
黄土高原草地植物叶脉性状沿环境梯度的变化规律
2022-10-27 03:14:42高慧蓉莫惟轶王小春王志波李丹洋王瑞丽
生态学报 2022年19期
张 明,高慧蓉,莫惟轶,陈 爽,朱 荣,王小春,王志波, 李丹洋,王瑞丽,4,*
1 西北农林科技大学,杨凌 712100 2 西北农林科技大学水土保持研究所,杨凌 712100 3 中国科学院水利部水土保持研究所,杨凌 712100 4 陕西秦岭森林生态系统国家野外科学观测研究站,杨凌 712100
叶脉由木质部和韧皮部组成[1],根据在叶片中的分布形态,叶脉主要可以分为网状脉、平行脉、叉状脉和单叶脉。其中,双子叶植物多为网状脉,平行脉多存在于单子叶植物[2],叉状脉在蕨类植物中最常见,而单叶脉类型的结构多存在于针叶树中[3]。在网状脉中,一般将其较粗的中脉定义为一级,由中脉分出的为二级,依此类推,三级以上称为次级叶脉。次级叶脉的长度占到叶脉总长度的80%以上,在叶片的水分运输中发挥着极其重要的作用[1]。平行脉植物由于其特殊的形成方式,并没有明显的等级之分,因此不区分主脉和次脉[4]。
叶脉结构性状在维持叶片水分和光合能力方面具有重要的作用[1, 5—6]。叶脉密度(VLA)反映了水分、养分和光合产物等物质运输的效率[7],叶脉直径(VD)决定了叶脉间的物质分配,影响植物叶片光合吸收最大化[8—9]。Sack等[10]提出了一个新的性状指标即单位面积叶脉体积(VV),由叶脉密度和直径计算得到,来表征植物对叶脉的投资大小。许多研究发现,叶脉密度和直径间存在负相关关系[11—13],这体现了植物在叶脉性状间资源投资的权衡[14]。但也有研究发现两者间并未存在相关性,例如,Brodribb等[3]就发现单叶脉植物因为特殊的叶脉结构,密度和直径间并不存在显著的相关性。由此可见,叶脉性状间的关系可能因为叶脉类型的不同而存在很大差异。目前对于叶脉性状间关系研究多集中在网状脉植物,对于其他脉型植物中这种关系具有怎样的差异仍较少研究。
环境因子是影响叶脉性状变化的重要因素。通常干旱环境中,植物倾向于增加叶脉密度来提高供水能力,以便在降水期获得足够的水分供应[15]。Sack等[1]分析全球796个植物叶片后发现叶脉密度和年降水量呈负相关。除了水分外,温度和光照也会对叶脉性状产生影响,一般随温度升高,植物的次级叶脉密度会增大。光照增强也会导致叶脉密度增大,通常阳生植物的叶脉密度要高于阴生植物[1]。Uhl等[15]还发现土壤养分越贫瘠的地区,植物的叶脉密度往往越大。因此,叶脉性状对于不同的环境因子所做出的响应往往不同。此外,研究发现植物在进化的过程中,随气候和大气成分的不断改变,被子植物逐渐具有高的叶脉密度,而非被子植物始终具有稳定的低叶脉密度[16—18]。不同生长型植物的叶脉性状在适应环境的过程中也存在很大差异。例如,叶脉密度通常在乔木中最高,灌木次之,草本植物中最低[1]。Wang等[11]研究发现乔木植物的次级叶脉直径随海拔升高显著增加,叶脉密度变化不明显,而灌木则呈相反趋势。从这些研究中可以发现,不同类型植物的叶脉性状对环境因子的响应不同。那么在不同叶脉类型植物中,随环境变化叶脉性状间的关系及叶脉沿环境梯度又会发生怎样的变化,却不得而知。
黄土高原位于我国的中北部,水土流失严重,生态环境脆弱,恶劣的环境条件使得植物生存和生长面临严峻考验。叶脉结构是植物保障水养运输和机械支撑的重要条件,对于植物在恶劣生境中的生存具有重要意义[1]。目前研究发现叶脉性状随环境的变化呈现出一定的经度地带性规律[12, 15],而黄土高原降水量和土壤养分含量从东向西均呈现出递减的趋势,在经度上形成了明显的梯度变化[19],是研究不同脉型植物叶脉性状沿环境梯度变化的理想场所。因此本研究沿水分和养分梯度在黄土高原从东到西选取了10个样地,对样地内常见物种的叶片进行取样,考察不同脉型植物(包括网状脉、平行脉和单叶脉)叶脉性状间关系是否存在差异?其叶脉性状沿环境梯度如何变化?从大尺度上揭示叶脉在黄土高原水平方向上的变异规律,旨在从生理结构的角度探究植物叶片对环境变化的适应策略,从而为黄土高原地区植被恢复提供理论参考。
1 研究区概况
黄土高原地处东经101°00′—114°33′,北纬34°00′—45°05′,横跨中国青、甘、宁、内蒙古、陕、晋、豫7省,面积64万km2,海拔200—3000 m。该地区年降雨量为150—800 mm,且多集中于6—9月(占全年降水量的55%—78%),年平均降水量从东南向西北递减,地跨干旱、半干旱和半湿润3个气候带。黄土高原中的植被由5种主要的自然植被组成,即沙漠、荒漠草原、草原、森林草原和森林,沿着高原从西北到东南逐渐增加的降水梯度分布。天然草地类型以白羊草(Bothriochloaischaemum(L.) Keng)、羊草(Leymuschinensis(Trin.) Tzvel)、铁杆蒿(ArtemisiagmeliniiWeb.ex Stechm)、针茅(StipacapillataL.)、胡枝子(LespedezabicolorTurcz)等群落类型为主[20]。
2 研究方法
2.1 取样方法
2.1.1取样点设置
以干旱指数反映水分的变化,土壤全氮、全磷含量反映养分的变化,黄土高原水分和养分含量自东向西逐渐减少(图1),形成了由东部湿润肥沃向西部干旱贫瘠逐渐过渡的环境,在经度上具有明显的梯度变化[21],同时由于环境因子间具有一定自相关,因此以经度梯度来综合代表水分和养分梯度的变化。在经度上沿水分和养分梯度共选取10个取样点进行取样。
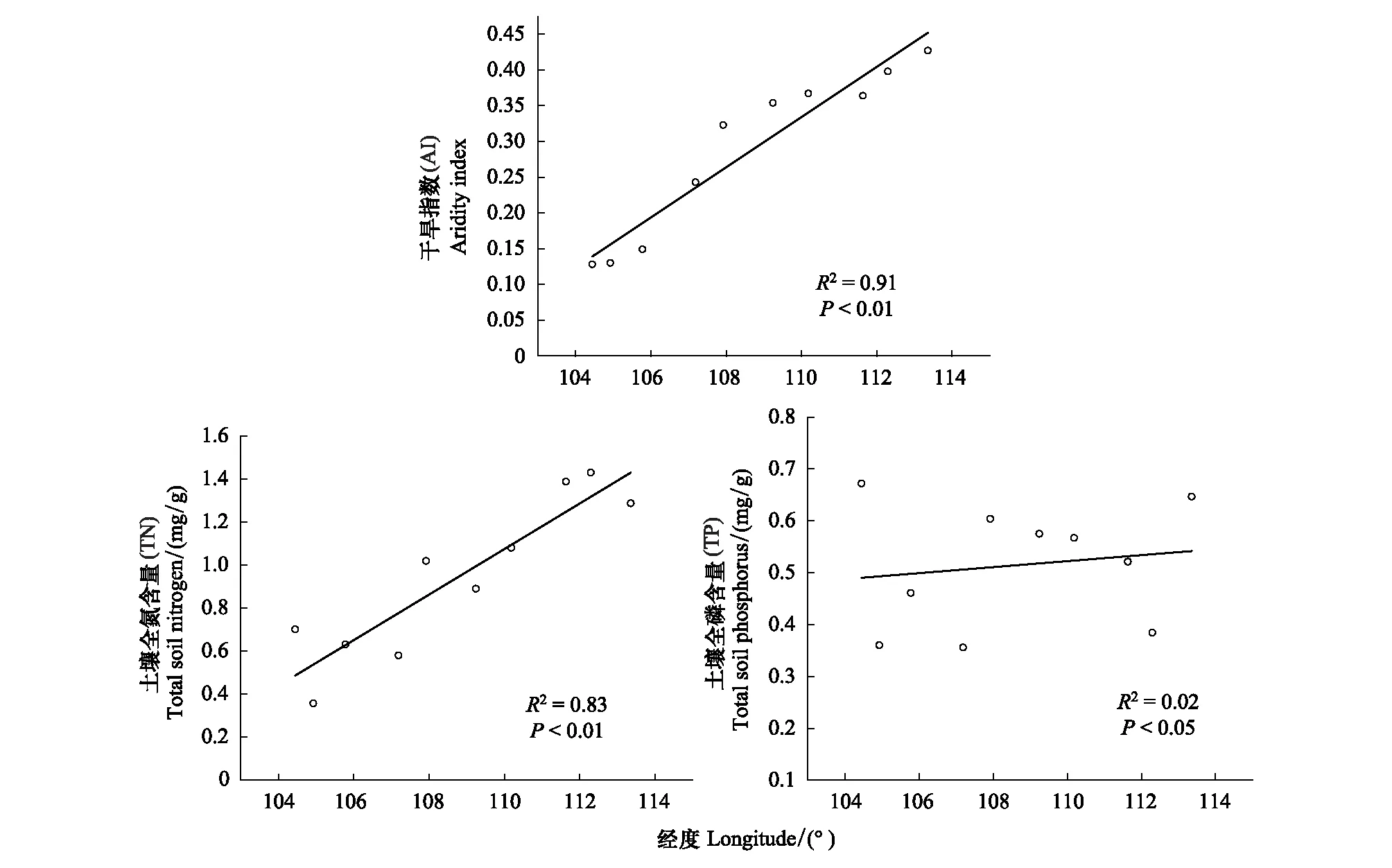
图1 黄土高原AI、TN、TP沿经度变化规律Fig.1 Variation of AI, TN and TP along the longitude on the Loess PlateauAI越小,表明干旱程度越大,数据来源于https://cgiarcsi.community
2.1.2野外取样
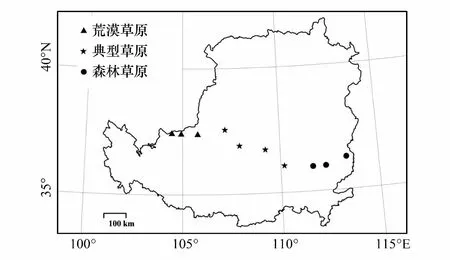
图2 黄土高原范围及取样点示意图 Fig.2 Schematic diagram of the Loess Plateau space and sampling points
2018年7—9月在黄土高原中部对设置的10个取样点进行取样(图2和表1)。取样点在经度上横跨整个黄土高原,相邻取样点之间间隔100 km左右,包括森林草原、典型草原和荒漠草原3种草地类型(基本信息见表1)。
在每个取样点内,选取能够代表当地植被类型的区域,采用样线法布设样方。按照一定方向布设100 m的样线,等间隔设置8个1 m×1 m的样方进行群落结构调查,确保相邻样方间间隔大于10 m。在每个样地半径2 km范围内,对样地内出现的常见物种进行取样。选取每个样方内每一个物种的3—5个个体,每个个体上选择健康干净、完全展开的叶片10—20片,放入10号自封袋内保存,尽快带回室内。随后,将每个物种叶片避开主脉剪至1 cm×1 cm大小,用FAA(5 mL 38%甲醛溶液:5 mL冰醋酸:90 mL 50%乙醇:5 mL甘油)固定液保存到10 mL离心管中。共获得519种植物的叶片,属于57科156属。
对叶片取样的同时,在每个样方内进行土壤样品的采集,在去除土壤表面的杂物后,用内径为5 cm的土钻对0—10 cm的土层进行取样,每个样方随机各取5处,将取出的土样混合均匀带回室内处理。
2.2 室内测定
2.2.1叶脉性状测定
叶脉密度(VLA)与叶脉直径(VD)的测定采用化学处理法。具体步骤为:每一个物种个体从固定液中随机选取3—5个叶片,使用7% NaOH溶液浸泡,且每2—3天更换一次溶液,可根据叶片变透明的情况适当增加NaOH的浓度,直到叶片透明。之后使用蒸馏水浸泡30 min,再转入5% NaClO溶液中5 min,后置于蒸馏水中浸泡3 min,选择3个透明度较高的叶片制成临时切片并使用1%番红试剂染色。然后将切片置于MOTIC显微镜(型号:BA210,产地:厦门)下,并利用配置的摄像装置进行观测,网状脉与单叶脉样品在20倍物镜、10倍目镜条件下观测,平行脉样品在10倍物镜、10倍目镜条件下观测,每一个叶片随机选择5个视野拍照,每个样品共有15个视野,最后从中挑选5个视野,利用配套的分析软件测定次级叶脉的长度与直径。
不同叶脉类型的叶脉性状计算方法如下。其中,网状脉的次级叶脉直径(VD,μm)为所有次级脉直径的平均值,次级叶脉密度(VLA,mm/mm2)为单位面积叶脉总长度(图3)。计算公式如下:
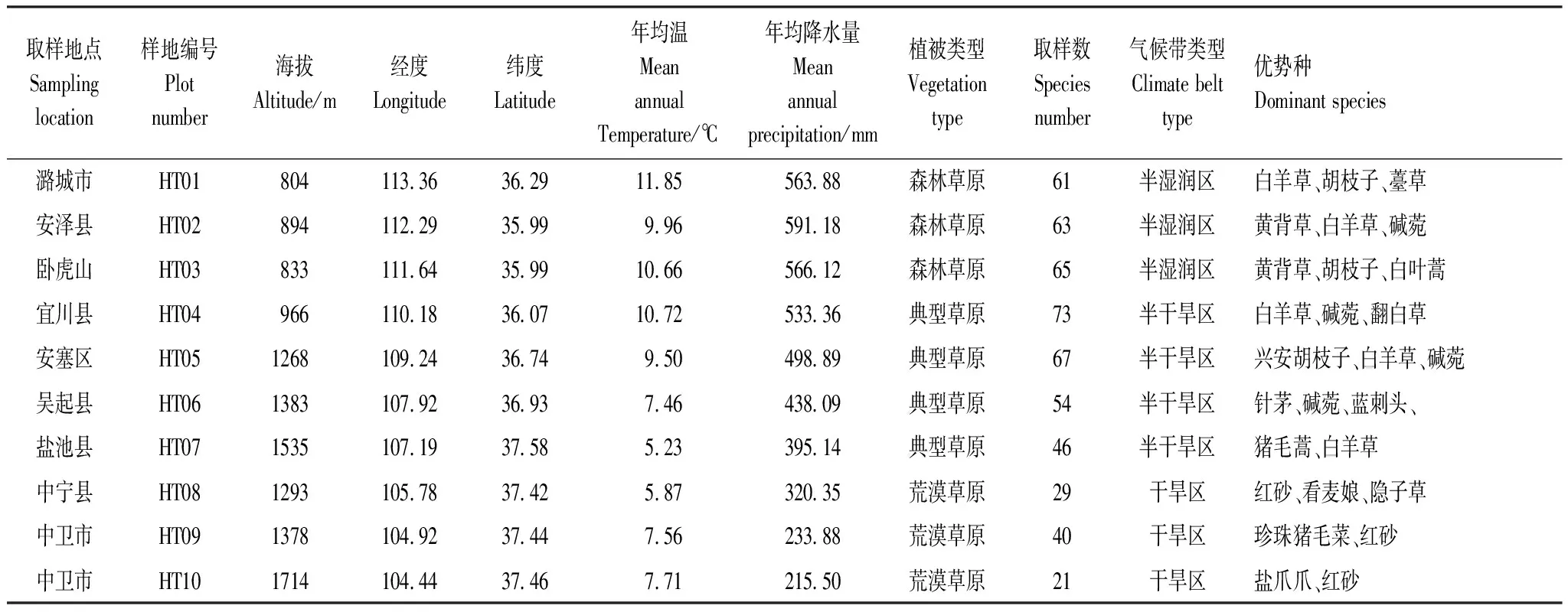
表1 取样点相关信息
VLA=视野中的叶脉总长度/视野面积
(1)
叶脉体积(VV,μm3/mm2)采用Sack等[10]的方法:
VV=π×(VD/2)2×VLA
(2)
平行脉具有大纵向脉、小纵向脉和横向脉之分,且不同等级叶脉性状差异较大,故将各等级叶脉密度分开测量,之后求其总和(图3):
VLA=大纵向脉密度+小纵向脉密度+横向脉密度
(3)
由于平行脉存在多个等级的叶脉直径且不同等级间的叶脉直径差别较大,以每个等级叶脉的相对长度为权重,计算得到不同等级的叶脉直径加权平均值:
VD=大纵向脉直径×(大纵向脉长度/总长度)+小纵向脉直径×(小纵脉长度/总长度)+横向脉直径×
(横脉长度/总长度)
(4)
VV=大纵向脉体积+小纵向脉体积+横向脉体积
(5)
对于单叶脉物种,其叶脉性状的测定方式与网状脉相同(图3)。
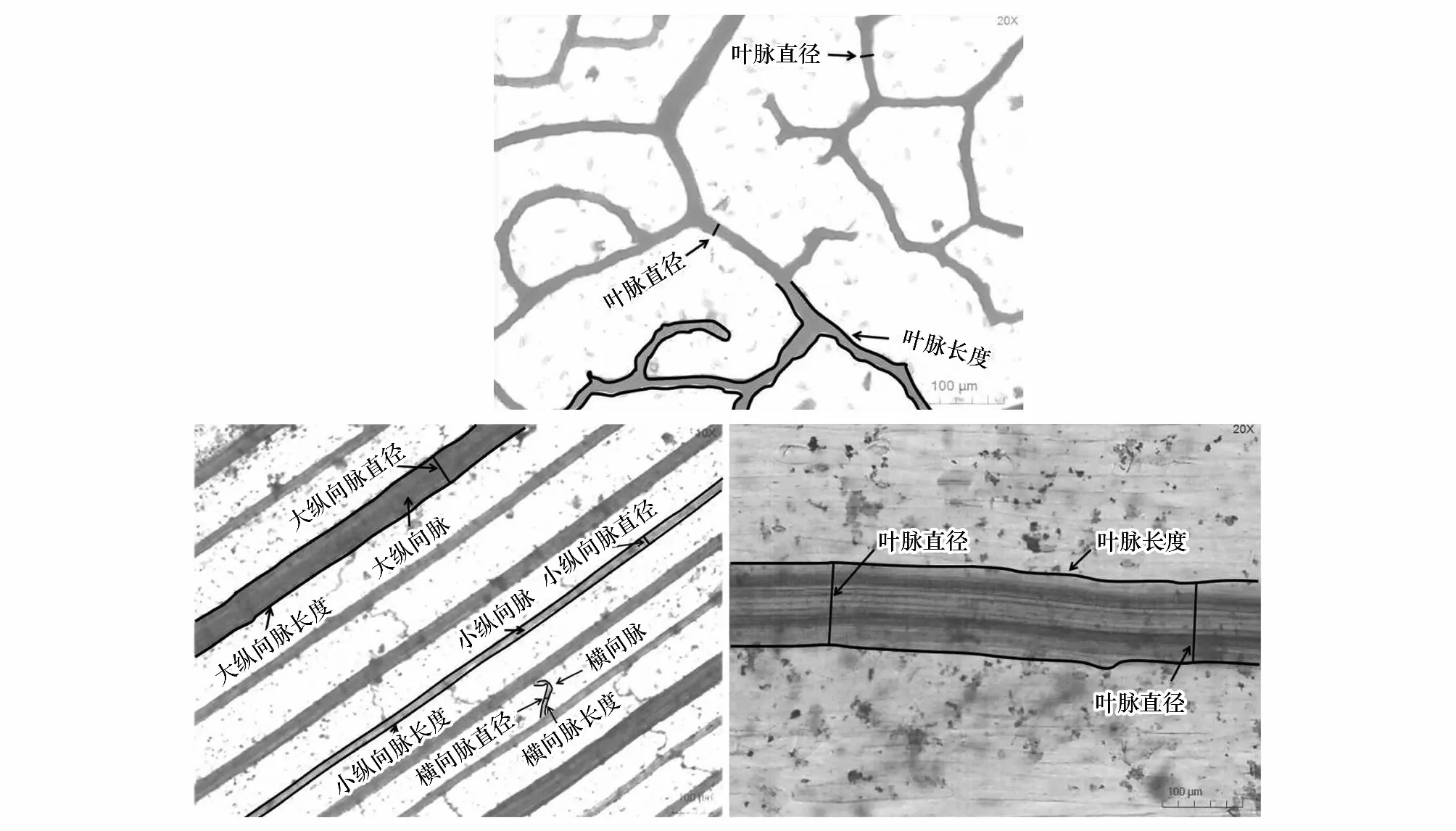
图3 不同脉型植物叶脉性状的测量Fig.3 Measurement of leaf vein traits in plants with different vein types叶脉直径每个视野测5个取其平均值,平行脉大纵向脉,小纵向脉和横向脉分开测量
2.2.2土壤理化性质的测定
将土壤样品在室温条件下自然风干,去除植物根和石砾等杂物,过2 mm土壤筛后,用球磨仪(MM400 ball mill,Retsch,Germany)和玛瑙研钵(RM200,Retsch,Haan,Germany)研磨。研磨好的样品装入自封袋中用于测定土壤元素含量。用烘干法对土壤含水量(SWC)进行测定;使用容量分析法测定土壤有机碳(TOC)含量;土壤全氮(TN)含量采用元素分析仪(Vario MAX CN Elemental Analyzer,Elementar,Germany)测定,土壤全磷(TP)含量的测定采用电感耦合等离子体发射光谱仪(ICP-OES,Optima 5300 DV,Perkin Elmer,Waltham,MA,USA),土壤pH采用多参数电导/ pH表(MYRON L Ultrameter II TM,USA)测定。
2.3 气象数据的获取
每个取样点的年均温(MAT)、年降水(MAP)来自于中国气象站数据的插值结果。对国家气象局提供的研究区域及附近省份的气象台站数据,应用ANUSPLINE插值软件将其插值到0.1°×0.1°经纬度网格上,根据样点经纬度从插值结果中提取样点的1961—2010年平均气温和年降水量数据(http://www.cma.gov.cn/)[20]。干旱指数数据(Aridity index,AI,降水量/蒸散量)来源于全球栅格数据(CGIAR-CSI,https://cgiarcsi.community),从CGIAR-CSI数据库中提取1970—2000年干旱指数数据[22]。
2.4 数据处理
首先,对所有叶脉性状进行以10为底的对数转换,使数据满足标准化正态分布。对水分和养分反应指标与经度进行拟合,利用Pearson相关分析各指标间的相关性。使用单因素方差分析(one-way ANOVA)比较不同叶脉类型间叶脉性状的差异,针对存在显著(P<0.05)的性状关系采用标准化主轴估计(SMA)方法来分析叶脉性状关系是否在不同叶脉类型间存在显著差异。即利用方程y=bxa表示不同叶脉性状间的关系,线性转化为 log(y)= log(b)+alog(x),x、y表示两个不同性状,b代表性状关系的截距,a表示相关性的斜率。该分析由软件(S)MATR Version 2.0来计算完成。
通过回归分析对不同叶脉类型的叶脉性状与经度进行拟合来探究其沿环境梯度的变化,拟合时选取了R2最大、P值显著(P<0.05)的回归方法进行拟合。使用冗余分析(RDA)来量化叶脉类型(网状脉、平行脉和单叶脉)、气候和土壤因子对叶脉性状的影响。在气候和土壤两组变量内,为了避免共线性,用逐步回归分析筛选出对叶脉性状影响显著的变量(P<0.05),并利用anova函数计算逐步回归分析筛选出来的单个因子贡献率。随后,利用RDA进一步分析这些具有显著影响的气候和土壤因子对叶脉性状的单独作用和交互作用。以上分析在 SPSS 20. 0 与 R 3. 5. 2 软件 vegan 程序包中完成,文中所用到的数据图都采用Origin 2017软件绘制。
3 结果
3.1 黄土高原植物叶脉性状概况
在所有物种中,VLA、VD和VV的平均值分别为11.29 mm/mm2、22.05 μm、51.15×105(μm3/mm2)。碱蓬(Suaedaglauca)的VLA最大,刺柏(Juniperusformosana)最小,而VD最小的是铁苋菜(Acalyphaaustralis),最大的是天门冬(Asparaguscochinchinensis)。VV的最大值和最小值则分别出现在羽茅(Achnatherumsibiricum)和黄鹌菜(Youngiajaponica)中。
其中,VLA(CV=0.38)的变异性小于VD和VV的变异性(CV分别为1.01和1.77,表2)。不同叶脉类型其性状具有显著差异(P<0.05,表2)。其中,网状脉的VLA显著大于平行脉和单叶脉(P<0.05),而VD和VV则相反。单叶脉的VD和VV的变异程度最大(CV分别为0.5和0.92),平行脉的VLA变异程度最大(CV=0.44)。
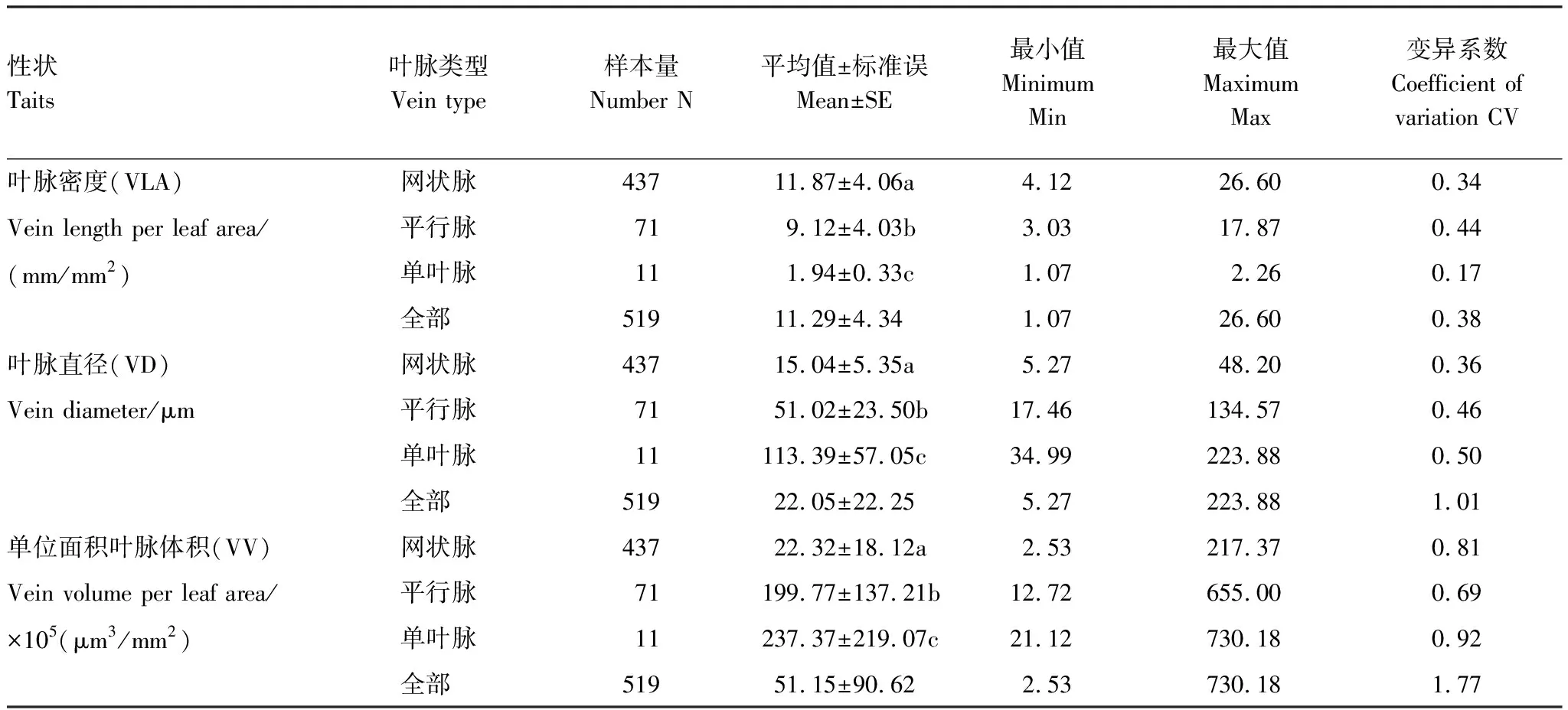
表2 不同叶脉类型叶脉性状数据描述
3.2 不同脉型植物叶脉性状关系的差异
VLA和VD的负相关关系在网状脉和平行脉及全部物种中都存在(R2=0.07—0.30,P<0.01),然而这一关系在单叶脉中并未发现(图4,P>0.05)。VLA和VV在全物种中表现出负相关,而在网状脉植物中表现出正相关(图4,P<0.01),在平行脉和单叶脉中没有发现两者的相关性。VD和VV的正相关关系在全物种和3种不同叶脉类型的植物中都存在(图4,P<0.01),斜率和截距之间都存在显著差异(P<0.01),其中网状脉的斜率最大,而平行脉的截距最小。
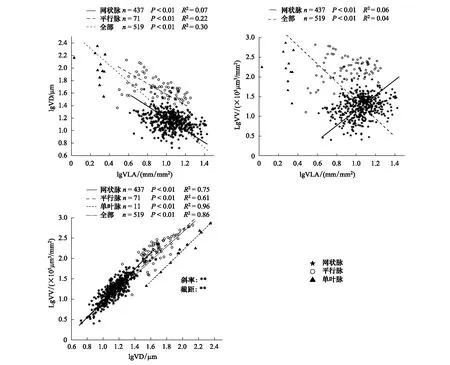
图4 不同叶脉类型叶脉性状间关系的标准化主轴回归关系Fig.4 Standardized major axis estimation regression among vein traits of different vein type plantsVLA:叶脉密度 Vein length per leaf area;VD:叶脉直径 Vein diameter;VV:单位面积叶脉体积 Vein volume per leaf area 只列出显著的回归关系(P<0.05);** P<0.01
3.3 不同脉型植物叶脉性状沿经度的变化
叶脉性状沿经度的变化规律在不同叶脉类型的植物中存在差异(图5)。当考虑所有物种时,VD和VV随经度增加而显著减小(P<0.01),而VLA随经度的增加并没有显著的变化(P>0.05)。在网状脉植物中,VD和VV随经度的增加均显著减小(P<0.01),虽然VLA随经度增加也减小,但其变化较弱(R2=0.02)。与网状脉不同,平行脉植物的VD和VV随经度的增加则显著增大(P<0.01),而VLA随经度的增加并没有显著变化(P=0.76)。在单叶脉植物中,三种叶脉性状沿经度变化均不显著(P>0.05)。
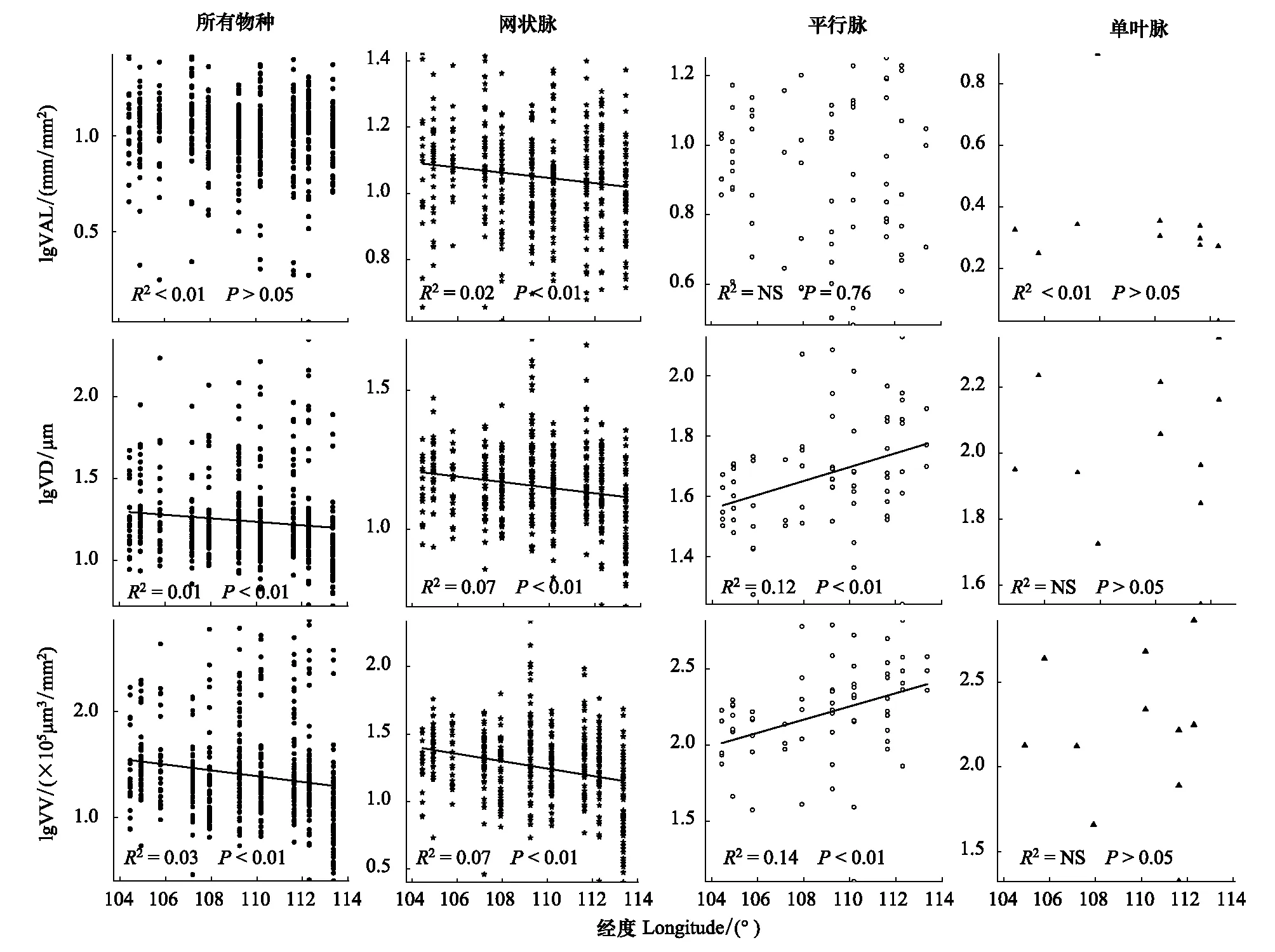
图5 不同叶脉类型植物叶脉性状沿经度变化规律Fig.5 Variation in leaf vein traits of different vein type plants along the longitude
3.4 环境因素对叶脉性状的影响
利用逐步回归分析筛选出对叶脉性状有显著影响的气候和土壤因子,再利用冗余分析进一步分析这些环境因子对叶脉性状的独立和交互作用,结果显示(图6):气候因子、土壤因子和叶脉类型三者的总效应对叶脉密度、叶脉直径和体积的解释度分别为30.15%、67.90%和56.96%。在独立效应中,叶脉类型对性状变异的解释量最多,其中对叶脉密度的影响最小(28.74%),而对叶脉直径的影响最大(62.25%)。土壤因子解释了叶脉性状0.54%—2.91%的变异性,其中影响叶脉密度的因子主要是土壤有机碳含量(TOC)和总磷含量(TP),而直径和体积的主要土壤因子是土壤含水率(SWC);气候因素解释了叶脉性状0.02%—0.77%的变异性,影响叶脉密度的因子主要是年均温(MAT),影响直径和体积的是年降水量(MAP)和干旱指数(AI)。气候、土壤、叶脉类型三者的交互作用解释了叶脉性状-0.45%—3.56%的变异性。
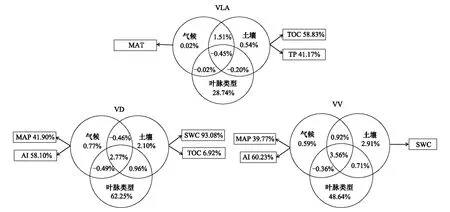
图6 气候、土壤和叶脉类型对叶脉性状变异的解释度Fig.6 Variation of leaf vein traits explained by climate, soil and leaf vein typesMAT:年均温;TOC:土壤有机碳含量;TP:土壤总磷含量;MAP:年降水量;AI:干旱指数;SWC:土壤含水率
4讨论4.1不同脉型植物叶脉性状的关系
与全球叶脉性状数据相比,黄土高原网状脉、平行脉和单叶脉植物的平均叶脉密度(分别为11.87、9.12、1.94 mm/mm2)均要高于全球平均水平(分别为7.36、7.25、1.52 mm/mm2)[1, 10],但叶脉直径均值(22.05 μm)低于全球平均水平(26.42 μm),而叶脉体积(51.15×105μm3/mm2)又高于全球平均水平(38.21×105μm3/mm2)[1]。造成这种差异主要与黄土高原的气候条件有关,植物以细而长的叶脉来适应干旱生境,从而保证水分的运输和防止栓塞的产生[23],同时较高的叶脉体积意味着黄土高原植物在适应环境的过程中对叶脉的高投资[10]。
本研究中,网状脉和平行脉植物叶脉密度和直径存在显著负相关(图4,P<0.01),这与已有的研究成果相同[10—12]。目前关于这种权衡关系的解释主要有投资收益和空间限制两种假说。投资收益假说认为,叶脉密度越大,叶片就会有更强的水分运输和光合能力[24—25],但是高叶脉密度也意味着高的建造成本[26],而空间限制假说认为,如果叶脉分布密集且直径很大,因为CO2无法通过木质部,就会导致气孔和光合组织之间的连接被切断[27],因此,不管是从投资收益还是空间限制的角度来看,叶脉变细变长是植物收益最大化的最优选择[6]。
与网状脉和平行脉不同,在单叶脉植物中,并未发现密度和直径间的相关性(图4,P>0.05)。这可能是因为在样本中,单叶脉植物多数为裸子植物,与被子植物相比,许多裸子植物的叶肉组织含有充满水分的木质化细胞,如石细胞等[28],它们穿插在整个叶肉组织中,作为水分运输到蒸发点的通道[2],这意味着水分可以通过叶脉之外的其他途径来运输。其次,因为单叶脉植物只有一条直线的叶脉,在视野中的长度差距不大(本研究中单叶脉植物叶脉密度的变异系数为0.17),但各物种的直径差距较大(CV=0.5),从而造成了密度与直径的不相关。此外,与其他叶脉类型相比,本研究中单叶脉植物的数量较少(n=11),数据分析时不易达到显著水平。未来需要对单叶脉植物进行更多物种和更大空间上的取样,来检查叶脉性状间关系在不同叶脉类型植物中的普适性。
叶脉体积作为变异性最大的一个性状(CV=1.77),反映了叶脉能够运输水分的量以及植物对叶脉的碳投资大小[10]。叶脉体积是由叶脉密度和叶脉直径计算得到的,理论上它应该和两者都存在正相关关系,但是在本研究中密度与体积的关系在不同叶脉类型植物中表现出较大差异。叶脉密度与体积之间的正相关关系只在网状脉植物存在,在所有物种中二者反而呈负相关,但这些关系均较弱(R2=0.04—0.06,图4)。与叶脉密度不同,叶脉体积与叶脉直径之间的正相关关系在不同叶脉类型植物中均存在,且相关性较强(R2=0.61—0.96,图4)。这表明叶脉体积主要受到叶脉直径,而不是叶脉密度的影响。
4.2 植物叶脉性状沿环境梯度的变化规律因叶脉类型而异
从黄土高原东部到西部,随着经度降低,所有物种中叶脉直径和体积均增大(P<0.01),但密度并没有显著变化(P>0.05,图5)。叶脉类型差异是影响叶脉性状变异的主要因素,解释了叶脉性状27.84%—62.25%的变异性(图6)。环境因子中,土壤因子(主要是土壤水分和养分含量)对3种叶脉性状的解释度(0.54%—2.91%)大于气候因子(0.02%—0.77%)。
不同脉型植物的叶脉性状沿经度的变化趋势差异较大(图5),反映了不同脉型植物适应环境的不同策略。其中,网状脉次级叶脉密度的变化趋势与其他研究结果一致,即在水分较少的环境中次级叶脉密度会增大[15, 29]。黄土高原自东向西随经度减小,降水量逐渐减少(表1),次级叶脉密度逐渐增大(图5),这主要是因为随环境干旱程度增大,蒸腾需求增加,植物通过增大叶脉密度来保障水分的供应[5]。同时,随经度减小,土壤养分含量逐渐减少,土壤变得贫瘠,植物通过增加次级叶脉密度来获得更大的养分运输能力,提高植物对养分的吸收和利用,这与Ulh等[15]的研究结果一致。
导管直径大小与栓塞脆弱性正相关,因此普遍认为干旱生境下树种的木质部导管直径较小[30]。然而,在本研究中干旱地区的网状脉植物直径却更大(图5)。这可能是因为黄土高原环境条件特殊,属于干旱半干旱地区[31],物种多具有抗旱耐旱的能力[32]。木质部栓塞是耐旱树种对水分胁迫的一种适应性反应[33],适当的木质部栓塞对植物的节水和在干旱环境中生存是有利的,这是植物在长期发展中形成的一种节水策略[34—35]。申卫军等[33]发现即使发生栓塞,在降雨或其他条件使水势升高时,栓塞能够很快恢复。黄土高原水土流失严重,土壤水分、养分补充能力较差,补给很大部分来自于降水[36],而降水主要集中在6—9月,这种水热并存的气候条件对植物的生长是极为有利的[37]。因此,在此期间,木质部栓塞对水分运输所造成的损失可能要小于增加木质部对树木导水所带来的收益,因此植物选择通过增大次级叶脉直径来获得最大的生长速率。
值得注意的是,尽管网状脉植物叶脉性状沿经度具有显著的地理格局,但这些变化趋势均较弱(R2=0.02—0.07,P<0.01,图5),这可能是由于叶脉性状具有高的系统发育保守性[11, 16, 38]。以往对网状脉的研究结果表明,其叶脉性状相比于其他叶片性状,系统发育信号更强,对环境变化的可塑性较小[39],尤其是次级叶脉密度在叶脉发育的过程中基本始终保持稳定[10]。这种系统发育保守性可能掩盖了环境对网状脉植物叶脉性状的可塑性。
平行脉植物随经度增加叶脉密度变化不显著(图5,P>0.05),而叶脉直径和体积表现为极显著的增大(P<0.01),即在养分和水分相对丰富的地区,平行脉植物具有更高的叶脉直径和体积。这主要是因为平行脉植物多为C4植物,光合能力较强,叶脉密度相对偏大[40]。并且多数平行脉植物的叶片相对较窄,对叶脉进行投资时,因为空间的限制,很难再对叶脉密度进行投资,这也符合空间限制假说。所以,黄土高原平行脉植物在适应干旱环境的过程中更多的去调整叶脉直径的变化。与之类似,Wang等[41]对青藏高原不同叶脉类型植物的研究中也发现了这一规律。单叶脉植物的3种性状随经度的变化均不显著(图5,P>0.05),这可能是由于本研究中单叶脉植物种类较少,且分布较为零散。此外,单叶脉具有特殊输水组织如石细胞等,因此单叶脉植物在应对水分短缺时可能会采取改变叶脉性状之外的适应方式。这些原因可能导致了黄土高原单叶脉植物叶脉性状未表现出明显的地理格局。
5 结论
黄土高原不同叶脉类型植物的叶脉密度和体积均高于全球平均水平,但叶脉直径均值低于全球平均水平,表明黄土高原植物整体上具有细而长的叶脉。黄土高原植物叶脉密度和直径存在显著负相关,且自东向西随经度减小,叶脉直径和体积均增大,而密度则无显著变化。但在不同脉型植物中,叶脉密度和直径的负相关关系只在网状脉和平行脉中存在(P<0.01),密度和体积的正相关则只在网状脉中存在(P<0.01),而直径和体积的正相关关系在3种脉型植物中都存在(P<0.01)。
不同脉型植物的叶脉性状沿环境梯度变化存在差异。黄土高原自东向西随经度减小,网状脉植物叶脉直径和体积均增大,而平行脉则减小,这说明对网状脉植物而言,木质部的增加对导水所带来的收益要大于栓塞的损害,因此在干旱生境中植物选择增大直径来保证生长,与之不同,平行脉则选择减少木质部的投入,降低叶脉直径,防止栓塞来适应干旱环境。在单叶脉植物中,3种叶脉性状沿经度均无显著变化。这些结果表明,不同叶脉类型的植物在适应环境变化时采取了不同的生态策略。
猜你喜欢
中学政史地·高中文综(2023年3期)2023-04-14 09:41:43
中学政史地(2023年8期)2023-04-14 07:19:08
动漫界·幼教365(大班)(2021年11期)2021-11-20 07:49:02
发明与创新(2020年26期)2020-06-27 07:25:06
小学生学习指导(低年级)(2019年9期)2019-09-25 07:43:16
红领巾·萌芽(2017年2期)2017-03-09 15:25:16
公民与法治(2016年3期)2016-05-17 04:09:00
公民与法治(2016年1期)2016-05-17 04:07:56
中国火炬(2015年7期)2015-07-31 17:39:57
世界科学(2014年8期)2014-02-28 14:58:25
