
红花SPL基因家族全基因组鉴定及表达分析
2022-10-28 01:21宋梦瑶董诚明朱畇昊
江苏农业科学 2022年19期
宋梦瑶, 赵 乐,2, 董诚明,2, 朱畇昊,2
(1.河南中医药大学药学院,河南郑州 450046; 2.呼吸疾病诊疗与新药研发河南省协同创新中心,河南郑州 450046)
转录因子是一类参与转录调控的蛋白,通过与DNA分子特异性结合激活或抑制靶基因的转录活性,从而使目的基因在特定的时间、空间转录及表达。SPL(SQUAMOSA promoter-binding protein-like)类转录因子为植物所特有,能识别并结合-基因启动子的SQUAMOSA区域。基因家族所编码的蛋白质序列具有高度保守的SBP结构域,该结构域大约由78个氨基酸残基组成,包含2个锌指位点(Zinc finger motif,Zn-1/Zn-2)和1个核定位信号(nuclear localization signal,简称NLS)。目前,越来越多的基因家族研究被相继报道,如拟南芥、水稻、丹参等。研究表明,基因参与植物生长和发育的调节,包括植物花和植物表皮毛的发育、植物胁迫反应和次生代谢物的合成等。
红花(L.)为菊科红花属植物,其干燥的管状花序入药称为红花,广泛应用于中医临床,具有活血通经、祛瘀止痛的功效。红花的籽粒中富含多不饱和脂肪酸——亚油酸和油酸,在食品及中医药有广泛应用。红花中分离出的羟基红花黄色素A,是其特有的黄酮类化合物,对心血管类疾病具有显著的治疗效果。近年来,随着红花高质量参考基因组的公布,利用基因组和生物信息学手段深入挖掘红花生长发育、亚油酸和黄酮的调控机制、抵抗生物及非生物胁迫基因,将成为红花功能基因组学研究的一个重要方向。基因家族在植物生长发育调控网络中起着关键作用,对其进行分析具有重要意义。本研究通过对红花基因家族成员的鉴定以及系统进化分析,结合其在不同组织器官中的表达模式以及在茉莉酸甲酯(methyl jasmonate,简称MeJA)和2,4-表油菜素内酯(2,4-epibrassinolide,简称EBR)诱导过程中的表达差异分析,以期深入解析该基因家族的作用机制,为红花功能基因组的研究提供候选基因资源。
1 材料与方法
1.1 红花CtSPL基因家族成员鉴定及蛋白特征分析
从红花基因组数据库(http://safflower.scuec.edu.cn)下载红花基因组、蛋白序列和注释文件,从PFAM数据库下载SBP结构域的HMMER文件,从红花基因组中提取具有SBP结构域的蛋白序列。利用 NCBI CDD在线预测 SBP蛋白结构域,除去无典型结构域、结构不完整和冗余序列。进而利用ProtParam (http://us.expasy.org/tools/protparam.html)进行红花SPL蛋白理化性质分析,利用SOPMA (https://prabi.ibcp.fr/htm/site/web/home)预测二级结构,用WoLF PSORT (https://wolfpsort.hgc.jp/)分析亚细胞定位。
1.2 红花CtSPL基因家族结构域和系统进化分析
使用DNAMAN9.0软件对红花SBP保守结构域进行多序列比对分析,并将21个红花CtSPL蛋白序列和下载于TAIR数据库(https://www.arabidopsis.org/)的拟南芥AtSPL蛋白序列以及下载于PlantTFDB数据库(http://planttfdb.gao-lab.org/)的毛果杨PtSPL蛋白序列用于系统进化分析。使用MEGA7采用邻近法构建系统发育树,Bootstrap设置为10000,其余参数为默认值。使用Evolview (https://www.evolgenius.info/evolview/)在线软件进行后期美化。
1.3 红花CtSPL基因家族染色体定位、基因结构和保守基序分析
运用MapChart2.32软件根据红花基因组注释文件绘制红花基因的染色体定位图。从基因组注释文件中获取红花的基因结构注释信息,同时通过MEME在线软件(http://meme-suite.org/tools/meme)分析家族成员的保守序列。最后用TBtools软件将结果可视化。
1.4 红花CtSPL基因家族顺式作用元件预测和共线性分析
利用TBTools软件,从红花基因组数据库中提取红花基因起始密码子上游1.5 kb作为启动子区域,将所有红花基因的启动子序列提交到PlantCARE数据库(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)预测顺式作用元件。使用 MCScanX软件对红花基因组内的基因以及红花与拟南芥、向日葵基因组之间的基因进行共线性分析,用Circos 软件(http://circos.ca)绘制基因组内和基因组间的共线性图谱。
1.5 红花CtSPL基因家族miRNA靶位点预测
在PmiREN数据库(https://www.pmiren.com/)中下载红花全部的miRNA成熟序列,使用psRNATarget (http://plantgrn.noble.org/)对红花基因编码区的miRNA结合位点进行预测,利用Cytoscape软件将预测结果可视化。
1.6 红花CtSPL基因家族表达模式分析
从NCBI数据库(https://www.ncbi.nlm.nih.gov)分别下载红花3个组织:种子、叶、花瓣(PRJNA76135)上传于2011年10月29日以及茉莉酸甲酯处理(PRJNA561476) 上传于2019年8月22日、表油菜素内酯处理(PRJNA628030)上传于2020年4月25日的高通量测序原始数据,利用NCBI SRA Toolkit将测序数据sra文件转换为fastq文件,利用转录组定量工具Kallisto计算TPM值,将其作为基因表达量。
2 结果与分析
2.1 红花CtSPL基因家族成员鉴定及蛋白特性分析
根据红花基因在11条染色体上的位置分布,将其分别命名为~。对21个CtSPL蛋白进行理化性质分析(表1),结果表明CtSPL蛋白序列长度介于147~1 078 aa之间;相对分子量范围为16.58~118.74 ku;等电点为 6.43~10.74,其中有19个CtSPL蛋白的等电点大于7,为典型的碱性蛋白;亚细胞定位结果显示,CtSPL蛋白大多定位于细胞核;二级结构预测结果表明,CtSPL蛋白组成结构相似,以无规则卷曲、α螺旋为主要组成部分。
2.2 红花CtSPL基因家族结构域和系统进化分析
使用DNAMAN软件对21个CtSPL蛋白的保守结构域进行多序列比对分析,结果见图1,大部分CtSPL蛋白均具有完整且保守的SBP结构域,该结构域一般具有2个锌指结构(Zn-1和Zn-2)和1个核定位信号(NLS),N端Zn-1结构除CtSPL20蛋白外其他成员均为 Cys-Cys-Cys-His (C3H)型,CtSPL20蛋白Zn-1结构为Cys-Cys-Cys-Cys (C4)型;C 端Zn-2结构为 Cys-Cys-His-Cys (C2HC)型,CtSPL1、CtSPL8、CtSPL10、CtSPL19、CtSPL21蛋白Zn-2结构有部分缺失或突变;核定位信号与Zn-2结构有4个氨基酸残基重合,CtSPL1、CtSPL16蛋白核定位信号末端变异。
为了解红花基因家族各成员的进化关系,选取红花、拟南芥、毛果杨基因家族各蛋白序列进行聚类分析,如图2所示,62个SPL蛋白可分为6个亚家族。Ⅰ亚家族包含的SPL成员最多,为21个,其中CtSPL蛋白8个,其次是Ⅳ亚家族共有SPL成员12个,CtSPL蛋白2个,Ⅴ亚家族共有SPL成员9个,CtSPL蛋白3个,Ⅱ亚家族共有SPL成员8个,CtSPL蛋白2个, Ⅲ亚家族共有SPL成员8个,CtSPL蛋白5个,Ⅵ亚家族共有SPL成员4个,CtSPL蛋白1个。
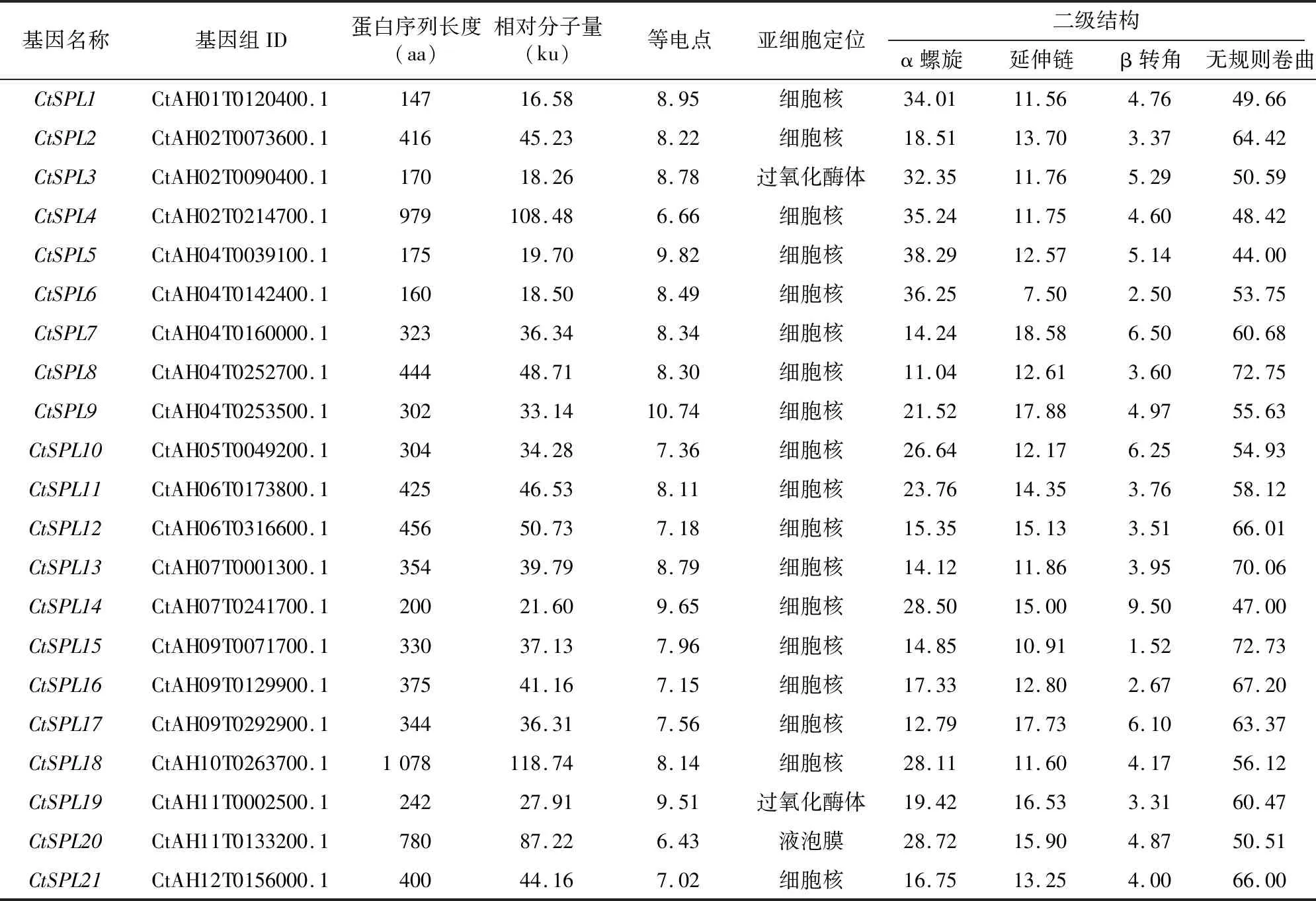
表1 红花CtSPL基因家族蛋白的理化性质
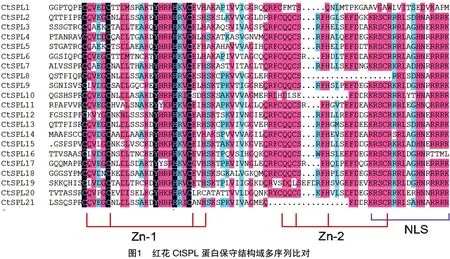
2.3 红花CtSPL基因家族染色体定位、保守基序和基因结构
根据红花基因组信息,绘制基因在红花染色体上的位置信息,由图3可知,21个基因不均匀分布在11条染色体上,其中4号染色体上基因数目最多,为5个;其次是2、9号染色体均有3个;6、7、11号染色体均有2个;而1、5、10、12号染色体只有1个。

TBtools可视化结果显示,亲缘关系较为相近的成员具有相似的保守基序和基因结构(图4)。基因家族的保守SBP结构域含有2个锌指结构、1个核定位信号,除CtSPL1、CtSPL8、CtSPL10、CtSPL16、CtSPL21外,其余成员均含有完整的SBP结构域motif1 (Zn-1)、motif2 (Zn-2)、motif3 (NLS)且位置较为固定。红花SPL蛋白还存在其他基序,如motif4、motif8、motif10仅存在于I亚家族,motif5仅存在于Ⅳ、Ⅴ亚家族,motif9仅存在于Ⅳ亚家族,motif6存在于Ⅰ亚家族的CtSPL2、CtSPL8、CtSPL10、CtSPL13、CtSPL17、CtSPL21,Ⅲ亚家族的CtSPL7以及V亚家族CtSPL11,motif7存在于Ⅱ、Ⅴ亚家族以及Ⅰ亚家族的CtSPL2、CtSPL8、CtSPL12、CtSPL13、CtSPL17、CtSPL21。
基因结构图显示,所有基因都至少含有2个外显子, 外显子数目分布在2~12个之间,内含子数目分布在1~12个之间。不同亚家族所含的外显子数量差异较大,同一亚家族的外显子数量较为稳定。CtSPL4所具有的外显子、内含子数量最多,均为12个,、、、具有的外显子、内含子数量最少,具有2个外显子和1个内含子。


2.4 红花CtSPL基因家族顺式作用元件预测
对基因启动子区段顺式作用元件进行预测分析发现,启动子区顺式作用元件主要可分为4个大类:光响应元件、胁迫响应元件、激素应答元件、生长发育响应元件(图5)。其中光响应元件最多,共有17种,包括G-Box、Box 4、GT1-motif、TCT-motif、I-box等。胁迫响应元件主要有厌氧诱导(ARE)、防御与应激(TC-rich repeats)、干旱诱导(MBS)、低温响应(LTR)、损伤响应(WUN-motif)等6种。激素应答元件有8种,其中脱落酸响应元件(ABRE)共有75个,除、、、外均有分布,说明大部分基因可能响应了红花体内的脱落酸信号。其他激素相关的顺式作用元件,如茉莉酸甲酯(CGTCA-motif和TGACG-motif)、生长素(TGA-element)、水杨酸(TCA-element)等也广泛分布。生长发育响应元件有7种,包括分生组织表达调控(CAT-box)、昼夜节律调控(Circadian)、胚乳表达调控(GCN4-motif)等。

2.5 红花CtSPL基因家族共线性分析
为了阐明红花基因家族的潜在进化机制,笔者所在课题组研究了它的复制事件,但没有发现串联重复和大片段复制(图6-A)。
为了进一步研究红花基因家族的系统进化机制,笔者所在课题组构建了红花与向日葵、拟南芥基因组之间基因的共线性图谱(图6-B),共鉴定出10个基因与向日葵存在共线性关系:、、、、、、、、、。5个基因与拟南芥存在共线性关系:、、、、。研究结果表明,红花与拟南芥之间的直系同源基因对远小于红花与向日葵之间,这可能与红花与向日葵亲缘关系更近有关,基因在同科植物之间的保守性较高。通过共线性分析可以从已知拟南芥的基因功能,推断红花中相应基因的功能。
2.6 红花CtSPL基因家族的miRNA靶位点预测分析
利用psRNATarget在线软件对下载的红花miRNA成熟序列和的cDNA序列进行分析,预测可能调控基因的miRNA, 期望值设置为3.5,其他为默认参数(图7)。中有15个基因受miRNA调控,参与调控的miRNA主要是miR156家族,其次是miR157家族,而miRN525、miRN560、miRN566分别只调控、、基因的表达。


2.7 红花CtSPL基因家族表达模式分析
利用NCBI公开的转录组数据,分析基因在不同组织、不同激素诱导下的表达特性。如图8-A所示,大多数基因均存在组织特异性表达差异。、的表达量在3个组织中均较高,在叶片中的表达量最高。部分基因仅在1个或2个组织中特异表达,如、基因只在叶片中表达;、、基因只在花瓣中表达。总之家族基因具有多样化组织表达。
经茉莉酸甲酯处理后(图8-B),基因的表达被诱导,其他还有11个基因的表达量上调,其中、、、表达量的上调最为明显,分别为对照组的101.71、45.20、19.19、15.25倍。共有9个基因在茉莉酸甲酯处理后表达量下调,其中处理后的表达量仅为对照组的8.85%。
前人收集红花种子在MS 培养基(对照)和包含1 μmol/L EBR处理的MS 培养基上生长2周后幼苗的叶,用于后续转录组测序。如图8-C所示,表油菜素内酯处理后大部分基因的表达量都呈现上升趋势,其中4个基因的表达量均上调为对照组5倍以上,分别为、、、。然而,、在表油菜素内酯处理后表达量均下调。
综上所述,基因的组织特异性表达可能和各个基因在不同组织中发挥的功能有关。、、、、、、等基因可能参与茉莉酸甲酯、表油菜素内酯等激素的调控,从而发挥多种生物学功能。
3 讨论与结论
随着高通量测序技术的飞速发展, 目前已完成了红花基因组的全长测序,为红花的分子生物学研究奠定了基础。本研究从红花基因组中鉴定出21个基因家族成员,比棉花(59个)、烟草(32个)、白菜(29个)数目少,比水稻(19个)、高粱(19个)、拟南芥(16个)等数目多,表明基因家族成员在不同的物种中具有一定的数目变异,从而导致了基因功能的多样化。对CtSPL蛋白结构域进行分析,发现有部分CtSPL在进化过程中发生变异或缺失,导致锌指结构或核定位信号保守性不高。笔者所在课题组将红花与向日葵、拟南芥基因组进行基因共线性分析,结果显示,红花基因与拟南芥、向日葵基因之间分别存在5、10个共线性基因对,、分别在与向日葵、拟南芥共线性组间均链锁了2个基因对,表明、可能是家族中较为保守的基因,其在进化过程中发挥着重要作用。通过共线性分析可以从已知拟南芥的基因功能,推断红花中相链锁的基因的功能。
对家族成员的miRNA靶基因预测结果显示,其主要受miR156家族的调控。miR156 是植物小RNA家族中表达丰度最大的家族,也是开花植物发育转变的主要调节因子。在拟南芥中,miR156-SPL10通过介导细胞分裂素反应,控制根分生组织活性和根源新生芽再生。Ding等认为,miR156-可以通过抑制下游相关热响应基因来适应环境温度变化。Zhang等发现,茶树中miR156的2个靶基因、对低温敏感,可能参与低温胁迫响应过程。Kouhi等结果表明,miR156的表达量在干旱胁迫的植株叶片中降低,因此推测,红花幼苗在长期受到轻度干旱胁迫时,miR156-可以使植株继续存活并提高开花率。热胁迫导致miR156表达增加,而叶片和根中基因下调,表明miR156可通过加速基因的裂解,从而调节红花中的应激反应。miR156-不仅参与植物的开花调控,同时具有介导植物生育阶段转变、调节植物次生代谢、参与植物光信号转导和胁迫响应等作用。本研究发现,共有12个基因可能受到不同的miR156成员调控,这为进一步探索miR156-SPL模块在红花中的功能提供了参考。
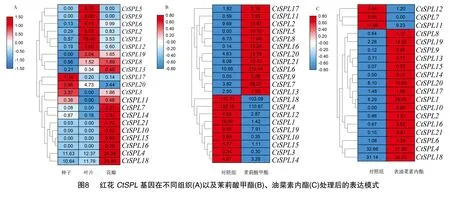
基因的组织表达模式可能与其功能特征相关,而基因在不同植物组织中表达差异显著。桃多数成员在幼果中表达量较高;不同龙眼成员在体胚发育不同阶段表达量有明显差异。本研究对基因在红花不同组织表达特性进行分析发现,成员具有不同的表达特性,同一亚家族表达模式相似。其中Ⅳ亚家族的、的表达量在3个组织中均较高,且在花瓣中最高。Ⅰ亚家族的、基因只在花瓣中表达,而同一亚家族的拟南芥和被证明参与拟南芥开花过程的调控,因此,推测红花中的这些同源基因,可能参与了红花花器官调控的相关功能。MeJA在植物胁迫响应、生长发育以及次生代谢中发挥着重要的作用。本研究中,、、、等4个成员的表达量经MeJA处理后均大幅度上调,且在顺式作用原件分析中发现,、均含有MeJA的应答元件,进一步说明能够应答MeJA信号。表油菜素内酯对提高植物的抗逆性:抗旱性、抗冷抗寒性、抗热性、抗盐性、抗药性、抗毒性以及对植物衰老的调节具有重要的作用。本研究发现,EBR处理后部分基因的表达量发生了显著的变化,说明其能响应油菜素内酯信号途径,从而参与EBR介导的植物抗逆性的调节等生理功能。
猜你喜欢
快乐语文(2021年34期)2022-01-18
快乐语文(2021年27期)2021-11-24
快乐语文(2021年11期)2021-07-20
快乐语文(2021年15期)2021-06-15
科学与财富(2021年3期)2021-03-08
——以多重共线性内容为例
长沙航空职业技术学院学报(2019年2期)2019-07-13
温州大学学报(自然科学版)(2019年2期)2019-06-04
生命科学研究(2018年1期)2018-05-29
上海农业学报(2017年3期)2017-04-10
山东农业工程学院学报(2016年6期)2016-12-01
