
荒漠草原向灌丛转变过程两种优势植物定植土壤水分阈值特征
2022-10-27 03:14张振杰王红梅
生态学报 2022年19期
张振杰,于 露,王红梅,2,*
1 宁夏大学农学院,银川 750021 2 宁夏草牧业工程技术研究中心,银川 750021
草原灌丛化是指木本植物密度、盖度和生物量增加导致以草地为主的生态系统在景观和区域尺度上发生重要变化的现象[1],目前,灌丛化在北美洲、亚洲、非洲、地中海、澳大利亚以及北极地区均有发生,被认为是气候变化和人为驱动的生态系统稳态转变过程[2—3]。虽有研究表明灌丛入侵会促进初级生产力、养分循环和土壤有机质积累[4],但其对草本竞争优势所引起草本植物生物量和生物多样性减少,土壤理化性质变化,水土流失加剧及荒漠化等一系列草原生态系统结构、功能弱化等问题[5—8],已经成为干旱和半干旱地区牧民生计和草原生态系统功能发挥的主要威胁[4],特别是一些干旱和半干旱地区人为植被恢复中大量引入灌木的行为实则会造成该地区草原灌丛化加速,改变草原植被更新趋势和过程[1]。
种子萌发和幼苗定植作为植被更新过程中的瓶颈阶段[9—10],对干旱区域内物种优势度、植被更新方向及速率具有调控作用[11—13]。但其对环境变化,尤其是降雨与土壤水分变异高度敏感[14—15]。如有研究表明格兰马草(Boutelouaeriopoda)、冰草(Agropyroncristatum)等草原植物的萌发率和出苗率均随降雨量增加而提高[1,16],幼苗存活率与干旱呈负相关[17],幼苗的生理、形态和解剖学属性受干旱胁迫影响显著[18]。此外,在降水模式改变所引起土壤水分变化直接影响草地潜在生物量的同时,植被类型转变对土壤水分的间接影响,即由于植被选择不当或植被转变(例如灌木取代温带半干旱草原)引起的因植被蒸腾造成的生态干旱也会对种子萌发、幼苗定植产生影响,从而改变植物种间关系,加剧资源竞争,调控植被更新[19—20]。如宁夏东部荒漠草原随着灌丛引入密度和年限的增加,引起土壤水分消耗的同时加剧了与草本植物的竞争,导致草本生物量、丰富度锐减,多年生草本土壤种子库密度萎缩、萌发更新受限[1,21]。而在奇瓦瓦区域的研究也表明,灌木入侵改变了土壤水分、理化性质以及植被繁殖体,在资源竞争中处于优势地位,限制了格兰马草属多年生草本植物更新,从而加快了土壤侵蚀速度与资源消耗[22]。并且大多数灌木种根系可进行横向和竖向延伸,不断增加与周围土壤的接触面积,入侵草本生存空间,尽可能攫取草本植物原本获取的土壤资源(以土壤水分为主)进行植株自身的生长繁殖活动。未来,由全球降水模式改变与灌丛入侵的协同效应所引起土壤的水分变化将对荒漠草原-灌丛交错带种子萌发和幼苗定植造成进一步影响,改变植被种间关系及资源利用模式,影响该地区植被更新方向和速度[23—24]。因此量化干旱胁迫下灌木与荒漠草原优势草本幼苗定植特征对于理解该地区草原灌丛化具有积极意义。
自20世纪七八十年代起,宁夏东部荒漠草原地区广泛应用旱生柠条锦鸡儿(Caraganakorshinskii)栽培种用于植被重建,在增加地上生物量的同时,引发了多年生草本如蒙古冰草(Agropyronmongolicum)等的退化、土壤旱化等一系列问题,柠条锦鸡儿逐渐成为优势种,使得荒漠草原正在向灌丛人为转变,形成大量的草原-灌丛土镶嵌体[24—25]。基于此,本研究选取典型草地-灌丛地镶嵌体区域,在沙芦草和柠条锦鸡儿种子萌发特性研究基础上,开展干旱胁迫下两种优势植物种子萌发后的定植试验,观测种子幼苗定植时期的生长状况以及土壤水势、水分特征,了解原生多年生禾草和引入灌丛定植阶段的土壤水分需求特征,确定其土壤水分阈值,探究全球气候变化背景下草原灌丛化过程中两种优势植物定植及其更新竞争机制,为荒漠草原人工植被恢复的演替趋势提供定量依据。
1材料与方法1.1研究区概况及样地设置
图1 研究区及试验采样点分布图Fig.1 The study area and the location of sampling sites
研究区位于宁夏回族自治区东部荒漠草原地区,地理坐标为37°04′—38°10′N,106°03′—107°04′E(图1),海拔为1450 m左右,属典型温带大陆性气候。年均气温7.7℃,无霜期162 d左右,年降水量250—350 mm,蒸发量2131.8 mm,全年降水量的60%—70%发生在7—9月份。本研究区主要土壤类型为灰钙土,土壤结构松散,土地贫瘠。地带性植被为荒漠草原,主要分布有蒙古冰草(Agropyronmongolicum)、短花针茅(Stipabreviflora)、糙隐子草(Cleistogenessquarrosa)、赖草(Leymussecalinus)、白草(Pennisetumflaccidum)等多年生植物和狗尾草(Setariaviridis)、猪毛菜(Salsolacollina)、猪毛蒿(Artemisiascoparia)等一年生植物,灌木、半灌木主要是柠条锦鸡儿(Caraganakorshinskii)和牛枝子(Lespedezapotaninii)。本试验在盐池县东北部的花马池镇四墩子村选取典型的草原灌丛镶嵌区域,从中确定荒漠草地区(Desert grassland,DG)、草地边缘(Grassland edge,GE)、灌丛边缘(Shrubland edge,SE)和柠条锦鸡儿灌丛区(Shrubland,SL)4个研究样地(图1,表1[25—26])。
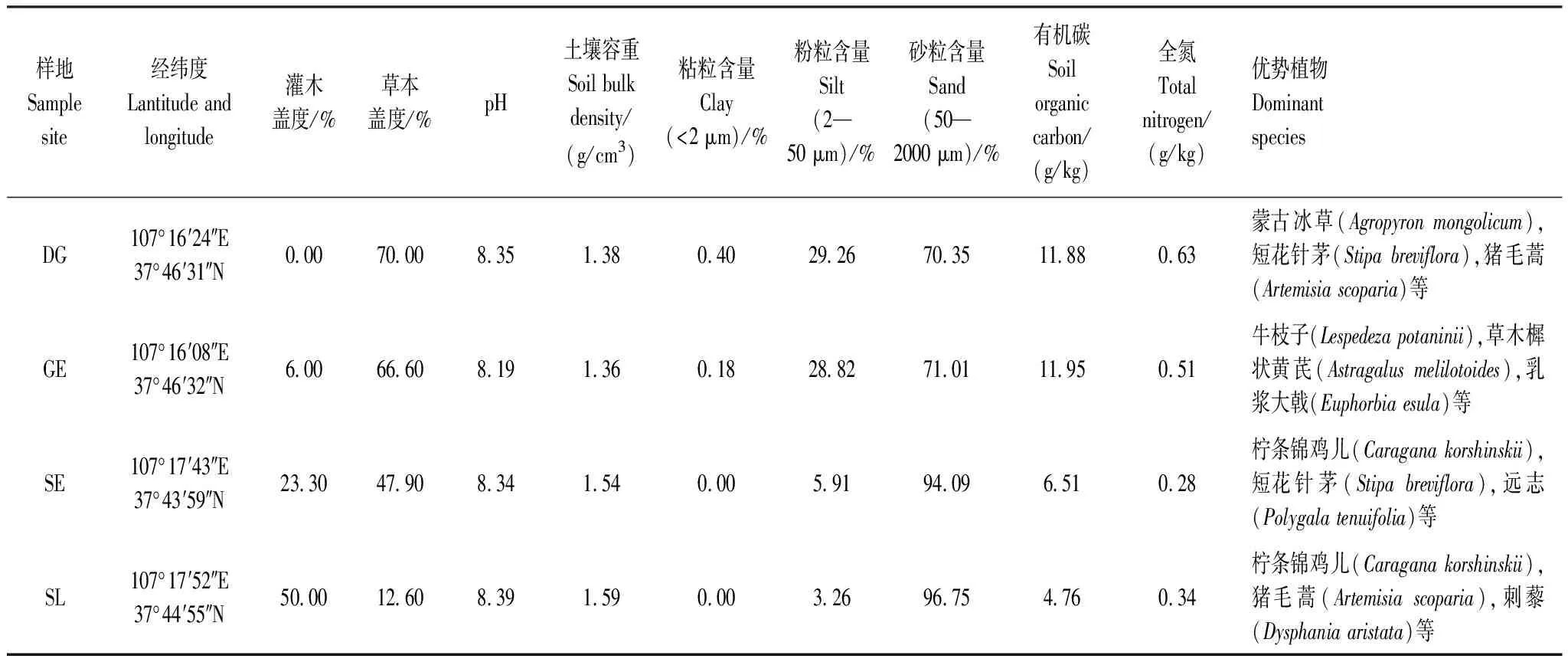
表1 样地概况
1.2 研究方法
1.2.1土壤水分及物理特性
在荒漠草地及草地边缘随机各设置6个5 m×5 m样区,灌丛(间距6 m,种植年限近30年)及灌丛边缘分别跨越带间、带内各设置3个10 m×10 m样区为本试验固定监测样区。(1)0—200 cm土壤水分:采用时域反射仪(Time Domain Reflectometry,TDR)对样地每个样区进行2个重复(每个样地共计12个重复)的土壤水分观测,通过不同方向(4次)调整TDR管读取数据并及时记录,每月2次,观测深度为0—200 cm,每20 cm为一层,共10层,观测时间为2019年4—11月。(2)0—20 cm土壤水分:使用环刀法采样,取样深度为0—20 cm,分为3层,分别为0—5,5—10,10—20 cm,取样后装入铝盒,以测定土壤质量含水量。
1.2.2土壤水势与土壤水分测定
采用0—10 cm的土壤含水率和土壤水势计算各样地表层土壤水分特征曲线,土壤含水率和土壤水势分别用烘干法和PSYPRO露点水势仪测定。于2019年9月在各样地内设置与上述一致的试验测定点,从7:00至17:00每隔1 h测定并取0—10 cm土样,及时记录土壤水势值,直接称其所取土壤样品鲜重。待全部测定结束后将所取土样带回,采用烘干法(105℃,24 h)测定土壤水分质量含水量。
1.2.3试验材料收集及处理
试验材料于2018年7—9月在宁夏盐池县四墩子基地荒漠草地试验区进行采集。供试的蒙古冰草种子由该试验区内野生蒙古冰草繁育基地采集,柠条锦鸡儿种子则通过野外随机收集方式获得,即选取健康柠条锦鸡儿植株后摘下已成熟荚果,特征详见表2[27]。采集的种子带回到实验室晾干后,人工除去种子内杂质和有虫蛀或失去生活力的种子并使用物理方法去除柠条锦鸡儿种皮,以避免其种皮过厚对本试验的影响,最后将其放置在4℃下贮藏供试验所用。同时为防止种子在试验中发霉,将其用0.1%HgCl2消毒10 s左右后用蒸馏水反复冲洗[1]。

表2 柠条锦鸡儿和沙芦草的种子特性
1.2.4种子破土及定植
在于露等[27]初步探明荒漠草原优势植物种子萌发水势阈值的基础上,进行幼苗定植试验,探索荒漠草原优势植物在灌丛转变各阶段幼苗定植水分阈值。筛取从4个样地所取得的定量土壤样品,并烘干以避免土壤中多余植物种子对试验所造成的影响。将各样地所取土壤各设置8个水势梯度,每个梯度3个重复,并据此将各样地所取土壤分为均等的24份,分别铺在花盆(花盆内径15 cm、高15 cm,塑料花盆底部预先填充5 cm厚、无种子的蛭石,铺设土样厚度为9—10 cm)中,4个样地,共铺设96个花盆。每个花盆分别种植柠条锦鸡儿种子50粒(播种深度为2—3 cm)。通过保持土壤其它性质不变的情况下,根据Michel等[28]配制方法,在相同样地土壤中加入浓度为2.9%(对照组CK)、8.2%、12.5%、17.5%、20%、21.1%、23%、23.8%(分别相当于0、-0.02、-0.1、-0.3、-0.6、-0.7、-0.9、-1.2 MPa的渗透势胁迫[27])且体积相同(350 mL)的PEG-6000聚乙二醇,改变土壤渗透势(溶质势)来建立土壤水势梯度。将试验样品准确称重后放置在适宜环境(光照长度为12 h,温度20—25℃)下。每隔2天采用恒重法向花盆内洒入蒸馏水,保持土壤渗透势不变,进行不同水势梯度、不同样地内两种植物破土及定植试验。PEG溶液的渗透梯度与土壤中的水势并不一致,采用不同浓度PEG溶液是为了通过改变土壤渗透势从而改变土壤水势建立不同水势梯度,采用恒重法是为保持土壤渗透势不变从而保持土壤水势基本恒定。因此定植试验开始后,及时记录每个PEG浓度下的土壤水势值,取3个重复的平均值作为该渗透势胁迫下的水势梯度值,并记录种子破土及幼苗死亡情况,50天后试验结束,测定幼苗叶绿素含量。蒙古冰草定植试验与之相同。
1.3 测试指标
1.3.1土壤水分特征曲线模拟
对各样地内土壤水势和体积含水量数据进行非线性曲线模拟并绘制土壤水分特征曲线。
1.3.2幼苗死亡率计算
式中,n和N分别为定植试验完成后存活的幼苗数和出苗总数。
1.4 数据分析
利用单因素方差分析(One-way ANOVA)中的最小显著差法(LSD)对PEG浓度模拟干旱胁迫梯度下定植的各项参应进行分析;使用S型曲线拟合渗透势和死亡率,确定幼苗死亡率对渗透势的响应曲线,并根据羊旻[29]的方法,令S型曲线的三阶导函数等于零,计算其加速增长阶段作为渗透势阈值范围。采用线性模型拟合渗透势和土壤水势,确定渗透势和土壤水势模型,利用非线性Log3P1模型拟合土壤水势和水分,确定土壤水分特征曲线;利用幼苗死亡率对渗透势的响应曲线、渗透势和土壤水势模型和土壤水分特征曲线推算定植土壤水分阈值。数据处理采用SPSS 13.0软件,采用Excel和Origin绘图并进行曲线拟合,求导函数及其他数值运算采用数学软件Maple完成。
2 结果
2.1 转变过程土壤水分动态变化
如图2可知,在荒漠草地向灌丛地人为转变过程中,0—20 cm土层土壤含水量显著低于0—200 cm土层,两者土壤水分含量均表现为荒漠草地>草地边缘>灌丛边缘>灌丛地(P<0.05),且荒漠草地显著高于灌丛地,分别高出4.6%和8.11%。除灌丛地0—200 cm土层没有明显的季节动态,各样地0—20 cm和0—200 cm土层土壤水分均呈现生长期降低和秋季降雨增加的季节波动。
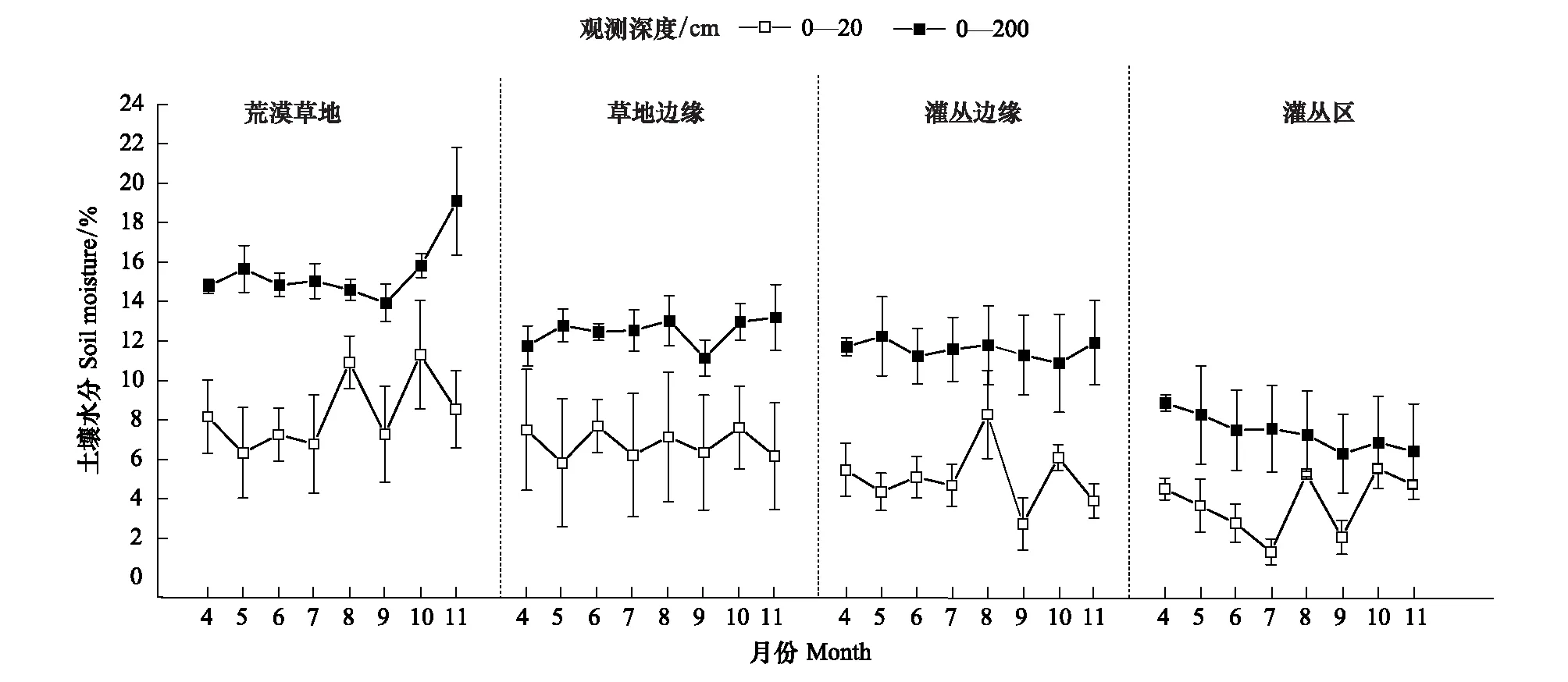
图2 各样土壤水分季节变化Fig.2 Seasonal dynamics of shallow soil moisture in each site
2.2 转变过程中干旱胁迫下植物定植叶绿素含量变化
由图3可知,蒙古冰草在草地边缘内叶绿素平均含量最大,为4.29 mg/g,对渗透胁迫加深响应也最为敏感,最值间差值为11.68 mg/g;在灌丛地内叶绿素含量波动区间为2.35—4.75 mg/g,波动最小且一直处于低值。柠条锦鸡儿同样在草地边缘内叶绿素平均含量最高,为24.46 mg/g;对渗透胁迫加深响应也最敏感,最值间差值为15.95 mg/g。各样地转变过程中,蒙古冰草和柠条锦鸡儿叶绿素含量均先上升,在渗透势为-0.02 MPa时达到最大值,两者分别平均为7.17 mg/g和29.40 mg/g,而后随渗透势胁迫加剧呈逐步下降趋势,至-1.2 MPa时含量最少,两者分别平均为1.67 mg/g和16.08 mg/g,且柠条锦鸡儿叶绿素含量显著高于蒙古冰草(P<0.05)。
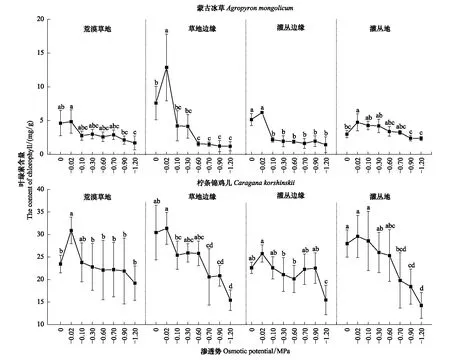
图3 荒漠草原-灌丛人为转变过程中柠条锦鸡儿和蒙古冰草叶绿素含量的响应Fig.3 Response of Chlophyll content of Caragana korshinskii and Agropyron mongolicum to different osmotic potential in desert grassland-shrubland state transition不同字母表示同一样地内不同渗透势间叶绿素的差异显著(P<0.05)
2.3 转变过程中干旱胁迫下植物定植过程死亡率变化
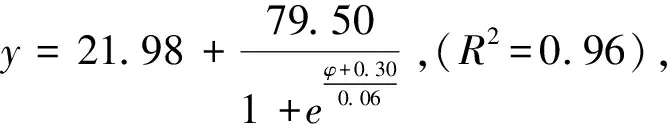
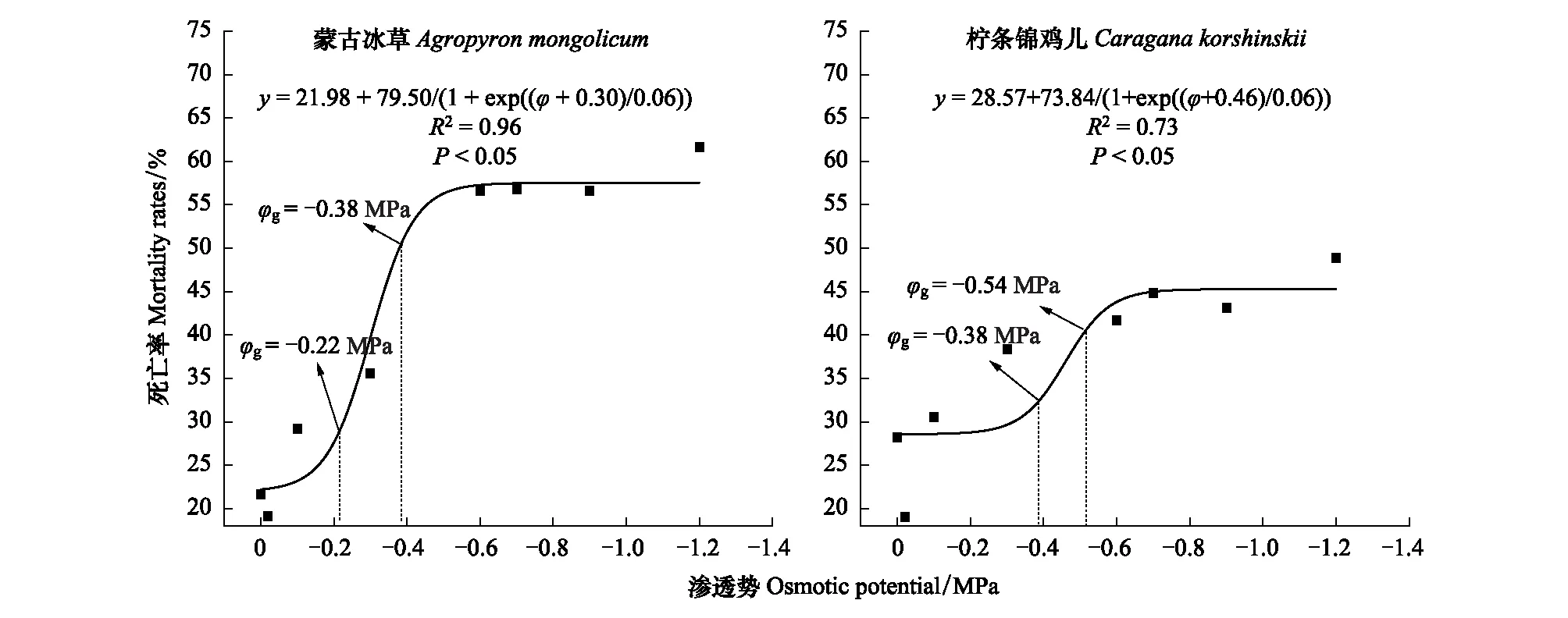
图4 柠条锦鸡儿和蒙古冰草在荒漠草地的死亡率受渗透势的影响 Fig.4 The impacts of osmotic potentials on mortality rate of Caragana korshinskii and Agropyron mongolicum in the Desert grassland
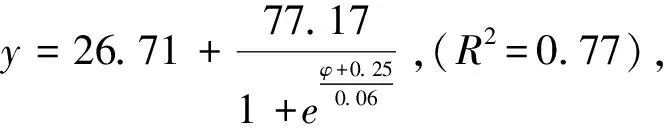
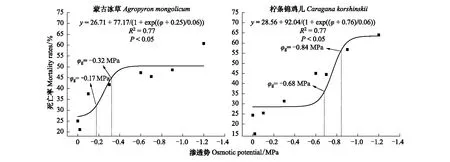
图5 柠条锦鸡儿和蒙古冰草在草地边缘的死亡率受渗透势的影响 Fig.5 The impacts of osmotic potentials on mortality rate of Caragana korshinskii and Agropyron mongolicum in the Grassland edge
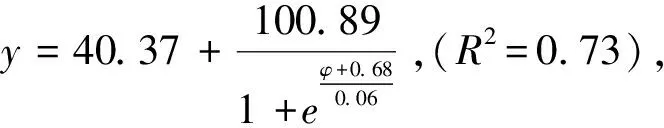
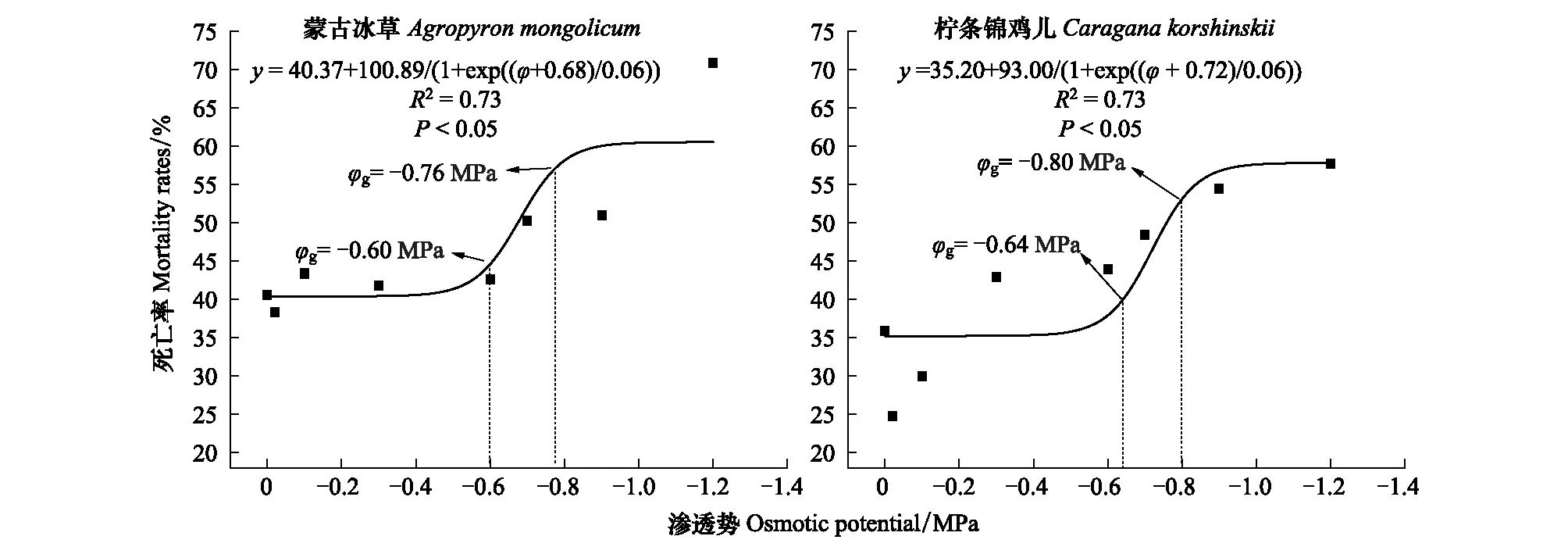
图6 柠条锦鸡儿和蒙古冰草在灌丛边缘的死亡率受渗透势的影响Fig.6 The impacts of osmotic potentials on mortality rate of Caragana korshinskii and Agropyron mongolicum in the Shrubland edge
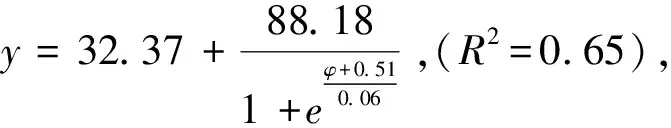
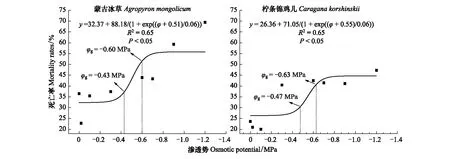
图7 柠条锦鸡儿和蒙古冰草在灌丛地的死亡率受渗透势的影响Fig.7 The impacts of osmotic potentials on mortality rate of Caragana korshinskii and Agropyron mongolicum in the Shrubland
2.4 渗透势与各转变样地土壤水势关系模拟
由图8可知,通过线性拟合模型进行模拟后发现,盆栽试验中用于干旱胁迫的溶液渗透势与各样地土壤水势关系分别如下:
荒漠草地:y1=-6.91+2.26x1;草地边缘:y2=-7.15+3.91x2
灌丛边缘:y3=-7.72+4.21x3;灌丛地:y4=-8.36+3.87x4
蒙古冰草在荒漠草地、草地边缘、灌丛边缘和灌丛地定植土壤水势阈值分别为:-6.79—-7.14、-7.81—-8.40、-10.24—-10.91和-10.02—-10.68 kPa,柠条锦鸡儿在四个样地定植土壤水势阈值分别为:-7.14—-7.51、-9.80—-10.43、-10.41—-11.08和-10.17—-10.79 kPa。
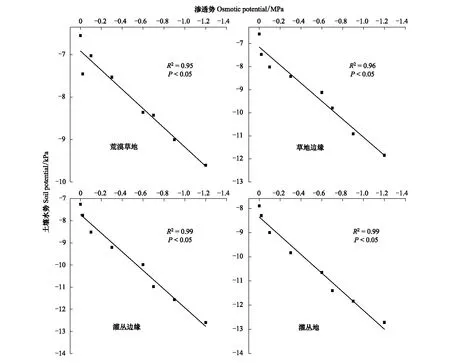
图8 渗透势和土壤水势模型Fig.8 The model of osmosis potential and soil potential
2.5 转变样地土壤水分特征曲线模拟
由图9可知,通过非线性Log3P1拟合模型进行拟合后发现,各样地土壤水势与水分关系分别如下:
荒漠草地:y1=-19.43+5.40ln(x1-1.57);草地边缘:y2=-12.81+2.47ln(x2-1.39)
灌丛边缘:y3=-17.46+8.01ln(x3-1.21);灌丛地:y4= -10.26+2.86ln(x4-1.78)
蒙古冰草在荒漠草地、草地边缘、灌丛边缘和灌丛地定植土壤水分阈值分别为10.23%—10.84%、7.35%—8.95%、3.47%—3.67%,2.64%—2.87%。而柠条锦鸡儿在四个样地内定植土壤水分阈值分别为9.67%—10.23%、4.01%—4.76%、3.43%—3.62%和2.61%—2.81%。
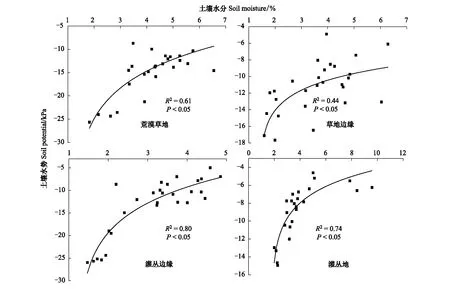
图9 土壤水分和土壤水势关系模型Fig.9 The model of soil moisture and soil potential
3 讨论
草原灌丛化作为全球面临的严峻生态问题之一,通过改变草地生态系统结构及功能,进而影响生态水文过程,造成土壤旱化[30]。虽然有研究指出丘陵地区灌木林盖度明显增加,减少地面接收太阳辐射,可降低土壤水分蒸发量,出现灌木林地表层土壤水分高于草滩草坡的现象[31—32],但大多数研究结果表明,随着草原灌丛化的加剧和灌丛生长年限的增加,地上植被群落结构被改变,木本植物逐渐取代多年生草本,土壤理化性质发生改变,土壤旱化加重,特别是深层次土壤水分消耗加剧[33]。在本研究结果中,随灌丛盖度从0%急剧增加到50%,土壤表层水分与深层土壤水分均呈下降趋势。这可能由于灌木高生物量需要大量水分支持,且其拥有较深的主根和发达的侧根,能够同时吸收深层土壤水分和浅层土壤水分,从而加剧灌丛区域内土壤水分蒸腾,造成深层土壤水分消耗加剧[1]。此外,研究发现随灌丛转变,土壤容重、粒径和土壤孔隙度显著改变,粘粒和粉粒减少,而沙粒增加[26];这可能是因为灌木根系在长期延伸过程中会与周围土壤进行相互作用,破坏土壤粒径和孔隙度。而土壤中形成的相对互联的大孔隙,可以改变土壤渗透性和水力传导特性,将部分水分转变成为“优势流”,迅速渗入并储存在土壤深处以适应干旱环境来维持其生物量[34—35]。荒漠草地向灌丛地转变可能会通过“沃岛效应”促进土壤碳(C)和氮(N)的积累[36],但在此样地中,灌丛地有机碳、全氮含量均显著低于荒漠草地,且灌下养分积聚不明显,可能是灌木对土壤养分库的影响是由于C、N累积和流失途径的相对重要性不同所致,即在转变过程中促进土壤C和N积累的因素部分或全部被促进增加的损失因素(呼吸作用、硝化作用和反硝化作用)抵消[37]。这些利己的资源利用方式,增加了草原群落尺度上的土壤资源空间异质性,加速草本的退化[1],增加了土壤空斑面积[38],进而加速了表层和深层土壤水分的蒸发和消耗。
幼苗定植期是植被更新的关键环节,对环境变化,尤其是干旱胁迫极为敏感。因此,在灌丛化所造成土壤日益旱化的情况下,植物幼苗定植的成功与否决定着该地区未来植被更新的走向[39]。本研究结果表明:在轻微干旱条件下,蒙古冰草和柠条锦鸡儿的死亡率均处于最低值,说明轻度干旱可以促进其定植成功[40—41],可能是因为荒漠植被在发育过程中已经形成一定程度的干旱适应机制。但随着干旱胁迫进一步加剧,两者死亡率均呈现上升趋势,而柠条锦鸡儿死亡率波动较小,且在不同干旱梯度下的变化均小于蒙古冰草,在极端干旱情况下的死亡率也低于蒙古冰草,说明在土壤旱化逐步加剧和水分变异较大情况下,柠条锦鸡儿的幼苗定植更具竞争性。叶绿素作为植物光合作用中的主要色素,参与吸收与转化太阳能以确保光合作用顺利完成,与植物根系之间联系密切的同时对干旱条件亦十分敏感[42]。Ammar等[43]研究发现,随干旱胁迫程度加剧,叶绿素呈先增加后递减趋势。本试验结果也表明随着干旱程度加深,柠条锦鸡儿和蒙古冰草的叶绿素含量均先升高后下降,这是因为干旱胁迫加剧减弱了两者根系吸收外界能源的能力,进一步使植株体内酶活性降解失活,继而降低了叶绿素含量,与前人研究相近[44]。并且随着干旱程度的加深,柠条锦鸡儿叶绿素含量的降低比例(45.32%)显著小于沙芦草叶绿素含量的降低比例(76.72%),说明沙芦草的叶绿素含量对于干旱胁迫更为敏感。结合土壤水分特征曲线,就两种植物的死亡率和各样地的土壤水分变化总结发现,随着干旱梯度加大,土壤水分含量逐渐减少,而死亡率随之显著增加,且在转变过程中两者死亡率均有所升高,以蒙古冰草最为显著;依据S型曲线所得出的阈值也发现:柠条锦鸡儿在各样地幼苗定植中所需土壤水分阈值均低于蒙古冰草,因此在长期干旱或水分匮乏的环境中柠条锦鸡儿幼苗更容易存活并定植。并且随荒漠草地向灌丛转变,两种植物幼苗定植所需土壤水势阈值变化不明显但水分阈值变化显著,这与该区域土壤颗粒组成、团粒结构、容重等土壤物理特性的改变相关[26]。
尽管诸多原因造成灌丛化现象,但全球气候变化导致的降雨模式改变,为耐旱、耐贫瘠的灌木植物在干旱、半干旱地区提供了扩张机会[45],对其群落结构和生态系统有着深远影响,即植被群落结构和空间分布的巨大转变[46],短期使生物量发生变化,长期则影响植被构成[47];在本研究中,就总体水分含量(0—200 cm)而言,荒漠草地春季土壤水分逐渐升高;当7、8月降水量最多时,与之对应的各样地内夏季土壤水分含量却最低;进入秋季后降雨使得各样地土壤水分得到补充继而升高,说明季节变化下土壤水分与所处环境中降水变化表现即使有所延迟,但大致同步。但在气候和植被共同调控下,灌丛地消耗了大量土壤深层水分,即使夏季降水较充沛,深层土壤水分也是通过再分配注入旱化的深层次土壤用以补充消耗[48],导致秋季土壤水分增加不显著,且春季返潮不明显。同时夏季植被生长需水较强,除地表蒸发失水外,地上植被维持自身所需水分以及植物蒸腾作用等也大量消耗土壤水分[49]。随着水分不断消耗,干旱胁迫将进一步加剧。在本文中,随干旱胁迫加剧,柠条锦鸡儿幼苗的死亡率逐步低于蒙古冰草低,且变异较小,叶绿素含量下降幅度也较低,总体水分阈值亦较低,因此在长期干旱下更容易存活并定植。另外,Heras等[50]研究推测奇瓦瓦地区灌丛扩张可能是由于石炭酸灌木(L.tridentata)在1960年干旱时期中保有抗旱竞争优势的结论也与本研究一致。上述研究结果反映了植被与气候之间存在反馈机制,在灌丛引入的条件下,气候与植被的共同调控作用可能会导致生态干旱[23],由于灌木和多年生草本植物在定植期间对于干旱胁迫的响应差异,将导致多年生草本植物在定植过程中处于劣势,进一步加剧灌丛扩张和土壤退化。
4 结论
荒漠草原-灌丛近30年转变过程中土壤水分呈下降趋势,旱化加剧。柠条锦鸡儿和蒙古冰草幼苗定植过程中叶绿素含量随干旱胁迫程度加深呈现先略微增加后降低趋势,死亡率逐渐上升。在转变过程中柠条锦鸡儿和蒙古冰草定植平均土壤水分阈值分别为4.93%—5.23%和5.92%—6.50%,且柠锦鸡儿幼苗在各转变样地所需土壤水分阈值均低于蒙古冰草,较蒙古冰草(幼苗)更适应土壤旱化。
猜你喜欢
农业工程学报(2022年10期)2022-08-22
小哥白尼(趣味科学)(2022年5期)2022-08-15
云南大学学报(自然科学版)(2022年4期)2022-08-03
农业工程学报(2022年5期)2022-06-22
水土保持学报(2022年3期)2022-05-26
当代陕西(2019年7期)2019-04-25
湖南饲料(2018年2期)2018-12-05
扬子江(2018年5期)2018-09-26
儿童故事画报·智力大王(2016年7期)2017-02-08
文史春秋(2016年3期)2016-12-01
