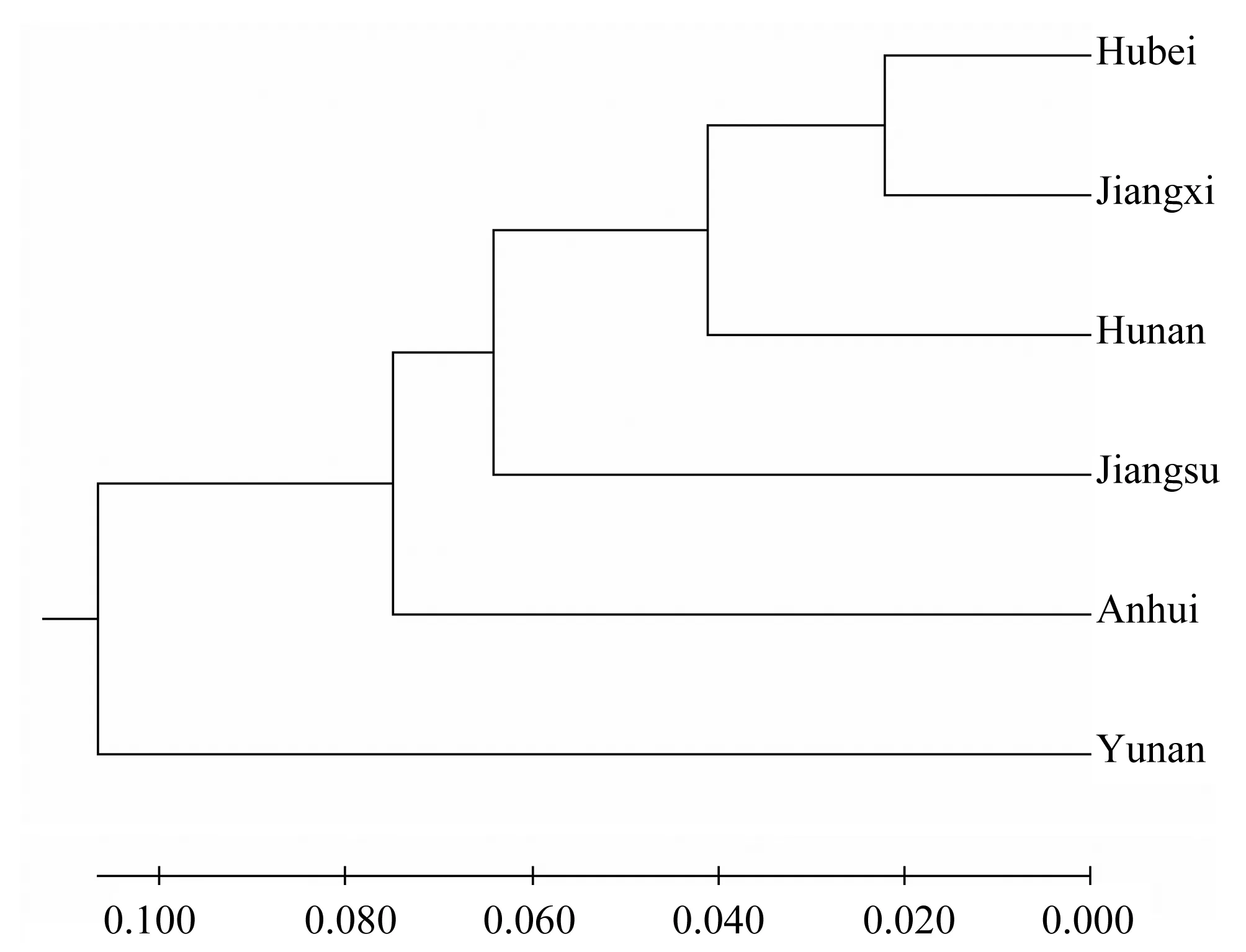
图3 6个藜蒿种群聚类图Figure 3 Chuster diagram of 6 Artemisia selengensis populations
2.5 RAPD分子标记聚类分析
使用R软件将RAPD标记矩阵进行分析并构建遗传进化树,基于遗传距离的UPGMA聚类分析显示,102个藜蒿个体的遗传相似系数在0.61~0.98之间(见图4)。当遗传相似系数为0.62时,102份材料可分为三大组。第1组包含96份藜蒿样本,第2组包含5个藜蒿样本,均来自于湖北,第三组来自于湖南的19号藜蒿样本单独分为一支。当相似系数在0.68时,第一大组又分为五个亚组,第一个亚组数量最多包含90个样本,江苏的2个样本21和62聚为第二亚组,湖北的样本71、样本72聚为一个分支组成第三亚组,样本52单独分支为第四亚组,样本61单独聚为一支为第五亚组。其中第一个亚组中,6个种群的样本并未按照各自种群分别聚为一类,而是随机分布在所在样本的小分支。通过对6个省份102份藜蒿资源的聚类分析,结果表明样本之间遗传距离较小,亲缘关系密切,藜蒿不同群体之间存在不同程度的基因流动。
2.6 群体结构分析
藜蒿多为无性繁殖,但是群体内单株也会产生遗传突变。为进一步揭示藜蒿单株种质的遗传组分,研究利用STRUCTURE2.3.4软件对102份藜蒿材料的RAPD标记位点信息进行群体结构分析。基于Evanno[19]的ΔK方法进行贝叶斯聚类分析的结果显示,ΔK值最高且K=3,根据具有不同遗传结构的每个样本百分比,可以将藜蒿材料分为3个亚群。在K=3时产生RAPD的最高峰值,表明试验材料中有三个可能的信息亚群(图5)。因此,供试的102份藜蒿材料被分为3个大组,红色(第1组)、绿色(第2组)和蓝色(第3组)。
群体结构分析结果中三个大组,第1组包含40个样本,4份来源于湖南,安徽和云南各1份,其余来源于湖北;第2组包含41个样本,2份来源于江西,安徽和江苏各1份,剩余为湖北样本;第3组包含10个样本,1份来自于湖南,其余来自于湖北;另外11个样本遗传结构来源复杂,无法明确归属类群。6个种群102份材料在划分的3个组中交叉分散分布,存在一定的基因流动。模型聚类结果在一定程度上与距离聚类表现出一致性,更直观的展现了各个样本之间的亲缘关系。
3 讨论与结论
分子标记技术由于其操作简单、快速,成本低,是研究种质资源亲缘关系、遗传多样性以及群体结构分析的常用方法[20]。本研究从18条RAPD引物中共筛选出11个条带清晰、多态性高和稳定性好的引物,并用其对102份藜蒿种质资源进行了亲缘关系分析,扩增出的多态性位点百分比为97.89%,徐彪等[21]利用RAPD分子标记研究向日葵自育恢复系遗传多样性,多态性百分比为73.68%,相比之下,多态性低于本试验。
通过遗传分化分析来预估物种的遗传变异在种群内部和种群之间的分布程度,可为之后的种质资源多样性保护和遗传改良提供参考[22]。本研究发现,6个藜蒿种群Gst为41.09%,表明大多数遗传变异发生在种群内部,而石丽敏[23]通过形态学、同工酶和RAPD鉴定收集到的4份不同的芦蒿资源,遗传多样性结果显示芦蒿种内遗传变异不高。基因流(Nm)为0.717 0(<1),也表明种群之间基因流动较少,藜蒿6个种群之间的遗传分化程度较高。
聚类分析是根据供试材料的多个数据指标进行归类,包括距离聚类和模型聚类等,是研究种质资源亲缘关系、遗传多样性和种群结构的重要手段[24]。聚类分析可以直观的反应出物种之间和种群之间的遗传关系和遗传一致度。本研究在RAPD分析基础上对102份藜蒿资源进行UPGMA聚类和基于贝叶斯模型聚类,初步将102份藜蒿资源分为3个大类。由于6个种群的采样不均,种群结构分析结果中安徽和江苏的种群样本聚类结果较为分散,并且有11份来源不同的资源遗传结构复杂,但总体距离聚类结果与模型聚类分析结果较为一致。
本研究对藜蒿材料进行亲缘关系和遗传多样性进行分析,然而,由于RAPD分子标记重复性和稳定性差,具有一定的局限性[25],并且本次试验各个种群采样不均,部分地区收集的藜蒿样品材料不够丰富,仅用1~2个样本作为种群代表进行分析难免会造成数据结果的偏差[26],不能完全概括藜蒿种群间的遗传关系。因此,之后的研究中需要增加不同种群的样本丰富度和均一度,采用多种分子标记技术,设计特异性引物,结合藜蒿的一些表型性状、生物学特性,在分子水平上进行综合鉴定亲缘关系,探索不同种群材料的遗传变异模式、遗传多样性和亲缘关系等信息,这样相比单一的分子标记信息更丰富[27]。
猜你喜欢
今日农业(2022年13期)2022-09-15 01:18:00
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:33:26
中国麻业科学(2018年6期)2018-04-09 11:22:12
西南农业学报(2016年5期)2016-05-17 05:42:21
系统工程与电子技术(2016年2期)2016-04-16 05:16:53
西南农业学报(2016年6期)2016-04-16 05:12:47
广西林业科学(2016年3期)2016-03-16 05:43:21
法医学杂志(2015年4期)2016-01-06 12:36:36
中国光学(2015年1期)2015-06-06 18:30:20
河南医学研究(2014年7期)2014-02-27 14:53:42
