
纳米抗体在口蹄疫防治中的研究进展
2022-11-07 02:10靳慧茹武艺贾婷张宏亮王申元凌宇刘羿羿刘春霞周欢敏李璐张焱如曹俊伟
中国动物检疫 2022年11期
靳慧茹,武艺,贾婷,张宏亮,王申元,凌宇,刘羿羿,刘春霞,周欢敏,李璐,张焱如,曹俊伟
(1.内蒙古农业大学生命科学学院,内蒙古自治区生物制造重点实验室,内蒙古呼和浩特 010018;2.赤峰市元宝山区农牧局,内蒙古赤峰 024076)
口蹄疫(foot and mouth disease,FMD)是由口蹄疫病毒(foot and mouth disease virus,FMDV)引起的高度接触传染性疫病,侵染对象为猪、牛、羊等偶蹄类动物[1]。FMD 发病率高,传播途径广,常引发幼畜患心肌炎而猝死,是威胁偶蹄动物健康的主要疾病之一。FMD 曾在欧洲、亚洲、非洲以及南美洲地区流行[2],其最具代表性的临床特点为口腔黏膜以及蹄部和乳房皮肤发生水疱、糜烂或溃疡性病变。FMDV 基因组是一个单链RNA,长约8.5 kb。FMDV 由4 种(VP1—VP4)结构蛋白组成的蛋白质外壳包裹,共有7 种血清类型,分别为A、O、C、Asia1 以及SAT1、SAT2 和SAT3,各血清型间无交叉免疫保护[3]。接种灭活疫苗是预防FMD 的主要措施之一。完整病毒粒子(146S)和空衣壳(75S)可以使疫苗具有较好的免疫保护力,而这两种颗粒在疫苗制造、配制和储存过程中都可能被解离成12S 亚单位,导致疫苗效力降低[4]。由于目前尚缺乏能够长期有效且完全抑制FMDV 的药物或疫苗,因此如何研究出高效的抗FMDV 药物仍需不断探索。
纳米抗体,一种存在于驼科动物体内仅含有重链可变区的特异性抗体。这些特异性抗体可以通过自身单个结构域进行抗原识别,最初被命名为VHH(variable domain of the heavy chain of heavychain antibody),而后根据其相对分子质量极小的特点被命名为纳米抗体(nanobody,Nb)[5]。Nb可与酶、底物、放射性物质和其他生物活性物质连接形成融合的Nb 以表达特定的功能效应[6-7]。相比传统单克隆抗体(monoclonal antibody,mAb),Nb 结合抗原的能力优于mAb。Nb 抗原结合系统非常广泛,可以与其他抗体片段通过巢式PCR 技术扩增获得中和能力更强的双特异性Nb,创建具有高亲和力和高效力的多价分子[8-9]。
Nb 因独特的结构成为对抗病毒的抗体之一,为病毒性疾病的诊断和治疗开辟了新天地,目前在兽医学领域应用范围很广,许多RNA 病毒疾病的治疗方向已被转入Nb 开发中。近年来,随着生物技术的不断发展,各领域对Nb 的研究愈发深入。Nb 不仅可以作为探针来诊断疾病,将药物传递到机体内部,还可以作为免疫抑制剂或治疗药物来中和病毒毒力,且Nb 的制备也可以采用噬菌体筛选技术快速获得[10]。目前已经有许多Nb 类药物被批准上市,应用于炎症性疾病、肿瘤等各种疾病的诊疗中,在未来的研发中Nb 将会发挥更大的作用。本文主要对Nb 的优势及其目前在FMD 防治中的应用等相关内容进行综述并作展望。
1 Nb 的结构
1993 年Hamers-Casterman等[11]在骆驼体内首次发现一种只有重链,缺乏轻链和第一个恒定 区CH1(conventional region of heavy chain 1)的重链抗体(heavy-chain antibody,HCAb),克隆其可变区得到的单域抗体即为Nb。Nb 是一种新型天然抗原特异性结合功能片段(antigenbinding fragments,ABF),也是目前已知的最小ABF[12]。Nb 可以通过基因工程改造融合后获得新的结构模块,从而实现多特异性和多功能性[13]。Nb 晶体呈椭圆形,短轴只有2.5 nm,长轴4 nm,相对分子质量为12~15 kDa,大小只有传统mAb(约150 kDa)的10%左右,单链可变片段(single chain variable fragments,scFv,约30 kDa)的50% 左右,抗原结合片段(约50 kDa)的33% 左右[14-15]。各类抗体相对分子质量示意图见图1[16]。Nb 由3 个抗原互补决定区(complementarity determining region,CDR)和4个框骨架区(framework region,FR)组成。其中CDR3 会与CDR1、CDR2 或FR2 之间通过二硫键相连,进而形成一种稳定结构,这有助于CDR3 凸形结构的形成。研究[16-17]表明,CDR3 通常采用α-螺旋构象进行抗原识别,是抗原结合的主要位点。
2 Nb 的特性
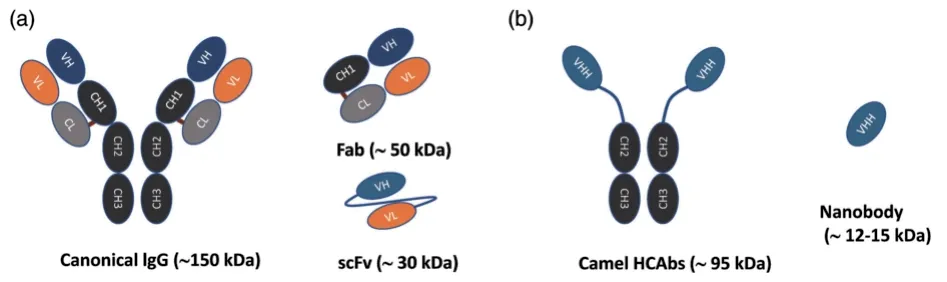
图1 各类抗体相对分子质量示意图
骆驼的VHH(Nb)相比传统mAb 单域可变区(heavy chain variable region,VH)而言,二者同源性可达到80%(图2)。与VH 相似,骆驼VHH(Nb)包含9 个β-链组成典型的IgV 折叠,但其免疫原性较低,且轻链可变区(VL)的缺失导致两种片段在功能上存在显著差异[18]。在传统的VH 区域中,FR2 片段是由4 个高度保守的疏水氨基酸(Val37、Gly44、Leu45 和Trp47)组成,这些氨基酸可以与Gln39、Gly44、Tyr91 和Trp103一起形成一个保守的疏水界面。而在VHH(Nb)中,为了避免如此庞大的疏水区暴露在溶剂中,其FR2片段则由更亲水的氨基酸(Phe37、Glu44、Arg45和Gly47)组成,且其对温度和pH 变化表现较稳定,表明VHH(Nb)溶解性更好,稳定性更强[19-22]。
VHH(Nb)与VH 的CDR3 环长度也存在明显差异。VHH(Nb)的CDR3 长度超过20 个氨基酸残基,而传统抗体可变区VH 的典型氨基酸长度仅为9 个(小鼠)或12 个(人)。VHH(Nb)的CDR1 和CDR3 氨基酸序列较长,其形成的凸环结构可深深嵌入到抗原分子的凹槽或缝隙中,以增大与抗原相互接触和作用的表面积,也相对弥补其因缺少轻链而丧失的抗原结合能力。而CDR1 和CDR2 由于在结构上容易弯折,非常便于免疫球蛋白与抗原表位相结合,因此可以增强抗原抗体的结合能力[23-26],使VHH(Nb)具有较高的亲和力及识别能力。
在目前的抗体治疗领域中,抗体大小可以决定其治疗效果[27]。由于Nb 体积小,且具有高组织渗透性,它们能够快速渗透到组织中,使其或相关药物的作用发挥到极致,从而避免机体被病毒感染[28]。同时,Nb 分子结构简洁,更易于被克隆与基因修饰,可在各种细胞中重组表达,也可在大肠杆菌和酵母菌等微生物表达系统中自由表达,这为工业化大规模生产奠定了基础。因Nb 缺少传统抗体的Fc(fragmentcrytallizable)片段,所以避免了Fc 段可能引起的补体反应,使其对人体的免疫原性很低,从而表现出较好的生物相容性[29]。
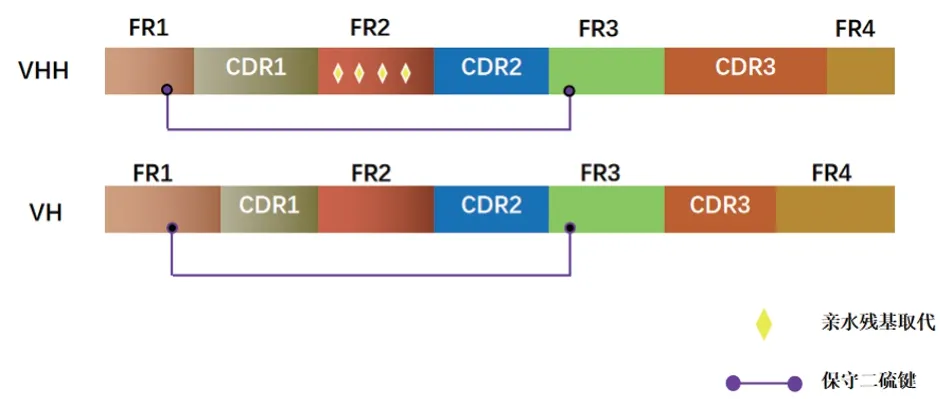
图2 VHH 和VH 结构示意图[30]
3 Nb 在FMD 诊断中的应用
基于FMDV 感染特性,研究者们非常重视开发新的早期诊断方法。血清检测是动物进出口认证、动物“无感染”状态确认以及证明疫苗有效性所必需的,因此被认为是FMD 诊断方法的重要组成部分[31]。目前针对FMD 诊断已经研发出几种酶联免疫吸附试验(enzyme-linked immunosorbent assay,ELISA),但需要多次评估其特异性、灵敏性及重复性,且试验抗体耗费较大,使得一些发展中国家无法承担。利用Nb 生产成本低等一系列优点,Gelkop等[32]针对乌干达牛群FMDV 非结构蛋白(non-structural proteins,NSP)3ABC 的Nb,开发了一种新型竞争ELISA 方法,用于血清检测。他们将O 型FMDV 的3ABC 蛋白成功纯化表达,注射到骆驼体内,使其产生免疫抗体,构建噬菌体展示文库,经淘洗、筛选得到与目的蛋白亲和力较高的Nb;利用自制的基于Nb 的竞争性NSPELISA 来检测FMDV 3ABC 蛋白的循环抗体水平,分析其诊断敏感性(diagnostic sensitivity,DSE)和诊断特异性(diagnostic specificity,DSP),并与PrioCHECK 商业化的FMDV NSP 检测结果进行比较,结果发现两种分析方法具有高度一致性,表明研发方案可应用于FMD 诊断,可大大节约检测成本。此外,另有研究[33-34]将FMDV O 型(INDR2/1975)、A 型(IND40/2000)和Asia1型(IND63/1972)疫苗株灭活全病毒作为抗原免疫骆驼,制备FMDV 特异性Nb。通过构建抗FMDV的VHH 片段,使其在细菌中成功大量表达,经筛选浓缩后发现,在ELISA 检测中VHH 的表位鉴定可以充分捕捉抗体,代替传统兔血清成为包被抗体,成功应用于液相阻断ELISA(liquid-phase blocking ELISA,LPB-ELISA),并且证实建立的LPB-ELISA 与标准LPB-ELISA 相似性极高,而重组VHH 的生产也非常有助于减少样品检测成本。还有研究[35]利用双抗夹心法(double antibody sandwich,DAS)分离获得具有广谱识别能力的特异性Nb。该抗体可以定量FMD 疫苗中的完整衣壳蛋白,是监测疫苗生产、配制和储存过程中完整衣壳数量和稳定性的极佳工具。2017 年Harmsen[36]等在美洲驼的免疫文库中,利用噬菌体筛选技术分离到针对FMDV SAT2 和Asia1 型毒株的特异性Nb。这些特异性结合146S 颗粒的Nb 不会与其他FMDV 血清型毒株结合,故利用这一特点构建了新型ELISA。试验表明,这些特异性Nb 可以用作FMD 疫苗质量的控制与研发。根据前人的试验积累,王迪[37]使用FMDV 灭活苗免疫双峰驼,构建亲和力较高的Nb 免疫文库,筛选得到两株Nb,并证明这两株Nb 特异性较好,亲和力较高。之后他们使用量子点——一种半导体纳米微晶,对单域抗体进行偶联,发现间接免疫荧光方法可检测到BHK-21 细胞内的FMDV,这有利于检测细胞与病毒间的关系,为探索Nb 在FMD 检测诊断中的应用提供了新型思路。生物传感器即通过分子识别元件(生物活性物质)去结合被测目标,从而产生某种物理或化学变化,将此变化转换为可以被检测到的信号进行分析[38]。Nb 因具有高度特异性被广泛用作生物传感器。Trilling[39]等利用识别FMDV 的Nb 蛋白制作生物传感器,采用点击化学方法,将暴露环辛炔的表面等离子体共振(SPR)芯片与具有叠氮化物的VHH 结构域反应,结果发现生物传感器的灵敏度提高了800 倍,表明这项技术可能有助于加强对FMD 的控制和早期诊断。
总的来说,随着科技的进步,抗体已被证明是诊断方法开发的核心[40]。尽管单克隆抗体在诊断中发挥着重要作用,但它们的生产技术要求很高,而且价格昂贵。Nb 则因其良好的生物特性、物理化学特性以及生产成本相对实惠的优势被广泛应用于病毒检测。
4 Nb 在中和FMDV 中的作用
现如今FMD 的防御主要依赖于灭活病毒疫苗的接种。目前的疫苗针对FMDV 虽具有一定的保护性免疫反应,但是由于病毒复制能力强、传播速度快,或许已经超过了其免疫防御系统的清除速度,且FMDV 血清型间显著的抗原多样性,使开发具有抗病毒逃逸能力的中和抗体显得尤为重要。Harmsen等[41]通过筛选4 个大羊驼的噬菌体展示免疫库,鉴定出24 个能够在体外中和O 型FMDV的Nb,其中M8 和M170 的单个中和活性最强;将M8 和M23 混合免疫时起到的保护作用更显著,说明M8 和M23 会对FMDV 表现出较强的协同中和作用。2008 年Harmsen等[42]又将3 个结合FMDV 的Nb(M3、M8 和M23)与结合FMDV的猪IgM 分别进行融合,在体外获得了3 个双功能的中和抗体,这3 株双特异性Nb 抗体半衰期比单价Nb 延长了100 倍,在体外与单价抗体一样有效中和了FMDV,而且可有效减少病毒血症和排毒,抑制病毒传播。随着研究的进行,Harmsen等[43]发现,仅两种融合的VHH 对猪起到的保护作用时间短,因此将先前构建的两个VHH 的基因融合体(VHH2)与其他型号FMDV 的VHH 结合,得到两个Nb。实验结果表明两个Nb 共同作用要比单个Nb(VHH)更有效地中和FMDV,在猪FMD 激发感染前24 h 注射这两个Nb 的混合物可减少并延迟临床疾病、病毒血症和病毒脱落的发展。Dong等[44]为了研究M8 和M170 的血清型特异性或交叉反应性,通过ELISA 检测发现M170仅与O 型结合,而M8 可以和A、Asia1 和C 血清型结合;为了探索这两种抗体是否能同时结合病毒,他们进行了竞争性表面等离子体共振(surface plasmon resonance,SPR)分析,证明一种抗体的结合会阻断另一种抗体的附着,因此二者存在竞争关系;然后他们给豚鼠体内注射特异性抗体,通过检测豚鼠体内病毒载量来评估这两种抗体的保护效果,结果发现在服用治疗性Nb 之前,豚鼠体内可检测到高水平的病毒复制,而注射M8 和M170 后豚鼠体内病毒滴度大幅降低。通过结构分析得出,M8 和M170 会分别识别RGD 基序和HS 结合位点,阻止FMDV 附着到宿主细胞表面,具有阻断病毒突变逃逸的巨大潜力[45]。
自1930 年以来,人们一直在尝试开发FMDV强效疫苗。FMDV 疫苗接种历史记录了各种类型的疫苗,例如灭活病毒疫苗和空衣壳疫苗,减毒疫苗和重组疫苗[46-47]。但它们都存在局限性,如昂贵、保质期短、保护期短等问题[48]。目前关于FMD Nb 治疗方面的研究较少,而Nb 作为一种简单、经济、有效的工具,可以快速诱导机体产生中和反应,以对抗传染性疾病和难治性疾病,因此此类型抗体研发非常必要。
5 Nb 在FMD 治疗中的展望
FMDV 的聚合酶缺乏校对能力,使其基因组具有非常高的突变率[49]。目前我国流行的FMDV血清型主要为A、O 型。Nb 作为一种溶解度高、热稳定性强、亲和能力高和生产成本低的理想抗体,可以弥补单克隆抗体生产成本高、渗透力弱等一系列缺陷,即使在严苛的外界环境条件下,Nb 仍然可以保持原有的抗原结合能力,并且使疫苗效力与储存得到保障。利用骆驼的天然文库或免疫文库筛选得到的Nb 可以有效中和病毒,且其序列和长度的高度可变性使得它们可以识别各种蛋白质表位,包括裂隙中的隐藏表位,从而抑制或降低受体与病毒蛋白的结合作用,增强抗病毒药物的中和作用。多价结合的Nb 也有希望提高病毒中和效力。晶体结构的解析也可以更具体地分析抗原位点,为后期FMD 治疗提供有力依据。有研究[50]从SARS-CoV-2 刺突蛋白受体结合域(RBD)免疫的骆驼中鉴定到一组高亲和力Nb,通过晶体解析了与RBD 形成复合物的两个非竞争Nb(NB1A7和NB1B11)的结构,发现Nb 针对SARS-CoV-2变异株和其他冠状病毒共有的高度保守的隐蔽表位,阻断了刺突蛋白与ACE2 受体的结合,且这两种抗体连接形成的多价抗体显著提高了亲和力与中和力,可进一步抑制病毒逃逸。另有研究[51-52]发现,针对不同表位的单价和双价表皮生长因子受体(epidermal growth factor receptor,EGFR)的特异性Nb 和双价异型Nb 都在体外和体内成功抑制了实体瘤的生长。
FMD 传播迅速且途径广泛,应及早预防。现阶段,已有诸多研究团队通过噬菌体展示等技术筛选出多种可以与FMDV 蛋白相互作用的高效Nb。研制靶向FMDV 的Nb 为将其用于FMD 的疫苗研发奠定了基础,对保障畜牧业的健康发展,改善因病毒感染造成的经济损失具有重要意义。
6 结语
Nb 研究是一个多元化的领域。Nb 凭借其自身独特的优点成为医学方面抗体研究不可或缺的角色,同时作为强大的治疗和诊断试剂,在生物技术中得到大量应用,成为当下疾病诊断、治疗和药物研发等的主要研究对象。Nb 在生物、物理和治疗方面的高效性和多功能性一直是其研发的重要驱动力,其高度稳定性使其在细胞内表达自如,并有助于产生功能性的抗原结合体。这种强有力的功能为设计分子生物学或发育生物学创新工具带来了无数可能性,为未来科研与疾病治疗开辟了新型道路。
猜你喜欢
现代仪器与医疗(2022年4期)2022-10-08
交通医学(2022年2期)2022-06-14
健康体检与管理(2022年4期)2022-05-13
临床肺科杂志(2021年4期)2021-12-23
现代临床医学(2021年5期)2021-11-02
现代临床医学(2021年4期)2021-07-31
昆明医科大学学报(2021年1期)2021-02-07
北京大学学报(医学版)(2020年6期)2020-12-14
生物学教学(2018年10期)2018-11-29
分析化学(2018年12期)2018-01-22
