
广西4个蒜头果家系幼苗光合生理特征分析
2022-11-07 01:50黄晓露杨日升骆相华苏方贵李宝财杨卓颖梁文汇
广西林业科学 2022年4期
黄晓露,杨日升,骆相华,苏方贵,李宝财,杨卓颖,梁文汇
(1.广西壮族自治区林业科学研究院 广西特色经济林培育与利用重点实验室,广西南宁 530002;2.砦牙乡农业林业水利技术服务中心,广西河池 547603;3.凤山县核桃科研开发中心,广西河池 547600)
蒜头果(Malania oleifera)为铁青树科(Olacaceae)蒜头果属单型种植物[1],自然分布于我国云南东南部和广西西部的喀斯特山区[2-3],多生长于海拔300 ~1 200 m 石灰岩低山地区,是石漠化地区重要绿化树种,分布区较窄,自然更新能力极弱,为我国重点保护的濒危物种[4]。前人研究发现野生状态下蒜头果幼苗生长需要荫蔽,在透光度较大的条件下难以存活[3]。蒜头果种子萌发率较高,实生育苗是目前主要的育苗方式,但幼苗保存率低,幼苗抵抗病害的能力差,移栽成活率低,成为蒜头果资源保护和利用的难题[3,5-6]。
光合作用是植物通过光能合成有机物质和能量的过程,是植物体内最重要的化学反应,是影响植物生长和形态结构的重要因素[7]。光环境影响濒危植物生长和更新已经得到广泛关注。徐清等[8]研究表明,光照过强导致濒危树种毛果木莲(Manglietia ventii)幼苗的生长和发育受到抑制,且过度采伐不利于生境光环境的维持。招礼军等[9]研究表明,膝柄木(Bhesa robusta)幼树在光照充足、环境开阔的林缘生境下,生长状况显著优于弱光条件的林窗、林下生境。李文英[7]报道濒危树种水青树(Tetracentron sinense)与伴生物种相比,其光合能力和光能利用率较低,在群落竞争中处于不利地位。韩彪等[10]研究表明,适当补充环境中的CO2可提高泰山花楸(Sorbus taishanensis)的光合效率,对其生长有积极效果。广西是蒜头果主要分布区,对其区内资源分布、优株筛选和种苗繁育的研究仍处于起步阶段。本研究以广西4 个蒜头果家系为材料,分析幼苗生长情况及其光合生理特征,测定叶绿素荧光参数,有助于科学合理地开发利用蒜头果种质资源,为今后加强蒜头果种苗繁育工作和原生境种群恢复,发展桂西北蒜头果经济产业奠定基础。
1 材料与方法
1.1 材料来源及育苗方法
供试材料来源于广西河池市凤山县(FS1、FS8)、巴马县(BM1)和百色市田林县(TL3)历年种子产量较高的4个野生蒜头果家系(表1)。试验苗圃位于广西壮族自治区林业科学研究院(108°35′E,22°92′N),属亚热带季风气候,海拔120 m,年均气温21.6 ℃,年均降水量1 304.2 mm,年均相对湿度79%。

表1 种子采集地基本情况Tab.1 Basic situation of seed collection places
2020年10月,采集成熟、饱满的新鲜蒜头果果实带回试验苗圃,剥离种仁,藏于消毒过的沙床中进行催芽,每7天浇1次0.1%多菌灵,催芽期间保持沙床湿润。2021年1月左右种子开始萌发,2月上旬实生苗长出真叶,2月下旬实生幼苗长至10 cm 左右时,将其转移至荫棚下,移栽于规格16 cm × 16 cm 的黑色塑料育苗杯中,育苗基质为草炭土、椰糠、蛭石和珍珠岩混合轻基质(有机物质≥15%,腐殖酸≥8%,pH值接近中性),进行常规育苗管理。
1.2 试验方法
1.2.1 生长情况测定
2021年7月,测定幼苗的苗高、地径,每个家系测定30株,取平均值。
1.2.2 光响应曲线测定
2021年7月中旬,在晴天8:00 ~11:30 采用Li-6400 便携式光合测定仪测定净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)。选择从顶端往下第7 ~12 片无破损的成熟叶片,每个家系重复3次。有效光辐射(PAR)设定为1 800、1 500、1 200、1 000、800、600、400、200、100、80、60、40、20、10 和0 μmol·m-2·s-1(红蓝光源),空气流速设置为500 mL/min,叶温控制为35 ℃,CO2浓度控制为400 μmol/m2,叶室湿度65%。
1.2.3 叶绿素荧光测定
利用便携式荧光仪测定系统FMS-2Hansatech测定初始荧光(Fo)、最大荧光(Fm)、可变荧光(Fv)、PSⅡ潜在光化学效率(Fv/Fo)和PS Ⅱ最大光化学效率(Fv/Fm)。选择生长健康无损伤的叶片用遮光板夹住30 min,暗适应后,进行测定。
1.3 计算公式
水分利用率(WUE,μmol/mmol)的计算公式如下[11]:

潜在水分利用效率(WUEi,μmol/mmol))的计算公式如下[12]:

光能利用效率(LUE)的计算公式为[12]:

采用非直角双曲线模型拟合光响应曲线,计算公式如下[13-14]:

式中,Pn为净光合速率;φ为表观量子效率;Q为光合有效辐射;Pnmax为最大净光合速率;Rday为暗呼吸速率;k为光响应曲角。
以0 ~200 μmol·m-2·s-1有效光辐射为横坐标,对应的净光合速率为纵坐标建立线性方程,其与X轴的交点数值为光补偿点(LCP),与非直角双曲线模型计算出的Pnmax的交点在X轴上的数值为光饱和点(LSP)。
1.4 数据处理
采用SPSS v20.0 软件对试验数据进行统计处理、方差分析、多重比较和相关性分析。
2 结果与分析
2.1 生长情况
TL3、BM1、FS1 和FS8 家系幼苗苗高、地径均差异极显著(P<0.01)(图1)。其中,TL3 的苗高、地径均最大,分别为47.20 cm 和5.60 mm;FS8 的苗高、地径均最小,分别为38.00 cm 和3.93 mm,分别比TL3小19.49%和29.82%。BM1的苗高、地径为43.80 cm和5.12 mm,FS1的苗高、地径为40.57 cm和4.76 mm。
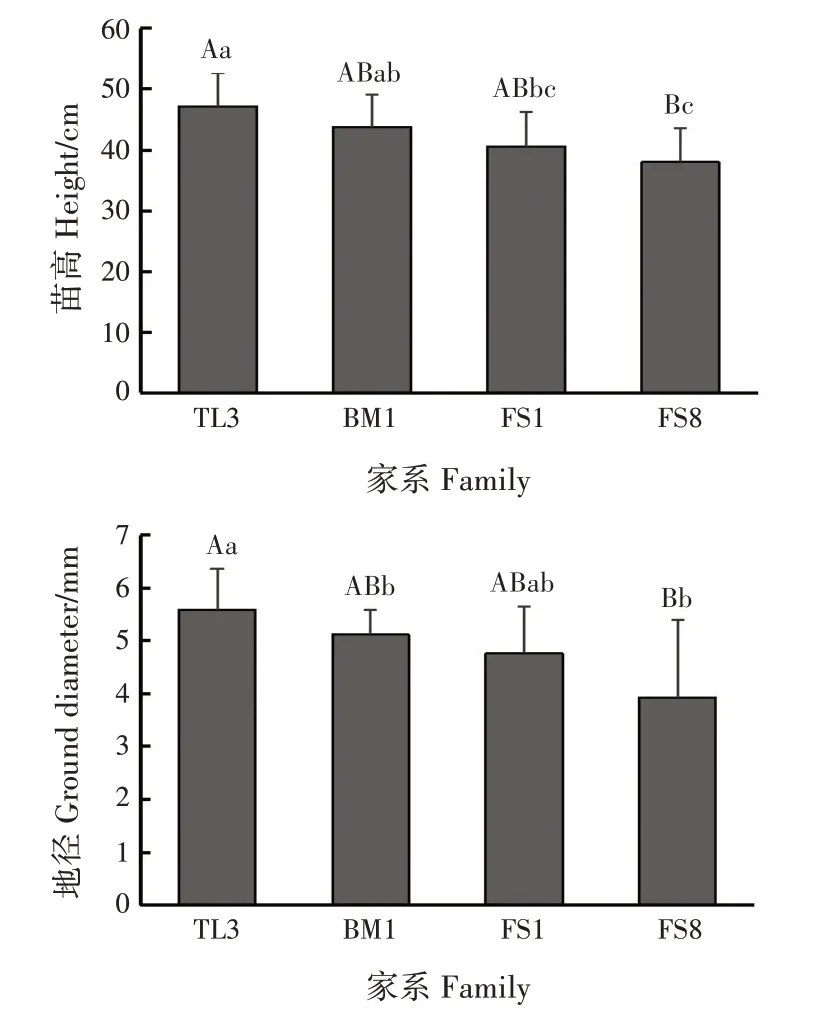
图1 幼苗生长情况Fig.1 Situation of seedling growth
2.2 光响应曲线参数
有效光辐射为0 ~600 μmol·m-2·s-1时,Pn呈增加趋势;有效光辐射为600 ~1 000 μmol·m-2·s-1时,Pn趋于平稳(图2)。有效光辐射为1 200 μmol·m-2·s-1时,FS8 幼苗叶片的Pn下降;有效光辐射为1 800 μmol·m-2·s-1时,TL3、BM1和FS1幼苗叶片的Pn均下降,出现轻微的光抑制现象。
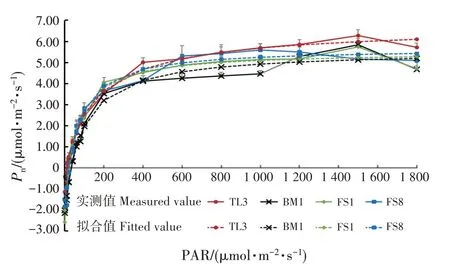
图2 光响应曲线Fig.2 Light response curves
非直角双曲线模型拟合计算结果表明,4 个家系幼苗叶片光合特征参数均差异极显著(P<0.01)(表2)。其中,TL3的Pnmax最大(7.890 μmol·m-2·s-1),其次是BM1(7.707 μmol·m-2·s-1),FS1 和FS8 的Pnmax较小,分别为7.520 和7.511 μmol·m-2·s-1;BM1 的LCP 最大(44.556 μmol·m-2·s-1),TL3 最小(6.170 μmol·m-2·s-1);TL3的LSP最大(380.266 μmol·m-2·s-1),FS1最小(292.703 μmol·m-2·s-1);BM1的Rday最大(2.142 μmol·m-2·s-1),TL3最小(1.143 μmol·m-2·s-1);TL3、FS1的AQY 最大(0.125 μmol·m-2·s-1),BM1 最小(0.093 μmol·m-2·s-1)(表2)。
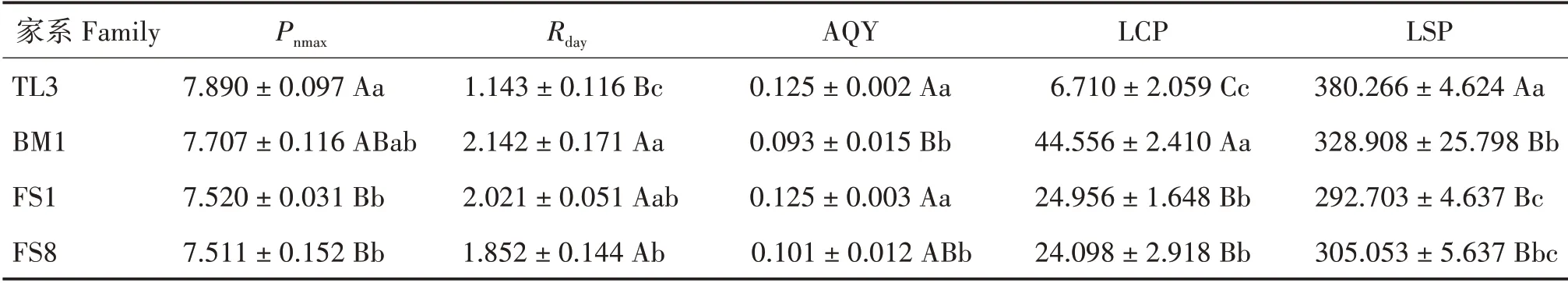
表2 光合生理参数Tab.2 Photosynthetic physiological parameters(μmol·m-2·s-1)
2.3 光合指标变化特征
4个家系幼苗叶片的Gs、Tr、WUE和WUEi均随有效光辐射增加呈上升趋势,Ci呈下降趋势,LUE 呈先升后降的趋势(图3 ~7)。TL3幼苗叶片的Gs和Ci比其他家系小;在有效光辐射为400 μmol·m-2·s-1时,WUE和WUEi最大,分别为4.32和158.55 μmol/mmol。BM1 的Gs和Ci比其他家系大,Tr、LUE 和WUEi偏小。FS1 的Tr在有效光辐射为0 ~400 和1 000 ~1 800 μmol·m-2·s-1时大于其他家系,WUE 小于其他家系。在有效光辐射60 μmol·m-2·s-1时,FS1、FS8 的LUE较大,分别为32.59×10-3和33.32×10-3。

图3 气孔导度变化特征Fig.3 Variation characteristics of Gs

图4 胞间CO2浓度变化特征Fig.4 Variation characteristics of Ci

图5 蒸腾速率浓度变化特征Fig.5 Variation characteristics of Tr

图6 光能利用效率变化特征Fig.6 Variation characteristics of LUE

图7 水分利用效率和潜在水分利用效率变化特征Fig.7 Variation characteristics of WUE and WUEi
2.4 叶绿素荧光参数
各家系幼苗叶片的Fo、Fv 和Fm均差异极显著(P <0.01),Fv/Fm和Fv/Fo差异显著(P<0.05)(表3)。FS8 的Fo最大(188.20),TL3 最小(94.50);FS8 的Fv最大(837.68),BM1 最小(507.18);FS8 的Fm最大(1 026.40),BM1最小(619.00)。TL3的Fv/Fm和Fv/Fo均最大,分别为0.85和5.64,FS8均最小,分别为0.82和4.45。

表3 叶绿素荧光参数Tab.3 Chlorophyll fluorescence parameters
2.5 相关性分析
对4 个家系幼苗生长指标、光响应参数和光饱和点附近(400 μmol·m-2·s-1)光合特征进行相关性分析。结果表明,苗高与地径、Pnmax均呈显著正相关(P<0.05),与Fo呈显著负相关(P<0.05)。地径与Fo呈极显著负相关(P<0.01),与Fv呈显著负相关(P<0.05)。Pnmax与LSP 呈显著正相关(P<0.05),LCP与Gs呈显著正相关(P<0.05),LSP与WUE呈显著正相关(P<0.05)。Fv/Fm与Pn、LUE 均呈极显著正相关(P<0.01),与WUEi呈显著正相关(P<0.05),与Ci呈显著负相关(P<0.05)。Fv/Fo与Pn、LUE 均呈显著正相关(P<0.05)。Pn与Ci呈显著负相关(P<0.05),与WUEi呈显著正相关(P<0.05),与LUE 呈极显著正相关(P<0.01)。Gs与Ci呈显著正相关(P<0.05),与WUEi呈显著负相关(P<0.05)。Ci与WUEi呈极显著负相关(P<0.01),与LUE呈显著负相关(P<0.05)。WUEi和LUE呈显著正相关(P<0.05)(表4)。

表4 光合指标与生长性状之间的相关分析Tab.4 Correlation analysis among photosynthetic parameters and growth traits
光合指标与原生地自然环境之间的相关分析表明,Pnmax与纬度呈显著负相关(P<0.05),Rday与经度、年均气温和年均降水量均呈显著正相关(P<0.05),Fv/Fm与经度、年均气温和年均降水量均呈显著负相关(P<0.05),WUEi与经度呈显著负相关(P<0.05)(表5)。

表5 光合指标与原生地自然环境之间的相关分析Tab.5 Correlation analysis among photosynthetic parameters and native natural environment
3 讨论与结论
光响应曲线是植株对生存环境适应性中光合生理信息的重要反映,是确定植株对光照强度适应范围的主要依据[15]。张光飞等[16]通过实测发现广南县蒜头果2年生实生苗的光补偿点光强为8.1 μmol·m-2·s-1,光强达750 μmol·m-2·s-1后,光合速率达到平稳,最大净光合速率接近5.5 μmol·m-2·s-1。本研究表明,广西蒜头果各家系幼苗在有效光辐射达到600 μmol·m-2·s-1后,实测的净光合速率增幅减少,并趋于平稳,与张光飞等[16]的研究结果有一些差异,可能与产地环境及育苗条件不同有关。采用非直角双曲线模型拟合的光响应曲线,可知4 个家系的最大净光合速率为7.511 ~7.890 μmol·m-2·s-1,光饱和点为292.703 ~380.266 μmol·m-2·s-1,光补偿点为6.170 ~44.556 μmol·m-2·s-1,说明蒜头果能在较低的光强下开始物质积累,且随光照强度增加较快达到最大光合效率,具有一定程度的耐荫能力。有效光辐射在1 200 ~1 800 μmol·m-2·s-1时,蒜头果各家系幼苗出现轻微的光抑制现象,与付一笑[5]研究结果相同。光抑制是植株通过调整光系统运作消散过度吸收太阳能的保护机制[17],在蒜头果幼苗培育时要适当遮阳,以免出现日灼现象,同时也对蒜头果苗木人工造林的光照控制提出要求[18]。4 个家系幼苗中,TL3幼苗的最大净光合速率最大,光补偿点最小,光饱和点最大,表明该家系幼苗光环境的适应能力较强,对光照要求较低且光合作用能力较强,而FS1 和FS8 幼苗的最大净光合速率较小,光饱和点较低,对光照要求较高,光合作用能力较弱。光合作用与植物的光生产力直接相关[19]。TL3 幼苗的最大净光合速率最大,其苗高、地径最大,其次为BM1幼苗。原生地自然环境对蒜头果幼苗的光合特性也有一定程度的影响,相关性表明原产地经度、年均气温和年均降水量越低,幼苗呼吸速率越低,水分利用潜力越高;纬度越低,净光合速率越高。
叶绿素荧光参数是有效衡量植物受害程度和光合潜能的指标[20-21],在健康生理状态下大多数高等植物的Fv/Fm值为0.80 ~0.85[22]。本研究发现蒜头果幼苗的Fv/Fm值为0.82 ~0.85,皆处于正常的生长状态。相关性分析表明Fv/Fm、Fv/Fo值越高,在饱和光强附近,幼苗的潜在水分利用率和光能利用效率越强。Fv/Fm与家系原生地自然条件关系密切,经度、年平均气温和年均降水量越低,Fv/Fm越高。4个家系幼苗中,TL3原生地的经度、年平均气温和年均
降水量均较低,其Fv/Fm、Fv/Fo值最大,电子传递活性最大,捕获的光能转化成化学能的效率更高[20]。
植物的光合作用不仅是物质和能量的累积,还能敏感反映水分和CO2利用变化的生理过程。叶片气孔导度反映植物气孔行为对光合、呼吸和蒸腾速率的调节作用,胞间CO2浓度能从侧面反映植物净光合速率和叶片CO2的利用率,蒸腾速率则表示植物的耗水情况[23]。本研究中的光合生理特征曲线表明,随光合速率增大,蒜头果幼苗叶片气孔增大,CO2消耗量增加,蒸腾速率和水分利用效率增加。4个家系幼苗光合生理特征表现与自身特性有关,在饱和光强下TL3 以较低的气孔导度水平,维持较高的光合速率、CO2消耗量,可见幼苗没有表现出气孔限制效应[24-26]。
本试验中蒜头果家系植株处于苗期阶段,其光合生理特征受育苗条件及管理措施影响较大,且由于蒜头果幼苗期和造林成年的生长环境、生长表现和适应性有较大差异,所以在后续的试验中需与蒜头果成年植株的光合特性进行对比分析,进一步研究探索蒜头果对光照的需求和响应,为科学指导蒜头果的种苗繁育与种植栽培提供理论依据。
猜你喜欢
农业工程学报(2022年4期)2022-04-24
农业与技术(2020年12期)2020-07-04
东方少年·阅读与作文(2019年2期)2019-06-11
中西医结合心血管病电子杂志(2018年15期)2018-09-12
读写算(2018年6期)2018-08-22
新课程·下旬(2017年7期)2017-08-14
考试周刊(2016年63期)2016-08-15
科技视界(2016年1期)2016-03-30
