
滇西北退化高寒草甸植物群落结构对刈割的响应
2022-10-27 03:14马燕丹郑秋竹梁克敏陈欢欢岳海涛
生态学报 2022年19期
马燕丹,郑秋竹,张 勇,崔 媛,梁克敏,陈欢欢,岳海涛
1 西南林业大学云南省高原湿地保护修复与生态服务重点实验室, 昆明 650224 2 西南林业大学国家高原湿地研究中心/湿地学院, 昆明 650224 3 曲靖师范学院生物资源与食品工程学院, 曲靖 655011
高寒草甸是重要的陆地生态系统[1],在青藏高原地区,近90%的生态系统服务价值来自高寒草甸[2]。气候变化和不当的人类活动已导致青藏高原约4.5×107hm2的高寒草地发生了不同程度的退化,其中,约1/3的草地退化形势严峻[3]。除了植被覆盖率和生产力下降外[4],物种数减少[5]、毒杂草入侵、优良牧草比例降低[6]、优势种变化[7—8]等也是青藏高原高寒草地退化的常见现象。除了上述传统指标外,群落连接性、组织程度和关键种也是描述植物群落结构变化的有效指标[9—11]。研究表明,高寒草甸退化后,植物群落连接度、组织程度会下降[12],关键种也会发生更替[13]。目前,青海、藏北、甘南以及滇西北等高寒地区对退化高寒草甸植物群落物种多样性、生物量和土壤养分变化的研究较多[14—17],但对退化高寒草甸植物群落结构对外界干扰响应敏感性的关注较少。
刈割是一种有效的草地管理方式[18—19],同时也是高寒草地响应环境变化研究中相对容易实施的控制实验手段[20—21]。刈割对草甸植物群落的影响主要集中在植物群落盖度、高度、物种多样性以及地上生物量等方面[22]。研究表明,重度刈割可导致高寒草甸植物群落总盖度、平均高度、物种均匀度以及地上生物量降低[23—24]、土壤养分含量下降[25],优势物种重要值降低[24]等。但适当的刈割处理可增加植物分蘖密度和相对增长率,进而提高草地植物群落的补偿生长能力[26]。另外,适当的刈割会抑制优势物种的生长,产生新的生存空间,促进土壤种子库中的种子萌发和生长[27],有助于提高植物种丰富度[14]。
滇西北位于青藏高原南端,该区域的高寒草甸分布特征为局地集中、零散分布[28]。滇西北地区大部分高寒草甸分布在迪庆藏族自治州的高山地带,据迪庆藏族自治州香格里拉市(原中甸县)国土部门1995年发布的土地利用现状调查资料,该州70%以上的高寒草甸集中分布在香格里拉市境内。同青藏高原其他区域一样,受气候变化和不合理利用的影响,香格里拉市境内的高寒草甸也发生了明显退化[29]。为探究退化高寒草甸植物群落结构对干扰的响应特征,本研究以香格里拉市的典型退化高寒草甸为研究对象,通过控制刈割实验,分析不同退化程度高寒草甸植物群落物种丰富度、群落连接度、组织程度及关键种的变化规律。本研究拟验证以下假设:滇西北地区高寒草甸植物群落结构对刈割干扰的响应敏感性随退化程度增加而降低。
1 材料与方法
1.1 实验设计
1.1.1研究区概况及退化梯度设置
本研究的实验样地位于云南省迪庆藏族自治州香格里拉市石卡雪山山麓(99° 38′ 49.7″ E,27° 48′ 03″ N,平均海拔3310 m),是纳帕海国际重要湿地的面山地带,对纳帕海国际重要湿地的保育有重要作用[30],也是当地重要的旅游资源。研究区属寒温带山地季风气候,夏秋多雨、冬春干旱,区域年均降水量619.5 mm,年均温6.9℃,土壤类型主要为亚高山草甸土[23, 31]。杂类草是研究区内高寒草甸的优势功能群植物。高强度的旅游践踏使该区域的高寒草甸被纵横交错的道路分割成许多破碎的斑块。根据道路宽度、土壤含水量及道路上的植被覆盖度,本研究设置了3个退化梯度:严重退化(记作S1,其道路宽度为2.5—3.0 m,土壤含水率为18.2%—35.97%,道路植被覆盖度为5%—50%,距道路10 cm处的植被盖度为40%—78%)、中度退化(记作S2,其道路宽度为1.8—1.9 m,土壤含水率为35.76%—64.21%,道路植被覆盖度为65%—85%,距道路10 cm处的植被盖度为68%—85%)和轻度退化(记作S3,无道路干扰,土壤含水率为52.09%—67.06%,植被盖度为85%—100%)。各退化梯度的优势植物详见参考文献[32]。2018年以前,研究区内高寒草甸作为夏季牧场使用,同时受旅游活动的干扰;自2018年始,研究区的高寒草甸采用围栏封育的方式进行修复。
1.1.2刈割实验
每个退化梯度设置3个重复样地(即选择3条同级别的道路),在每个样地中,于距路边10 cm处设置1个1 m×10 m的调查样带,从样带中随机选取3个1 m×1 m的小样方进行刈割实验(图1)。于2018—2020年的7月进行刈割,植物留茬高度为1 cm。每年在进行刈割前记录样方中的植物物种数、物种高度和投影盖度。

图1 刈割实验设计示意图Fig.1 The mowing experiment designS1-1…S1-3: 严重退化梯度S1的三个重复样地;S2-1…S2-3: 中度退化梯度S2的三个重复样地;S3-1…S3-3: 轻度退化梯度S3的三个重复样地
1.2 数据处理
通过网络分析分析群落的复杂度和关键种。
植物群落网络构建分为三个步骤[13]:(1)计算植物种在群落中的重要值(IV),其计算公式为:IV=(RC+RH)/2,其中,RC为相对盖度,RH为相对高度[33];(2)基于物种重要值,计算物种间的Pearson相关系数r;(3)将物种作为节点,将相关性系数r达到显著性(α=0.05)的种间连线作为边进行无向网络构建。用网络连通性指标衡量群落复杂度,用模块化指标衡量群落组织程度。从群落结构维持的角度出发,通常将网络中Hub值最大的节点(物种)确定为关键种[34]。本研究将Hub值排在前10位的物种逐一剔除,以分析高Hub值物种对群落结构的作用[35],从而验证关键种识别的准确性。本研究的网络分析在Gephi 9.2中完成。
1.3 统计分析
采用双因素方差分析分析退化程度和刈割对物种丰富度的影响;采用线性回归分析探讨高Hub值物种剔除对群落连通性的影响,以上分析均在 SPSS 26中完成。采用ANOSIM分析草甸植物群落在不同退化程度和不同刈割时间上的差异,该分析使用Vegan包在R4.0.4中完成。
2 结果与分析
2.1 草甸植物群落组成对刈割的响应特征
由表1可知,退化、刈割以及它们的交互作用对S1、S2和S3的植物丰富度有显著影响。刈割前(2018年),S2和S3的物种丰富度显著高于S1(P<0.05);刈割后,S1的物种丰富度显著增加(P<0.05),S2和S3的物种丰富度无显著变化(P>0.05)(图2)。

表1 退化和刈割对草甸物种丰富度的影响
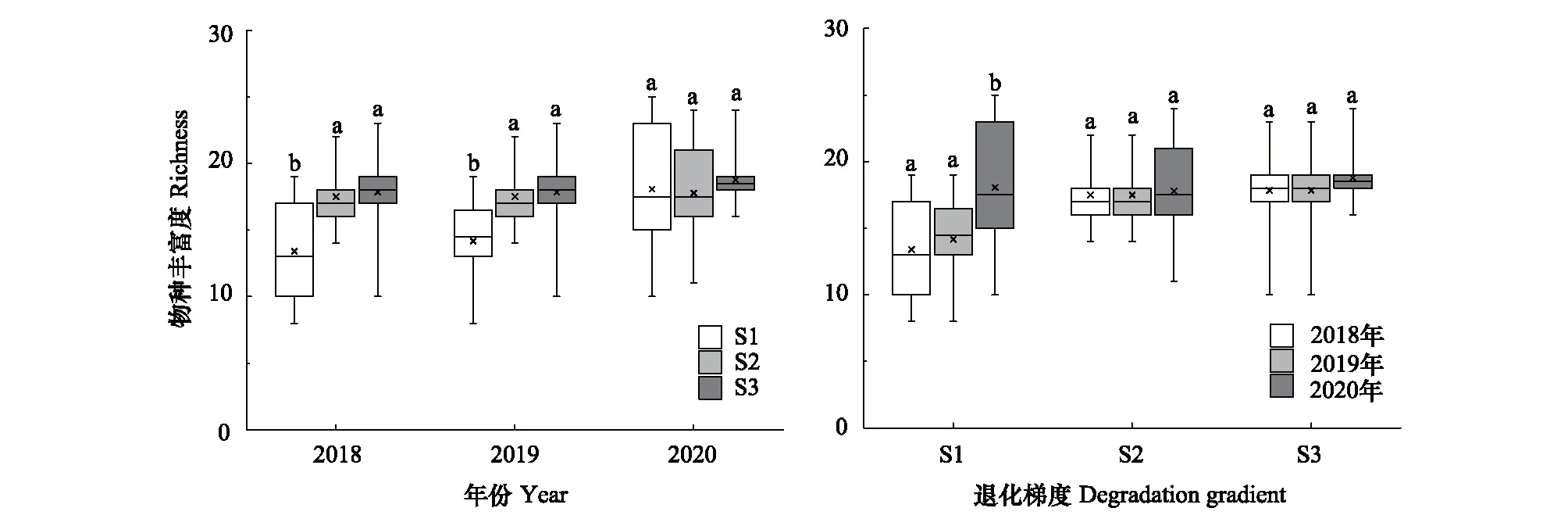
图2 2018—2020年退化梯度上草甸植物物种丰富度变化特征Fig.2 Changes of species richness among different degradation gradients from 2018 to 2020 S1:严重退化Severe degradation;S2: 中度退化Moderate degradation;S3: 轻度退化Slight degradation;不同小写字母表示显著差异
ANOSIM分析表明,刈割前(2018年),草甸植物群落组成在三个退化梯度间有显著差异;刈割后,植物群落组成在退化梯度间的差异性降低(表2)。从单个退化梯度看,相较于S2和S3,刈割后S1样地的植物群落组成变化最大(表3)。

表2 2018—2020年的三个退化梯度之间的植物群落差异系数

表3 2018—2020年各退化梯度植物群落差异系数
2.2 草甸植物群落复杂度和组织程度对刈割的响应
刈割前(2018年),草甸植物群落网络连通性(即植物群落复杂度)排序为:S1>S2>S3。植物群落网络模块化指数 (即植物群落组织程度)排序为:S2>S3>S1。刈割后,S1、S2和S3的群落连通性均先下降后增加(图3、图4)。群落组织程度的变化与群落复杂度相反,呈先增加后下降的特征(图4)。
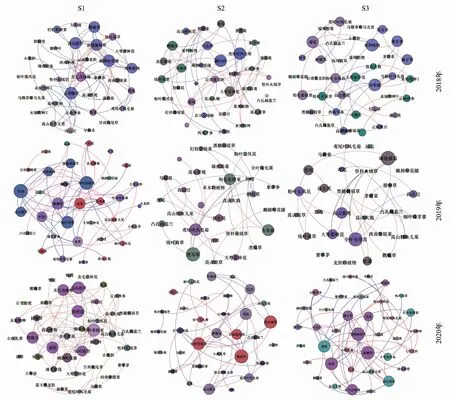
图3 2018—2020年三个退化梯度上的草甸植物群落网络Fig.3 Networks of plant communities for each degradation gradient from 2018 to 2020蓝线表示负相关关系,红线表示正相关关系
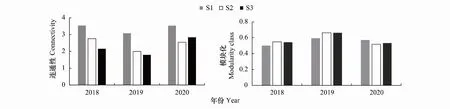
图4 三个退化梯度植物群落连通性和模块化水平变化特征Fig.4 Changes in connectivity and modularity class of plant communities at different degradation gradients
2.3 草甸植物群落关键种对刈割的响应
2018—2020年,S1样地的关键种变化为:云南棘豆 (Oxytropisyunnanensis)→云南棘豆→钉柱委陵菜(Potentillasaundersiana);S2样地的关键种变化为:黑穗画眉草(Eragrostisnigra)→牡蒿(Artemisiajaponica)→短颖披碱草(Elymusburchan-buddae)。S3样地的关键种变化为:西南委陵菜(Potentillafulgens)→坚杆火绒草(Leontopodiumfranchetii)→短颖披碱草(图3)。
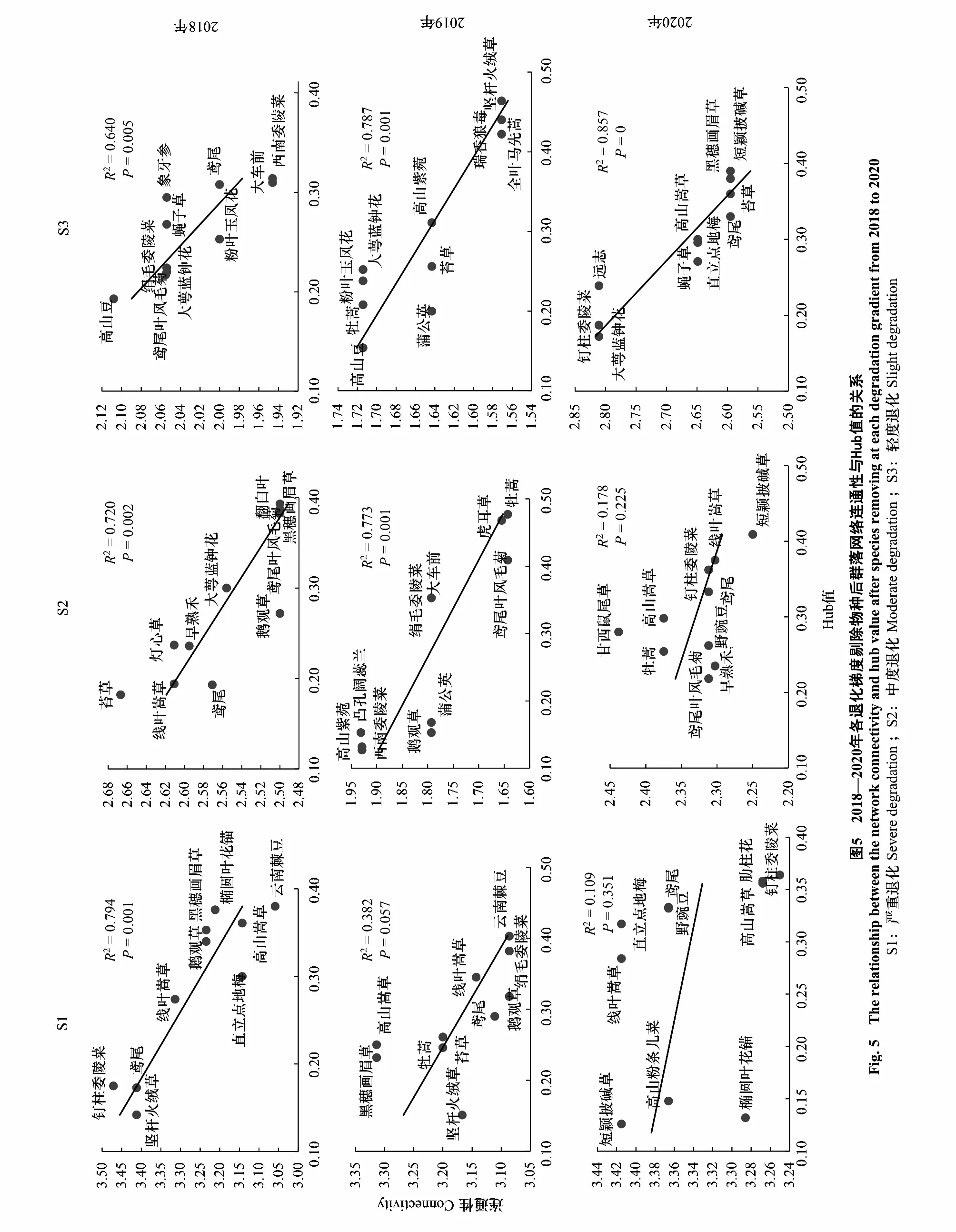
将包括关键种在内的Hub值排序前10位的物种逐一剔除发现,这些物种剔除对S1、S2和S3样地中植物群落连通性均造成了负面影响,且被剔除物种的Hub值越大,相应的群落连通性越差。特别地,将Hub值最高的物种(即关键种)剔除后,草甸植物群落的连通性最差(图5)。
3 讨论
青藏高原的高寒草甸植物物种丰富度会随退化程度加剧而下降[36]。本研究中,刈割前重度退化草甸的物种丰富度明显低于中度退化和轻度退化草甸,其与现有研究结论一致。但刈割后,重度退化草甸的植物物种数增加,其他两个退化梯度的物种数无显著变化,导致刈割后期三个退化梯度间物种丰富度没有差异。不同土壤养分状况下植物种间竞争强度的变化可能是导致该现象的原因。研究表明,土壤养分供给对高寒地区植物物种多样性维持有重要作用[37]。土壤养分供给充足时(如健康高寒草甸),草甸植物群落中物种数较多,且植物种间关系复杂、稳定[38],因此物种丰富度对干扰表现出较强的抵抗能力。土壤养分供给不足会加剧植物种间竞争强度,土壤养分争夺中占优的物种会压缩其他物种的生存空间,最终导致群落中的物种数下降[39—40]。当外界干扰打破此优势格局,在土壤养分竞争中不占优的物种可通过拓展地上空间以占据更大的生态位[41],因此群落中的物种数可能会出现反弹。高寒草甸土壤养分随退化程度增加而下降[42],由此推论,草甸植物对土壤养分的竞争强度也会随之加剧,某些竞争力强的物种(如狼毒等耐旱、耐贫瘠的物种)往往在退化草甸中形成毒杂草的单优群落[43]。本研究表明,刈割减少退化草甸中优势物种的优势度时,群落的物种丰富度有逐步增加的趋势。此外,种间竞争可能是群落结构形成过程中最基本的内在驱动因素[44—45],物种数量变化可对群落的种间竞争格局产生影响[44,46]。本研究中,刈割后重度退化样地的物种数明显增加,导致该梯度植物群落的物种组成变化大于其他退化梯度。
植物群落物种丰富度与群落复杂度有很高的相关性[47],但根据植物群落构建的生态过滤机制,在生存环境相对良好的中度退化或轻度退化情境下,由于竞争过程缓慢以及短暂的竞争排除现象,植物群落的物种数较多,但群落复杂度不一定最高[48]。本研究中,刈割前重度退化草甸植物群落物种丰富度低,其群落复杂度与其他两个梯度的草甸没有显著差别,正印证了这个规律。
关键种维持生态系统的结构,一旦关键种丧失,系统结构将发生彻底变化[49]。从这个基本认识出发,本研究采用物种剔除的方式证明,在理论层面通过网络分析识别关键种具有较高的可信度:将关键种(Hub值最高的物种)剔除后,群落网络的连通性最低。功能性状是解释物种生态功能的重要视角[50]。在高寒地区,具有保水、土壤养分维持、耐干扰等功能性状的物种是潜在的关键种[13]。本研究中,重度退化草甸的关键种为耐牧、耐干扰的委陵菜属植物,其耐牧和耐干扰特征可以延缓或阻止草甸进一步退化[51—52],另外是植株低矮、匍匐贴地生长、可固氮的豆科植物,它们可以帮助其他物种在重度退化环境中生存[53]。在中度和轻度退化样地中,关键种具有直立高大等有利于占据更多生态位的形态特征,它们在光的竞争中处于优势[39,54],这可能利于维持群落种间竞争的平衡关系。从功能群(根据牧草质量划分)角度看,本研究中重度退化草甸植物关键种均为杂类草,其他两个退化草甸植物关键种在杂类草和禾本科之间转变。
4 结论
本研究表明滇西北地区重度退化草甸的植物物种丰富度、物种组成和群落复杂度对刈割干扰的响应敏感性高于中度退化和轻度退化,这表明本研究的研究假设不成立,即:滇西北地区高寒草甸植物群落结构对刈割干扰的响应敏感性可能随退化程度增加而上升。但各退化梯度上群落关键种的变化则可能表明了相反的结论。
猜你喜欢
绿色天府(2022年9期)2022-11-19
农业工程学报(2022年12期)2022-09-09
流行色(2022年7期)2022-08-31
世界热带农业信息(2022年8期)2022-07-19
风景园林(2022年7期)2022-07-15
贵州畜牧兽医(2022年3期)2022-06-28
林业调查规划(2022年2期)2022-05-07
海外文摘·文学版(2021年3期)2021-05-19
西部资源(2018年3期)2018-11-06
延河·绿色文学(2016年8期)2016-05-14
