
解淀粉芽孢杆菌YA289 降解虾头蛋白的发酵工艺优化
2022-11-08 01:56:12李雨霖倪婕薛璐瑜杨冬梅余炼刘小玲
现代食品科技 2022年10期
李雨霖,倪婕,薛璐瑜,杨冬梅,余炼*,刘小玲
(1.广西大学轻工与食品学院,广西南宁 530004)(2.桂林师范高等专科学校,广西桂林 541199)
对虾养殖业是我国水产养殖的代表性产业之一,2019 年产量为487.141 t[1],养殖总产量可占全球对虾养殖总产量的26%[2]。目前以无外骨骼冷冻形式生产的产品需求量逐渐增加,而在该加工过程中大约会有40%~50%的虾头、虾尾、虾壳等产生[3],其中虾头含有较为丰富的蛋白质、脂质、甲壳素等多种营养成分。这些副产物通常被丢弃导致资源浪费,少部分进行了工业处理,如从副产物中提取甲壳素,但所使用的化学试剂易造成环境污染且生产过程流失了大量蛋白质资源。综上,有必要对虾头中蛋白质的提取降解技术进行研究。
现有研究表明可以通过发酵、酶解、超临界二氧化碳萃取[4]等方式对虾头中的蛋白质进行回收利用[5]。微生物可以在生长繁殖过程中分泌大量的蛋白酶及各种内肽酶从而降解蛋白质,生成易被动植物消化吸收的多肽和游离氨基酸,且微生物发酵后多肽和游离氨基酸的产量可能高于酶水解[6]。用其制作氨基酸液体肥料,可提高农作物的产量和品质。梁田[7]、王爽[8]的研究表明蛋白在降解中伴随脱氨基的过程,部分转化为氨气从发酵培养基中释放出来,产生大量不愉快气体,既造成了原料中氮元素的浪费[8],也是发酵过程中恶臭形成的原因之一。这一问题在一定程度上限制了微生物发酵方式的进一步应用,而目前对于该问题解决的相关报道较少。
目前,芽孢杆菌[9]、酵母菌[10]等微生物被证实具有一定降解氨态氮的能力,在降低水体氨态氮含量方面有良好的前景[11]。解淀粉芽孢杆菌Bacillus amyloliquefaciens是芽孢杆菌属,其可从土壤、植物、畜禽和水生动物的肠道中被分离培养,营养来源广泛、环境适应能力强、非致病性,常用于谷物和豆类、酒的发酵剂、加工或农业方面的抗菌剂等[12],在益生菌添加剂等领域发挥着重要的作用。本团队从北海红树林中分离了一株B.amyloliquefaciensYA289,前期实验表明其蛋白酶活性较强,具有良好降解虾头蛋白质的能力,且发酵过程中产生的氨态氮含量较低,氨臭味不明显。因此本文以氨基酸态氮、多肽和氨态氮含量为指标,采用B.amyloliquefaciensYA289 对虾头蛋白质的培养基及发酵工艺条件进行优化,为促进虾头副产物资源的再利用提供一定理论参考和技术支持。
1 材料与方法
1.1 材料与试剂
1.1.1 材料
虾头:南美白对虾虾头由广西北海正午有限公司提供。
菌株:本研究所使用菌株YA289 由本团队分离自广西北海红树林生长区,经16S rRNA测序鉴定YA289与Bacillus amyloliquefaciensDSM 7 具有99.86%的同源性,初步鉴定该菌株为解淀粉芽孢杆菌。
1.1.2 培养基
ISP2固体培养基(g/L):麦芽粉2,酵母粉2,葡萄糖2,琼脂15。
LB 液体培养基(g/L):蛋白胨10,酵母粉5,NaCl 10。
基础发酵培养基:称取清洗并沥干水的虾头与水1:8(m/V)匀浆,在121 ℃条件下灭菌20 min。
1.1.3 试剂
氢氧化钠、盐酸、氯化钠、硫酸铜、苯酚,成都金山化学试剂有限公司;尿素、乳糖、果糖、牛血清蛋白,北京索莱宝科技有限公司;牛肉膏、蛋白胨,北京奥博星生物技术有限责任公司;蔗糖、葡萄糖、淀粉、酒石酸钾钠、硫酸铵、乙酰丙酮、亚硝基铁氰化钠、结晶乙酸钠,天津市大茂化学试剂厂;柠檬酸三钠、次氯酸钠、冰乙酸、甲醛,成都市科隆化学品有限公司;琼脂、麦芽粉、酵母粉,德国BioFrox 公司。
1.2 仪器与设备
BS-600L 电子天平,北京赛多科利斯天平有限公司;SUNRISE 酶标仪,帝肯(上海)贸易有限公司;DGH-9140A 电热恒温鼓风干燥箱,上海精宏实验设备有限公司;SW-CJ-2F 洁净工作台,苏州苏净安泰公司;GI80TW 高压蒸汽灭菌锅,致微(厦门)仪器有限公司;LRH-70 生化培养箱,上海一恒仪器有限公司;ZWY-2102C 振荡培养箱,上海智城分析仪器制造有限公司;CR21N 高速冷冻离心机,日立公司;PHS-3BW pH 计,上海般特仪器制造有限公司。
1.3 实验方法
1.3.1 菌种的活化与菌悬液的制备
将B.amyloliquefaciensYA289 接种于ISP2培养基,于37 ℃培养箱中倒置培养24 h。挑取1环活化好的菌体接种于装有100 mL LB 液体培养基的250 mL三角瓶中,在200 r/min、37 ℃的条件下培养24 h,取菌液离心(4 000 r/min,10 min)收集沉淀,并于0.9wt%的无菌生理盐水中重悬,并调整其OD600nm在0.7 左右。
1.3.2 单因素试验设计
1.3.2.1 发酵培养基优化
以接种量5wt%、初始pH 值8、发酵温度37 ℃、转速200 r/min、发酵时间48 h 为基础发酵条件,对基础发酵培养基中几种重要成分及其浓度进行筛选优化。设置不同碳源成分(淀粉、葡萄糖、蔗糖、果糖和乳糖,添加质量浓度均为5 g/L)、不同葡萄糖质量浓度(0、5、10、15、20 g/L)、不同氮源成分(硫酸铵、牛肉膏、蛋白胨、酵母粉、尿素,添加质量浓度均为5 g/L)、不同氯化钠质量浓度(0、5、10、15、20 g/L),每组设3 个平行,待发酵结束后取发酵液离心(10 000 r/min,5 min),测定发酵上清液中氨基酸态氮、多肽和氨态氮的质量浓度,结果取平均值(下同)。
1.3.2.2 发酵条件优化
以发酵培养基优化的试验结果(即在基础虾头发酵培养基中添加5 g/L 葡萄糖及5 g/L NaCl)作为发酵条件优化的培养基。设置不同接种量(1wt%、3wt%、5wt%、7wt%、9wt%),不同发酵初始pH 值(2、4、6、8、10),不同料液比[1:2、1:4、1:6、1:8、1:10(g/100 mL)],不同发酵时间(24、48、72、96、120 h),不同发酵温度(28、31、34、37、40 ℃),不同发酵转速(140、160、180、200、220 r/min),根据发酵上清液中氨基酸态氮、多肽和氨态氮的质量浓度确定最佳发酵条件。
1.3.3 Plackett-Burman(PB)试验设计
在发酵条件单因素实验的基础上,分别选取接种量、pH 值、料液比、发酵时间、发酵温度和发酵转速这6 个因素的高(1)和低(-1)两个水平进行试验设计,筛选出对发酵上清液中多肽质量浓度显著影响的因素。试验因素及水平编码如表1 所示。
1.3.4 最陡爬坡试验设计
选择PB 试验筛选得到的4 个显著因子,以多肽质量浓度为响应值进行最陡爬坡试验,参考各显著因子的正负效应值确定试验的方向和步长。以最大响应值对应的工艺参数为Box-Behnken 试验设计的中心点,试验设计如表3 所示。
1.3.5 Box-Behnken 试验设计
在PB 试验和最陡爬坡试验确定的4 因素3 水平的基础上,利用Design-Expert 12 软件进行Box-Behnken试验的设计和数据分析,试验因素与水平见表4。其他影响因素的条件为接种量5%,转速180 r/min。
1.3.6 氨基酸态氮质量浓度的测定
参照GB 5009.235-2016《食品安全国家标准 食品中氨基酸态氮的测定》,采用比色法测定发酵上清液中氨基酸态氮的质量浓度。
1.3.7 多肽质量浓度的测定
参考李伟民[13]的方法,取1 mL发酵上清液以1:1(V/V)的比例与10%三氯乙酸混匀,静置10 min,5 000 r/min 离心15 min,取1 mL 上清液和4 mL 双缩脲试剂,混匀,室温下静置30 min,在540 nm 测定吸光值,以蒸馏水作空白。
1.3.8 氨态氮质量浓度的测定
参考梁剑光[14]的方法,取发酵上清液稀释适当倍数后加显色剂混匀,置于37 ℃孵育20 min 显色,待反应结束后冷却至室温,在637 nm 测定其吸光值,以蒸馏水作空白。
1.3.9 数据统计与分析
试验数据分析采用 Minitab 17、IBM SPSS Statistics 26 和Origin 2021,结果均以平均值±标准差的形式呈现。
2 结果与分析
2.1 单因素试验
2.1.1 发酵培养基优化
2.1.1.1 碳源的优化
碳源不仅能提供微生物生长所需的能量,还可以提供菌体合成目的产物所需的原料,减少对氨基酸的消耗[15]。由图1 可知,以无外加碳源的基础发酵培养基为对照组,分别添加葡萄糖、果糖、蔗糖和乳糖后,发酵上清液的氨基酸态氮和多肽质量浓度都有不同程度的增加,其中以添加葡萄糖或果糖后增幅较明显,说明对于B.amyloliquefaciensYA289 而言,葡萄糖或果糖优先于氨基酸被作为碳源利用,这可能是由于葡萄糖或果糖较有利于促进B.amyloliquefaciensYA289 分泌蛋白酶,并进而分解虾头中的蛋白质产生更多的多肽与氨基酸。孙力军等[16]和Chen 等[17]的研究也表明葡萄糖对解淀粉芽孢杆菌抗菌脂肽的合成有明显的促进作用。另外,图1 显示添加蔗糖和乳糖后与对照组之间无显著性差异(p>0.05),而添加淀粉后氨基酸态氮和多肽质量浓度都相较对照组有所下降;添加果糖、葡萄糖及乳糖时氨态氮产量较高,而添加蔗糖和淀粉时氨态氮产量则较低。宋君等[18]的研究也表明在外加蔗糖时Bacillus pumilusBP-171有较好的降解无机氮的效果,与本研究结果符合。这可能是由于不同碳源的空间结构和分子量不同,使其被利用的难易程度有差异,所以不同碳源下菌株的生长状况和脱氮能力也不同。因此在培养基中添加葡萄糖以获得更高的氨基酸和多肽产量,同时相对果糖有更低的氨态氮损失。

图1 碳源对氨基酸态氮、多肽和氨态氮质量浓度的影响Fig.1 Effect of carbon source on amino acid nitrogen,polypeptide and ammonia nitrogen content
2.1.1.2 葡萄糖添加量
由图2 可知,随着葡萄糖添加量的增加,氨基酸态氮、多肽和氨态氮的质量浓度都呈现先增加后减少的趋势,在5 g/L 和10 g/L 时,氨基酸态氮质量浓度都达到最高且氨态氮质量浓度较低,其中多肽质量浓度在5 g/L 时有最大值,这与LEI 等[19]用Bacillus licheniformisSWJS33 发酵豆粕的研究结果一致,可能的原因是适当的葡萄糖添加量有利于微生物生长和产酶,添加过多会加速菌体的呼吸作用,过度消耗培养基中的溶氧,导致其自身生长和产酶受到抑制。因此,选择5 g/L 的葡萄糖添加量进行后续试验。

图2 葡萄糖添加量对氨基酸态氮、多肽和氨态氮质量浓度的影响Fig.2 Effects of glucose addition amount on amino acid nitrogen,polypeptide and ammonia nitrogen content
2.1.1.3 氮源的优化
由图3 可知,以不外加氮源的基础虾头发酵培养基为对照组,当外加不同氮源后氨态氮的质量浓度都有不同程度的增加,这可能是由于碳氮比变化造成的影响[20]。添加尿素和酵母粉后氨基酸态氮质量浓度升高,但与对照组无显著性差异(p>0.05),而添加其他氮源后氨基酸态氮质量浓度均低于对照组。外加硫酸铵发酵后多肽质量浓度最高,其次为尿素,其他外加氮源多肽质量浓度反而出现下降。MAO 等[21]以虾头作为Bacillus licheniformisOPL-007发酵唯一氮源的试验结果表明该菌株可以直接从虾头中获得氮,满足自身生长需求和脱蛋白作用。综上,推测以虾头作为氮源已满足B.amyloliquefaciensYA289 基本的生长和产酶需要,再添加其他氮源还会导致资源浪费,因此培养基中以不外加氮源为适宜。

图3 氮源对氨基酸态氮、多肽和氨态氮质量浓度的影响Fig.3 Effect of Nitrogen source on amino acid nitrogen,polypeptide and ammonia nitrogen content
2.1.1.4 NaCl 添加量
无机盐可以调节渗透压,提供微生物生长必须的离子,还可调节生理活性作用或作为生理活性物质的组成部分[22],同时无机盐浓度对产酶的影响也十分关键。由图4 可知,发酵上清液中氨基酸态氮的质量浓度随NaCl 浓度的增加呈现先上升后下降的趋势,在NaCl 质量浓度为5 g/L 时氨基酸态氮质量浓度最高,这说明该B.amyloliquefaciensYA289 具有一定的耐盐性。而NaCl 添加量对多肽和氨态氮的质量浓度没有显著影响(p>0.05)。袁艳超[23]的实验结果表明B.amyloliquefaciensY1 在添加0.3%的NaCl 时蛋白酶活力最高,且酶活力也呈先上升后下降的趋势,因此,选择5 g/L 的NaCl 添加量进行后续试验,该浓度下氨基酸态氮产量较高且氨态氮质量浓度较低。

图4 NaCl 添加量对氨基酸态氮、多肽和氨态氮质量浓度的影响Fig.4 Effect of NaCl addition amount on amino acid nitrogen,polypeptide and ammonia nitrogen content
2.1.2 发酵条件优化
2.1.2.1 接种量
接种量的大小影响菌体生理状态及发酵的周期和产物的合成[24]。由图5 可知,随着接种量增加,氨态氮质量浓度和多肽质量浓度均有增加,但没有显著性差异(p>0.05);氨基酸态氮质量浓度逐渐增加,当接种量上升到5%时氨基酸态氮和多肽质量浓度最高,接种量继续增加后氨基酸态氮质量浓度下降,这与宋敏[25]的研究结果一致,其原因可能是接种量的增加缩短了微生物的延滞期,菌体增多,促使了蛋白酶分泌增加,多肽质量浓度和氨基酸态氮质量浓度都逐渐升高;而接种量过大菌体大量生长繁殖,可能导致发酵后期营养不足、某些初级或次级代谢产物累积增多,从而抑制产酶[26,27],导致氨基酸态氮质量浓度下降。因此,选择5%的接种量进行后续试验。
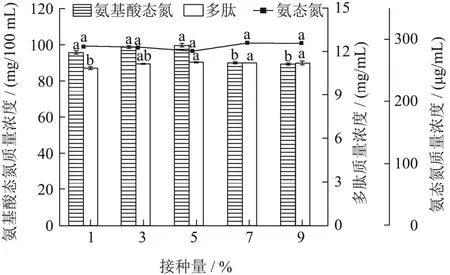
图5 接种量对氨基酸态氮、多肽和氨态氮质量浓度的影响Fig.5 Effect of inoculum on amino acid nitrogen,polypeptide and ammonia nitrogen content
2.1.2.2 pH 值
微生物在生长代谢过程中会引起培养环境pH 值的变化,pH 值过高或过低都会影响其进一步的生长和产物的合成,同时也是影响NH3挥发的关键因素之一。由图6 可知,随着pH 值升高氨基酸态氮和多肽的质量浓度都逐渐增加,在pH 值为6 时有最大值,随着pH 值继续升高,氨基酸态氮和多肽质量浓度反而下降,与Wang 等[15]有关枯草芽孢杆菌的水解度趋势一致。氨态氮质量浓度在中性和碱性环境下较低,这与王涛[11]和宋君[18]的研究结果一致,可能是因为细菌在中性和偏碱性环境下反硝化酶的活性较强[28],对氨态氮的降解率更高。虽然pH 值为6 时氨态氮的质量浓度与pH 值为8 时有显著差异,但无令人不愉快的气味,综合氨基酸态氮和多肽质量浓度选择pH 值为6进行下一步试验。

图6 pH 对氨基酸态氮、多肽和氨态氮质量浓度的影响Fig.6 Effect of pH on amino acid nitrogen,polypeptide and ammonia nitrogen content
2.1.2.3 料液比
料液比的大小会影响菌种发酵虾头副产物的效果,如图7 所示,在一定范围内随着料液比逐渐增大,即虾头浓度逐渐减少,氨基酸态氮质量浓度没有显著性变化(p>0.05),多肽质量浓度先增加后减少,这是因为料液比过大菌种不能完全发挥作用,可能会进一步分解生成的营养物质,导致发酵效率降低,而料液比过小会导致虾头不能被充分分解。在周莲[29]利用B.amyloliquefaciens3-2 降解羽毛的研究中,一定范围内羽毛含量的增加使羽毛降解率也随之增加,超过这个范围后羽毛降解率也逐渐降低。氨态氮质量浓度随着料液比减小而减少,在1:6、1:8 和1:10(g/100 mL)处均无令人不愉快的气味,因此综合氨基酸态氮和多肽质量浓度选择料液比1:6(g/100 mL)进行下一步试验。

图7 料液比对氨基酸态氮、多肽和氨态氮质量浓度的影响Fig.7 Effect of solid-to-liquid ratio on amino acid nitrogen,polypeptide and ammonia nitrogen content
2.1.2.4 发酵时间
发酵时间影响微生物的生长、蛋白酶的分泌和蛋白质的降解及氨基酸的消耗[15]。由图8 可知,随着发酵时间的增加,氨基酸态氮质量浓度和多肽质量浓度都随之增加,在48 h 时达到最大值,与Moayedi 等[6]和Wang 等[15]的趋势一致。产物氨基酸的积累是综合蛋白质降解所产生的和微生物生长所消耗的动态变化的结果。发酵时间过长使菌体开始衰亡、产酶减少,同时由于后期营养不足,微生物也会利用已生成的多肽等物质进行自身的生长繁殖[15]。而发酵时间对氨态氮质量浓度无显著影响(p>0.05),在48 h 处质量浓度最低,综上,选择48 h 发酵时间进行下一步试验。

图8 发酵时间对氨基酸态氮、多肽和氨态氮质量浓度的影响Fig.8 Effect of fermentation time on amino acid nitrogen,polypeptide and ammonia nitrogen content
2.1.2.5 发酵温度
微生物有自身适宜的生长温度,因而发酵温度能影响其生长和代谢过程。由图9 可知,当温度上升时,氨基酸态氮质量浓度变化无显著性差异(p>0.05),多肽质量浓度和氨态氮质量浓度增加。在Wang 等[15]的研究中,枯草芽孢杆菌的水解度也随着温度的升高而升高,在35 ℃时达到最大值。综上,由于34 ℃与37 ℃的氨基酸态氮和多肽质量浓度没有显著差别,但34 ℃时氨态氮质量浓度较低,因此选择34 ℃的发酵温度进行下一步试验。

图9 发酵温度对氨基酸态氮、多肽和氨态氮质量浓度的影响Fig.9 Effect of fermentation temperature on amino acid nitrogen,polypeptide and ammonia nitrogen content
2.1.2.6 发酵转速
Lei 等[19]的研究表明,温度和溶氧可以控制细菌中酶的合成与能量代谢之间的联系。由图10 可知,与Wang 等[30]的研究结果趋势保持一致,随着转速增加氨基酸态氮质量浓度先增加后减少,转速为180 r/min时氨基酸态氮最高。这是由于随着转速的增大溶氧量增加,利于菌的生长,而转速过高可能会造成菌体机械性损伤而影响生长,甚至改变酶结构[31]。转速的改变对多肽质量浓度的影响不显著,当转速超过180 r/min 后氨态氮质量浓度显著增加,因此选择180 r/min 的发酵转速进行下一步试验。

图10 发酵转速对氨基酸态氮、多肽和氨态氮质量浓度的影响Fig.10 Effect of fermentation speed on amino acid nitrogen,polypeptide and ammonia nitrogen content
2.2 Plackett-Burman(PB)试验
对发酵条件单因素试验中的6 个因素利用Minitab 软件进行显著性分析,试验结果如表1 所示。对表1 试验结果进行分析可得到回归模型方程:

表1 Plackett-Burman 试验设计及响应值(n=12)Table 1 Results of Plackett-Burman experimental design
多肽=10.657-3.221A+0.209B+1.197C-0.886D-0.965E-0.953F,R2=95.57%
说明模型拟合良好。由表2 的显著性分析结果可知,料液比(A)、pH(C)、温度(E)、时间(F)对多肽质量浓度均有显著性影响(p<0.05),其中pH 值具有正效应,料液比、温度和时间具有负效应,选择这4 个显著因子进行下一步最陡爬坡试验。

表2 Plackett-Burman 实验结果显著性分析Table 2 Significance test of factors for Plackett-Burman test
2.3 最陡爬坡试验
由表3 试验结果可知,随着料液比、pH 值、温度、时间条件的变化,多肽质量浓度呈现先上升后下降的趋势,在第3 组取得较好结果,故以其作为中心点进行下一步Box-Behnken 试验。

表3 最陡爬坡实验设计及结果Table 3 Experimental design and results of steepest climbing
2.4 Box-Behnken 试验
对表4 的试验结果进行二次多项式回归拟合,可得多肽质量浓度(Y)的回归方程:
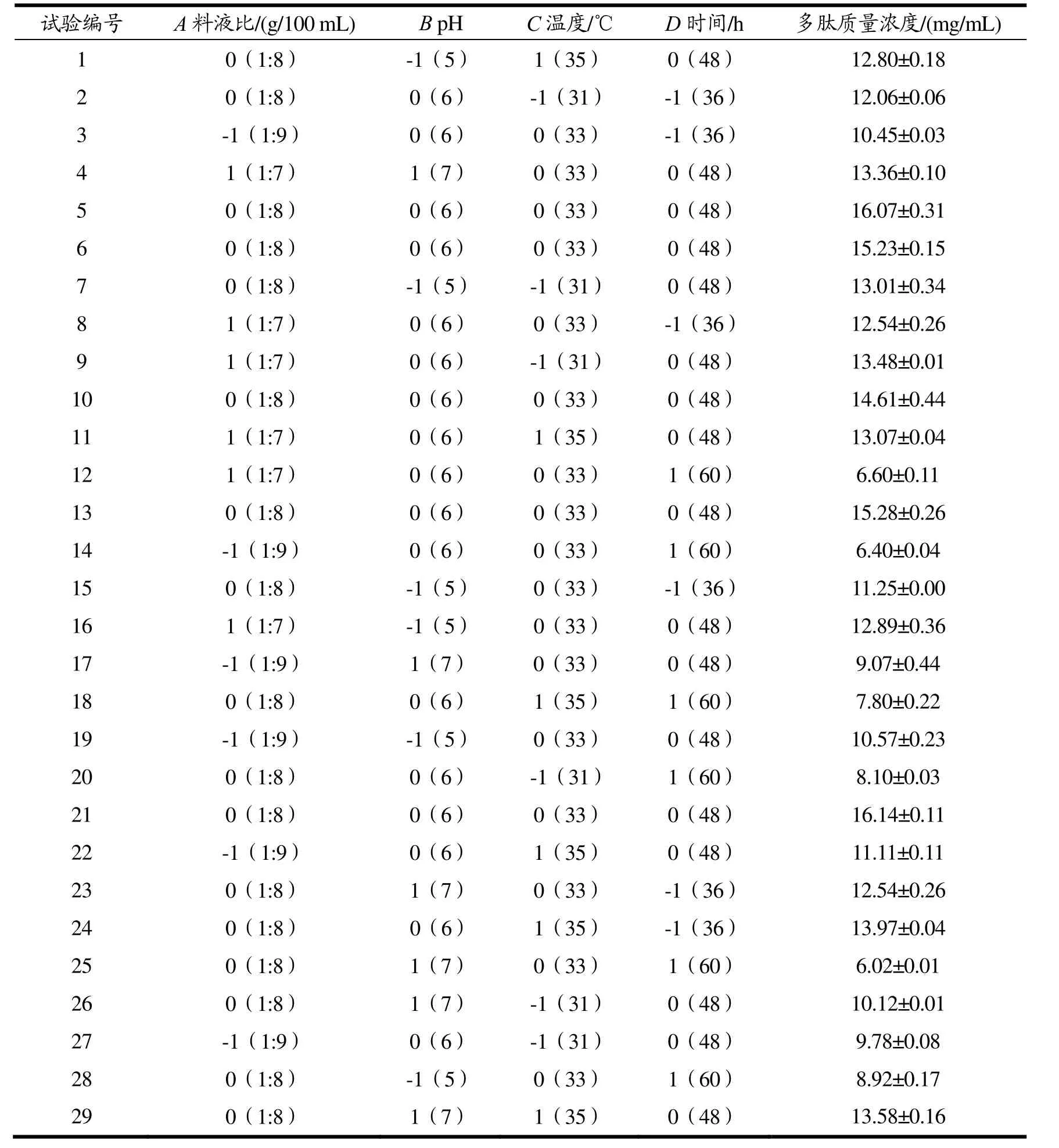
表4 Box-Behnken 试验设计和结果Table 4 Design and results of Box-Behnken experiment

对回归模型进行方差分析,结果如表5 所示,该模型显著(p<0.0001),失拟项不显著,模型的R2=0.975 7,校正后R2adj=0.951 5,信噪比=20.980 1>4,以上表明该模型拟合程度较好,误差较小,可以较准确的预测响应值的变化。根据方差分析结果的p值可知各因素对多肽质量浓度影响大小的依次顺序为:D>A>C>B,即时间>料液比>温度>pH 值。

表5 回归模型的方差分析结果Table 5 Analysis results of the regression model
通过Design-Expert 12 软件得到料液比、pH 值、温度、时间这4个因子之间的响应曲面图如图11所示。曲面图颜色深浅的变化表示多肽质量浓度的变化,变化越快则坡度越陡,即试验因素对响应值影响更显著,由图可知,多肽质量浓度分别随pH 值、料液比、温度和时间的增加都呈现先增加后降低的趋势,在各因素的中心水平有较高值,各因素之间存在明显的交互作用。综上所述,根据建立的模型预测,当4 个影响因子分别在A=1:7.6、B=6.10、C=33.55、D=43.76 时,有最佳响应值(16.11 mg/mL)。为了验证该模型预测结果的可靠性,在考虑实际操作的条件设置后,将预测的试验条件改为料液比1:8 (g/100 mL)、pH 值6、温度34 ℃、时间44 h 进行了3 次验证试验,测得的多肽质量浓度为16.35 mg/mL,与预测值接近,说明该模型准确性良好,优化所得的工艺条件参数可靠。

图11 各因素交互作用响应曲面图Fig.11 Response surface diagram of interaction of various factors
3 结论
利用B.amyloliquefaciensYA289 降解废弃虾头中的蛋白质相较于成本较高的酶解法,是一种更为环保、经济的方法,可作为未来实现对虾副产物资源化利用的方向进一步研究。本研究在单因素试验的基础上,结合了Plackett-Burman 试验和Box-Behnken试验优化了B.amyloliquefaciensYA289 发酵虾头的工艺条件,确定了其最佳的发酵条件,即在葡萄糖添加量5 g/L,NaCl 添加量5 g/L,料液比1:8(g/100 mL),接种量5%,pH 值6,转速180 r/min,温度34 ℃,时间44 h的条件下,虾头发酵上清液中多肽的质量浓度为16.35 mg/mL,比优化前提高了2.7 倍,其发酵效果优于Sun 等[32]用Bacillus subtilisOKF04 发酵南极磷虾得到的多肽质量浓度(9.5 mg/mL)。因此发酵需要综合选取合适的菌株、培养基成分和发酵条件,才能更有效提升发酵效果。但是本研究存在采用的三角瓶摇床发酵存在容量小等不足,后续研究仍需进行放大试验验证确定符合实际生产的参数,同时还可以从现代诱变育种技术和复合菌株的角度出发,进一步提高发酵效果。
猜你喜欢
天津农学院学报(2023年5期)2023-12-25 10:34:28
天津师范大学学报(自然科学版)(2023年5期)2023-12-03 09:13:28
环保科技(2020年1期)2020-03-04 04:37:28
山西建筑(2016年23期)2016-11-03 02:45:46
水科学与工程技术(2016年2期)2016-07-10 10:19:57
中国酿造(2016年12期)2016-03-01 03:08:11
中国酿造(2014年9期)2014-03-11 20:21:03
中国酿造(2014年9期)2014-03-11 20:21:03
食品工业科技(2014年9期)2014-03-11 18:15:28
养殖与饲料(2014年10期)2014-02-28 22:15:02
