
翼首草鲨烯合酶(PhSQS2)全长克隆、表达特性分析及蛋白功能验证
2022-11-05 03:37蒋庆锋兰小中
中草药 2022年21期
王 芸,吴 宇,蒋庆锋,何 焱,兰小中,张 磊, 2*
• 药材与资源 •
翼首草鲨烯合酶(PhSQS2)全长克隆、表达特性分析及蛋白功能验证
王 芸1,吴 宇2#,蒋庆锋3,何 焱2, 4,兰小中4*,张 磊1, 2*
1. 上海大学医学院生物医药创新研发中心,上海 200444 2. 海军军医大学药学院 药用植物学教研室,上海 200433 3. 西藏军区总医院 医疗保障中心药剂科,西藏 拉萨 850000 4. 西藏农牧学院 西南大学药用植物联合研发中心,西藏 林芝 860000
从匙叶翼首草转录组中挖掘参与三萜合成的鲨烯合酶(squalene synthase,SQS)并研究其作用机制。以翼首草基原植物匙叶翼首草为研究对象,基于前期转录组数据筛选克隆基因,并进行生物信息学分析;通过烟草瞬时转化实验观察PhSQS2亚细胞定位情况,利用实时荧光定量PCR分析PhSQS2在不同器官中的表达特征;构建原核表达载体经体外酶促反应鉴定PhSQS2功能。基因开放阅读框(open reading frame,ORF)区域全长1242 bp编码414个氨基酸,蛋白相对分子质量为47 400,理论等电点为6.57,总平均疏水性为−0.075,为不稳定蛋白。PhSQS2主要定位在细胞核和细胞质中;组织分布具有特异性,在根和叶中均有表达,叶中表达量显著高于根中。体外酶促实验证明PhSQS2催化法尼基焦磷酸生成鲨烯。PhSQS2的功能鉴定为解析翼首草中萜类物质生物合成途径,提高三萜皂苷类成分含量及优质种质资源筛选培育提供了前期理论基础。
匙叶翼首草;翼首草;鲨烯合酶(PhSQS2);生物信息学;表达特性;基因功能
中药翼首草系藏族习用药材,为川续断科植物匙叶翼首草(C. B. Clarke) Höeck的干燥全草[1],具有解毒除瘟、清热止痢、祛风通痹等功效。作为藏医药常用药材之一,是二十五味余甘子丸、十二味翼首散等中药的组方药,多用于瘟病时疫、痢疾、热病发烧、风湿性关节炎等疾病的治疗[2]。富含皂苷类成分是川续断科植物的一大特点,化学成分研究发现翼首草活性成分主要包括五环三萜类皂苷、环烯醚萜苷等萜类成分[3],目前已分离出齐墩果酸、熊果苷酸、匙叶翼首草苷A~D等五环三萜类化合物[4-5],马钱苷等环烯醚萜苷类化合物[6-7]。其中三萜总皂苷具有抗肿瘤、护肝、抗炎、调节免疫等药理活性[8],环烯醚萜苷化合物的药理活性研究发现其具有抗氧化、抗病毒等作用[9]。
萜类化合物是植物体内广泛存在的重要次生代谢产物,由异戊二烯结构单元构成基本骨架,主要通过异戊二烯首尾相接以及环化2种方式合成。植物胞质中的甲羟戊酸(mevalonate,MVA)和质体中的甲基赤藓醇-4-磷酸(methylerythritol- 4-phosphate,DXP/MEP)2种生物合成途径[10]催化生成萜类物质的基本单元异戊烯焦磷酸(isopentenyl diphosphate,IPP)及其同分异构体二甲基烯丙基焦磷酸酯(dimethylallyl diphosphate,DMAPP)。IPP和DMAPP在异戊烯基转移酶作用下合成萜类前体异戊二烯焦磷酸链,包括单萜前体香叶基焦磷酸(geranyl pyrophosphate,GPP)、倍半萜前体法尼基焦磷酸(farnesyl pyrophosphate,FPP)和二萜前体香叶基香叶基焦磷酸(geranylgeranyl pyrophosphate,GGPP);其中2个FPP在鲨烯合酶(squalene synthase,SQS)作用下缩合生成三萜前体鲨烯。单萜、二萜、三萜等前体在萜类合酶的作用下生成各种萜类骨架,进一步经氧化、酰基化及糖基化等化学修饰最后形成一系列复杂萜类化合物[11]。目前已从黄花蒿[12]、刺五加[13]、三七[14]、丹参[15]、白花丹参[16]、雷公藤[17]、黄芪[18]等多种药用植物中成功筛选克隆基因。三七中鲨烯合酶基因表达量同三萜总皂苷含量进行共分析发现两者存在正相关关系[14],提高刺五加中鲨烯合酶基因表达量可以促进总皂苷的积累[13],而体外重组蛋白酶促反应直接证明黄花蒿鲨烯合酶可以催化FPP转化生成鲨烯[12],由此看出鲨烯合酶是决定着法尼基焦磷酸流向的关键酶,是开启植物三萜皂苷类生物合成途径的重要开关。迄今,有关翼首草三萜皂苷类物质生物合成途径解析的研究尚未有所报道,因此发现并证明翼首草中鲨烯合酶功能对于解析五环三萜皂苷类物质的生物合成途径(图1),提高三萜皂苷类等活性成分含量具有重要意义。

图1 植物五环三萜生物合成途径[19-20]
课题组前期从翼首草转录组数据中挖掘到1条鲨烯合酶基因[21],通过并对其进行克隆、生物信息学分析、表达模式分析、蛋白诱导表达及基因功能验证等研究,证明PhSQS2具有鲨烯合酶的功能,为翼首草三萜皂苷类活性成分生物合成途径解析及提高三萜总皂苷类物质的含量奠定了基础。
1 材料
匙叶翼首草完整植株采于西藏自治区林芝市巴宜区,经西藏农牧学院兰小中教授鉴定为川续断科植物匙叶翼首草(C. B. Clarke) Höeck。样品洗净后分别收集根和叶,于液氮速冻后保存至−80 ℃冰箱。
2 方法
2.1 翼首草总RNA提取及cDNA合成
采用TransZol Up(全式金公司,中国)法提取翼首草样品RNA,经1%琼脂糖凝胶电泳和NanoDrop 2000c超微量分光光度计测定RNA纯度和浓度。而后依次使用TransScript First-Strand cDNA Synthesis SuperMix(全式金公司,中国)和Advantage 2 PCR Kit(Clonetech公司,美国)完成双链cDNA扩增,最终构建翼首草cDNA文库,于−80 ℃冰箱保存备用。
2.2 PhSQS2基因序列数据挖掘及克隆
从翼首草转录组数据库中获得基因序列全长[21],通过Primer在线软件设计扩增引物,引物序列见表1。翼首草RNA反转录后获得的cDNA作为模板,进行普通PCR扩增完成基因克隆。
表1 引物序列
Table 1 Primers sequences
引物名称序列(5′→3′)用途 PhSQS2-FATGGGAGGTATGGGAGAAATTTTGA克隆 PhSQS2-RGAAAGCTAACCATGTGTTATGTGAT克隆 PhSQS2-Q-FTTGGCCTCGTGAAATTTGGGqRT-PCR PhSQS2-Q-RCCAATTGCCATGACCTGAGGqRT-PCR PhSQS2-sub-FGACGAGCTGTACAAGGGATCCATGGGAGGTATGGGAGAAATTTTGA亚细胞定位 PhSQS2-sub-RCTAGAGGATCAATTCGAGCTCGAAAGCTAACCATGTGTTATGTGAT亚细胞定位 PhSQS2-pET-FGGATCCGAATTCGAGCTCATGGGAGGTATGGGAGAAATTTTGA原核表达 PhSQS2-pET-RGAGTGCGGCCGCAAGCTTGAAAGCTAACCATGTGTTATGTGAT原核表达
2.3 PhSQS2生物信息学分析
用ProtParam(http://web.expasy.org/compute)在线软件分析PhSQS2蛋白基本理化性质;通过ProtScale(https://web.expasy.org/protscale)在线软件对PhSQS2进行亲疏水性预测;利用Pfam32.0 (http://pfam.xfam.org/search)和NCBI数据库BLAST (CD-Search)(https://www.ncbi.nlm.nih.gov/ Structure/cdd/wrpsb.cgi)在线软件对PhSQS2蛋白进行功能域的预测;依赖Swiss-Model(http:// www.expasy.ch/swissmod/SWISS-MODEL. html)在线软件进行蛋白的三级结构预测,并用PyMOL实现PhSQS2三维结构可视化。
2.4 PhSQS2实时荧光定量PCR分析
选取翼首草基因作为内参基因[22],使用NovoScript®SYBR One-Step qRT-PCR Kit(近岸公司,上海)试剂盒进行qRT-PCR,使用2−ΔΔCt法进行处理,实验设置3组生物学重复,数据使用平均值和标准差,并且相对定量时将目的基因在根中的表达量设为“1”。
2.5 PhSQS2亚细胞定位载体构建
选择H I和I作为酶切位点设计引物构建- pCAMBIA1301-YFP亚细胞定位载体,测序正确后将重组定位载体转化农杆菌用作后续亚细胞定位实验。p-YFP载体经相同处理转化农杆菌作为阴性对照。
2.6 PhSQS2亚细胞定位分析
取200 µL已转入目的基因的农杆菌加入5 mL YEB(50 mg/L Rif+50 mg/L Kan)液体培养基中,28 ℃,220 r/min,振荡培养16 h;5000 r/min,离心10 min,去上清。MS溶液重悬菌体,待菌液值约为0.6时加入终浓度为10 mmol/L吗啉乙磺酸和100 µmol/L乙酰丁香酮,室温静置3 h;缓慢匀速注射于本氏烟草叶片背面,暗培养植株2~3 d后通过激光共聚焦显微镜观察结果。
2.7 PhSQS2原核表达载体构建及转化
选择I和d III作为酶切位点设计引物构建PhSQS2-pET-32a+原核表达载体,测序正确后将重组表达载体转化BL21(DE3)感受态细胞用作后续蛋白表达。pET-32a+载体经相同处理转化BL21(DE3)感受态细胞后作为阴性对照。
2.8 PhSQS2重组蛋白诱导表达
取含有PhSQS2-pET-32a+和pET-32a+阳性克隆的菌液各10 µL加入1 mL含有LB(100 mg/L Amp)液体培养基中,37 ℃振荡培养2 h。取500 µL活化后菌液加入1000 mL 乳酸杆菌(100 mg/L 磷酸腺苷)液体培养基培养到600为0.6加入终浓度为1 mmol/L的IPTG,80 r/min,16 ℃过夜培养;4 ℃离心收集菌体,5 mL 1×PBS缓冲液重悬菌体;冰上超声(功率25%,超声5 s,间隔5 s)破碎20 min;4 ℃离心收集上清液;使用SDS-PAGE电泳检测上清液中是否存在目的蛋白表达。
2.9 PhSQS2体外酶促实验及产物检测
使用Bio-ScaleTM Mini ProfinityTM IMAC Cartridges纯化柱进行蛋白纯化,根据重组蛋白内含有His标签与相应抗体特异性结合的原理经Western blotting实验检测是否表达最终目的蛋白,使用Bradford Protein Assay Kit(碧云天)测定蛋白浓度。
PhSQS2催化反应体系(300 µL):50 µmol/LFPP;20 mmol/LMgCl2;1 mmol/LDTT;2% Glycine;3 mmol/LNADPH;5 μg纯化蛋白;50 mmol/LTris-HCl补齐。30 ℃,300 r/min反应过夜;结束后加入400 µL正己烷淬灭反应,400 µL正己烷萃取3次,合并上层有机相氮吹挥干浓缩,200 µL正己烷复溶,进行GC-MS检测。
3 结果与分析
3.1 PhSQS2克隆及序列分析
以翼首草cDNA文库为模板进行PCR扩增,产物经1%琼脂糖凝胶电泳检测,结果显示在约1200 bp处出现一特异片段(图2),与基因全长预期结果一致。

M-Marker 1-PhSQS2基因克隆条带
克隆结果测序后证实与翼首草转录组数据中所获序列全长完全相同。利用开放阅读框(open reading frame,ORF)Finder(https://www.ncbi. nlm.nih.gov/orffinder)在线软件对序列进行信息分析,发现基因ORF序列全长1242 bp,编码414个氨基酸。利用DNAMAN软件将其与其他相似度较高的植物氨基酸序列进行多重比对(图3),分析结果显示PhSQS2氨基酸序列与丹参Bunge、黄花蒿L.相似度均达到80%以上。

图3 不同物种SQS氨基酸序列多重比对
3.2 PhSQS2生物信息学分析
基因ORF区域编码414个氨基酸,蛋白相对分子质量为47 400,理论等电点为6.57,蛋白总平均疏水性为−0.075,属于不稳定蛋白。采用ProtScale在线分析软件对翼首草PhSQS2亲水性/疏水性进行预测与分析,其中PhSQS2多肽链第248位氨基酸分值最低,亲水性最强;第398位氨基酸分值最高,疏水性最强,结果同时可以看出蛋白疏水区域大于亲水区域,故PhSQS2属于疏水蛋白(图4-A)。
利用Pfam 32.0及BLAST(CD-Search)在线软件预测分析翼首草PhSQS2蛋白构域,结果显示,PhSQS2含有关键的SQS_PSY功能域和PLN02632保守域(图4-C),采用SWISS-MODEL在线软件以TcSQS(PDB:3wca)作为模板经同源建模预测PhSQS2蛋白三级结构,经PyMOL可视化后结果见图4-B。
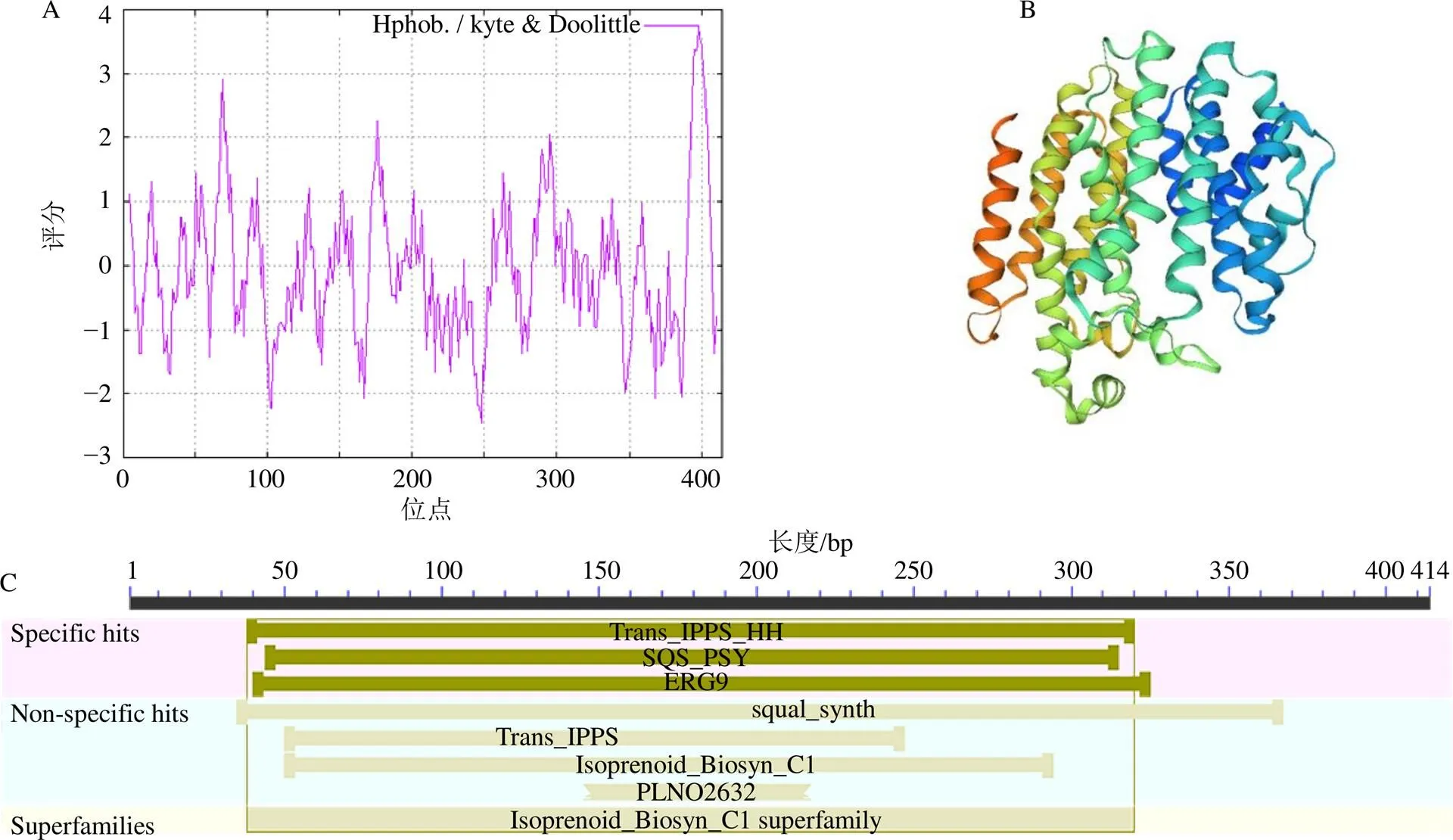
图4 翼首草SQS2蛋白亲水性/疏水性(A)、蛋白三级结构预测(B)、蛋白保守结构域(C)
3.3 PhSQS2组织表达特异性
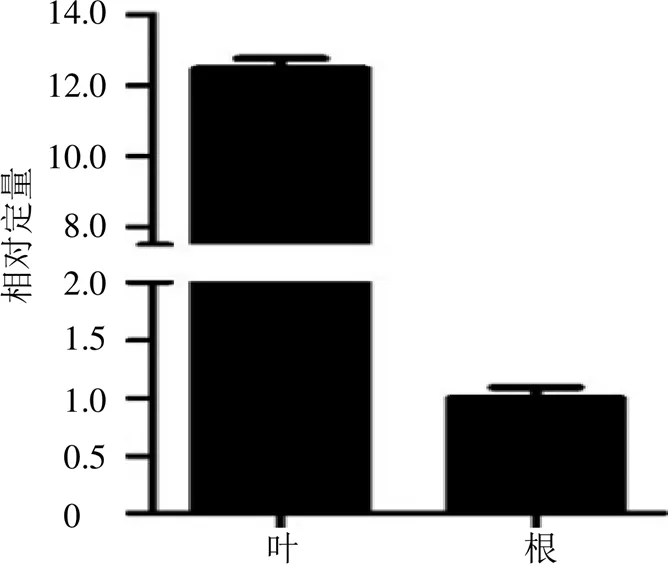
使用qRT-PCR检测在翼首草不同组织(根、茎)中的分布情况。结果(图5)显示在根、叶中的表达量具有显著差异,叶中的表达量约为根中的12.5倍。
3.4 PhSQS2亚细胞定位特征
构建PhSQS2-YFP融合表达载体并转染本氏烟草,通过激光共聚焦显微镜扫描YFP黄色荧光信号(514 nm)观察PhSQS2在烟草中亚细胞定位结果。结果显示YFP对照组信号(图6-A~C)分布在细胞核和细胞质;PhSQS2-YFP融合蛋白荧光信号(图6-D~F)在细胞核和细胞质中均有存在,据此判断PhSQS2-YFP融合蛋白定位在细胞核和细胞质中。
3.5 PhSQS2重组蛋白诱导表达
经IPTG诱导BL21(DE3)菌株表达后由SDS-PAGE检测重组蛋白表达情况。如图7-A所示,含有重组质粒PhSQS2-pET-32a+的菌株(PhSQS2目的蛋白相对分子质量约为47 400)在65 000附近有蛋白条带;而pET-32a+空载体对照组菌株在相同位置无蛋白表达,仅在18 000附近表达His-Tag标签蛋白,以上结果表明PhSQS2在大肠杆菌BL21(DE3)中成功表达。
**P<0.01

图6 PhSQS2在烟草表皮中的亚细胞定位

1-Marker 2、4-pET-32a+空载 3、5-PhSQS2重组蛋白
进一步利用Western blotting检测所得蛋白是否为目的蛋白。结果如图7-B所示,与空载体对照组相比,检测到PhSQS2-pET-32a+表达信号,且条带单一、纯度良好。采用Bradford准曲线法对PhSQS2- pET-32a+融合蛋白进行浓度测定,最后获得质量浓度为3.60 mg/mL的PhSQS2重组蛋白用作后续酶活实验。
3.6 PhSQS2体外酶促产物检测
对纯化后的PhSQS2酶促反应产物进行GC-MS检测,结果见图8。以pET32a+空载表达的未纯化蛋白作为阴性对照组,鲨烯对照品作为阳性对照,气相色谱结果显示PhSQS2酶促反应产物和鲨烯对照品在保留时间9.87 min处出现相同的色谱峰,对该色谱峰进行质谱分析,特征离子一致,说明PhSQS2酶促反应生成了鲨烯,PhSQS2具有鲨烯合酶的功能。
4 讨论
翼首草作为常用传统藏药材,在西藏民间具有悠久的用药史,其需求逐年增长,但因基原植物匙叶翼首草主要生长在西藏、云南、四川等海拔3000 m以上的草地、林间、林缘、高山草甸[23],严苛的生长环境制约了其后续应用,目前药材来源主要以野生采挖为主,长期野生采挖使翼首草的野生资源濒临灭绝[24]。近些年对于翼首草的研究主要集中在基原植物匙叶翼首草的人工栽培[25],翼首草中活性成分生物合成途径解析、活性成分调控,以及优良种质资源的培育等方向。生物合成途径中的关键酶活性影响活性成分的产量,因此调控植物中关键酶基因的表达是提高活性成分产量、改善中药材品质的重要手段。对于翼首草萜类物质途径解析,本研究主要聚焦于SQS的克隆以及相关功能研究。体外蛋白功能验证PhSQS2具有鲨烯合酶的功能,可催化鲨烯生成。后续构建PhSQS2过表达载体或通过激素诱导提高植物体内基因表达量,使法尼基焦磷酸更多地流向三萜类物质合成,将有利于三萜皂苷类成分的积累。组织特异性表达结果表明PhSQS2在根和叶中均有分布,叶中表达量显著高于根中。但已有报道的翼首草三萜总皂苷代谢数据与上述转录数据并不一致,比较翼首草不同部位的总皂苷含量,发现根和叶中均含有丰富的总皂苷类物质,但根中含量要高于叶中[26-27]。类似的结果在三七中也被检测到,SQS在根中的表达量要高于芦头和茎中,但芦头中总皂苷含量最高[14]。分析造成转录代谢分布不一致的原因,认为可能是因为SQS位于三萜合成途径上游,下游其他关键酶如环氧酶、环化酶、单加氧酶等在翼首草叶中的表达影响了皂苷的合成;或是三萜合成途径中存在着中间体的定向转运影响了终产物的分布。但可以明确的是提高鲨烯合酶的表达量可以提高植物中三萜皂苷的含量[13],鲨烯合酶在三萜生物合成途径中具有重要作用。挖掘到翼首草中的鲨烯合酶对于提高三萜皂苷类物质含量以及筛选培育翼首草优质种质资源具有重要意义。

A-鲨烯对照品 B-PhSQS2 C-pET-32a+ D-峰1质谱图 E-峰2质谱图
利益冲突 所有作者均声明不存在利益冲突
[1] 中国药典[S]. 四部. 2020: 23.
[2] 中国科学院西北高原生物研究所. 藏药志 [M]. 西宁: 青海人民出版社, 2019: 36.
[3] 郭晨旭, 朱国福. 藏药翼首草化学成分及药理作用研究进展 [J]. 世界中医药, 2015, 10(9): 1440-1443.
[4] 权红, 甄梓娟, 李连强, 等. 藏药匙叶翼首草中齐墩果酸及熊果酸的含量测定 [J]. 中国现代中药, 2016, 18(6): 762-765.
[5] Zeng Y B, Mei W L, Zhao Y X,. Two new epimeric pairs of iridoid from mangrove plant[J]., 2007, 18(12): 1509-1511.
[6] 田军, 吴凤锷, 丘明华, 聂瑞麟. 匙叶翼首花的化学成分 [J]. 天然产物研究与开发, 2000, 12(1): 35-38.
[7] Graikou K, Aligiannis N, Chinou I B,. Cantleyoside-dimethyl-acetal and other iridoid glucosides from: Antimicrobial activities [J]., 2002, 57(1/2): 95-99.
[8] Zhang L, Hu J J, Lin J W,. Anti-inflammatory and analgesic effects of ethanol and aqueous extracts of(C. B. Clarke) Höeck [J]., 2009, 123(3): 510-514.
[9] 张雪梅, 杨丰庆, 夏之宁. 藏药翼首草的药理作用及其质量评价研究进展 [J]. 中国药房, 2012, 23(35): 3356-3358.
[10] Pu X J, Dong X M, Li Q,. An update on the function and regulation of methylerythritol phosphate and mevalonate pathways and their evolutionary dynamics [J]., 2021, 63(7): 1211-1226.
[11] Singh B, Sharma R A. Plant terpenes: Defense responses, phylogenetic analysis, regulation and clinical applications [J]., 2015, 5(2): 129-151.
[12] 李振秋, 王花红, 王红, 等. 中药青蒿鲨烯合酶的大肠杆菌表达、纯化与功能鉴定 [J]. 应用与环境生物学报, 2007, 13(3): 309-312.
[13] 邢朝斌, 龙月红, 劳凤云, 等. 刺五加鲨烯合酶基因的表达及其对皂苷含量的影响 [J]. 经济林研究, 2013, 31(1): 25-29.
[14] 吴耀生, 朱华, 李珅, 等. 三七鲨烯合酶基因在三七根、茎、芦头中的转录表达与三萜皂苷合成 [J]. 中国生物化学与分子生物学报, 2007, 23(12): 1000-1005.
[15] 马艺沔, 袁丽钗, 张林甦, 等. 2个丹参鲨烯合酶基因的克隆和鉴定 [J]. 中草药, 2014, 45(9): 1307-1312.
[16] 荣齐仙, 姜丹, 查良平, 等. 白花丹参鲨烯合酶SQS2的克隆与原核表达分析 [J]. 中国中药杂志, 2015, 40(7): 1259-1265.
[17] 刘雨佳, 苏平, 王秀娟, 等. 雷公藤鲨烯合酶基因全长cDNA克隆及诱导表达分析 [J]. 药学学报, 2016, 51(4): 657-661.
[18] 李振秋, 王晓明, 金亚明, 等. 黄芪鲨烯合酶基因的克隆和序列分析 [J]. 河北林果研究, 2011, 26(1): 16-19.
[19] Misra R C, Maiti P, Chanotiya C S,. Methyl jasmonate-elicited transcriptional responses and pentacyclic triterpene biosynthesis in sweet basil [J]., 2014, 164(2): 1028-1044.
[20] Unland K, Pütter K M, Vorwerk K,. Functional characterization of squalene synthase and squalene epoxidase in[J]., 2018, 2(6): e00063.
[21] 何焱, 李忠玥, 李卿, 等. 藏药翼首草角鲨烯合成酶的生物信息学分析 [J]. 基因组学与应用生物学, 2020, 39(7): 3139-3150.
[22] 何焱, 李忠玥, 刘江, 等. 翼首草实时荧光定量PCR内参基因筛选 [J]. 分子植物育种, 2020, 18(9): 2987-2993.
[23] 关昕璐, 阎玉凝, 任子和, 等. 翼首草的鉴别研究 [J]. 中国中药杂志, 2004, 29(11): 1027-1030.
[24] 甄梓娟, 徐元江, 廖志华, 等. 藏药匙叶翼首草及其同属植物的研究进展 [J]. 中药材, 2016, 39(1): 223-228.
[25] 兰小中, 周战. 一种高海拔地区翼首草的高产人工栽培技术: CN103141291A [P]. 2013-06-12.
[26] 林升得, 江道峰, 张艺, 等. 分光光度法测定藏药翼首草不同药用部位总皂苷的含量 [J]. 中国实验方剂学杂志, 2013, 19(5): 101-104.
[27] 杨荣平, 向春艳, 张小梅, 等. 不同产地翼首草中总皂苷的含量比较 [J]. 时珍国医国药, 2010, 21(7): 1797-1798.
Cloning, expression pattern and functional analysis of squalene synthase (PhSQS2) in
WANG Yun1, WU Yu2, JIANG Qing-feng3, HE Yan2, 4, LAN Xiao-zhong4, ZHANG Lei1, 2
1. Biomedical Innovation R&D Center, School of Medicine, Shanghai University, Shanghai 200444, China 2. Department of Pharmaceutical Botany, School of Pharmacy, Second Military Medical University, Shanghai 200433, China 3. Pharmacy Department, Medical Security Center, General Hospital of Tibet Military Region, Lhasa 850000, China 4.Animal Husbandry College and TAAHC-SWU Medicinal Plant R&D Centre, Tibet Agricultural and Animal Husbandry University, Nyingchi 860000, China
To explore the squalene synthase (SQS) from the transcriptome ofand investigate its mechanism of action.Takingas research object , thegene was cloned and bioinformatically analyzed based on preliminary transcriptome data; The subcellular localization of PhSQS2 was observed by transient transformation experiments in tobacco, and the expression characteristics of PhSQS2 in different organs were analyzed by real-time fluorescence quantitative PCR; Finally, the prokaryotic expression vector was constructed to identify the function of PhSQS2 by enzymatic reaction.Thegene was screened with the length of open reading frames 1242 bp, encoding proteins 414 amino acids. Bioinformatics analysis showed that it was an unstable hydrophobic protein with a relative molecular weight of 47 400, a theoretical PI of 6.57 and an average hydrophobicity of −0.075. PhSQS2 was mainly localized in nucleus and cytoplasm;Its tissue distribution was specific,and it was expressed in both roots and leaves, and the expression in leaves were significantly higher than those in roots.enzymatic reactions proved that as a squalene synthase, PhSQS2 catalysed the production of squalene from farnesyl pyrophosphate.The functional identification of PhSQS2 provides a theoretical basis for the further analysis of the terpenoid biosynthesis pathway and the improvement of triterpenoid saponins from, as well as for in-depth research on the screening and cultivation of high-quality germplasm resources.
(C.B.Clarke) Höeck;; squalene synthase (PhSQS2); bioinformatics analysis; expression pattern; gene function
R286.12
A
0253 - 2670(2022)21 - 6840 - 08
10.7501/j.issn.0253-2670.2022.21.022
2022-04-03
国家自然科学基金资助项目(31970316);上海市优秀学术带头人计划(19XD1405000)
王 芸(1993—),女,助理实验师。E-mail: ssalvia4444@shu.edu.cn
兰小中(1973—),男,教授,研究方向为藏药资源、中药资源与品质调控。E-mail: lanxiaozhong@163.com
张 磊(1977—),男,教授,研究方向为中药资源与品质调控。E-mail: starzhanglei@aliyun.com
#共同第一作者:吴 宇(1988—),女,助教,研究方向为重要活性天然产物的生物合成与代谢调控。E-mail: wuyuagnes@163.com
[责任编辑 时圣明]
猜你喜欢
中医药导报(2022年6期)2022-11-07
湖北农业科学(2022年16期)2022-09-29
环球时报(2022-09-20)2022-09-20
中国现代医生(2022年21期)2022-08-22
昆明医科大学学报(2022年1期)2022-02-28
广东教育·职教版(2021年3期)2021-04-20
学校教育研究(2018年27期)2018-05-14
小资CHIC!ELEGANCE(2015年14期)2015-09-23
小资CHIC!ELEGANCE(2015年15期)2015-09-01
时代英语·高二(2015年2期)2015-05-18
