
江苏滨海湿地芦苇和互花米草光合特性对模拟增温的响应
2022-10-27 03:13江星浩谢柳娟叶思源裴理鑫赵俐红
生态学报 2022年19期
江星浩,谢柳娟,叶思源,*,周 攀,裴理鑫,陈 皓,赵俐红
1 山东科技大学地球科学与工程学院,青岛 266590 2 青岛海洋地质研究所,中国地质调查局滨海湿地生物地质重点实验室,青岛 266071 3 青岛海洋科学与技术试点国家实验室,海洋地质过程与环境功能实验室,青岛 266061 4 江苏省测绘工程院,南京 210003
全球变暖是全球气候变化的主要表现之一。根据政府间气候变化专门委员会(Intergovernmental Panel on Climate Change,IPCC)第五次报告,人为温室气体的排放已成为自20世纪中期以来气候变暖的主要原因,使得全球气温相较于工业革命之前升高了1℃左右[1—3]。据中国气象局(CMA)发布的《中国气候变化蓝皮书(2020)》和国家海洋局(SOA)发布的《中国海平面公报(2019)》,1980—2019年,中国沿海海表温度和气温均呈上升趋势,上升速率分别为0.023℃/a和0.038℃/a。2019年,中国沿海海表温度和气温较常年均高0.9℃,比2018年分别高0.4℃和0.5℃,均为近40年来最高。
湿地是地球上具有重要环境功能的生态系统。科学家们普遍认为,湿地具有较低的有机质分解速率和较高的生产力,是重要的碳汇资源[4]。特别是滨海湿地,由于海水可持续提供硫酸根离子,硫酸盐还原菌会与产甲烷菌竞争代谢所需的底质,从而有效抑制了CH4的产生。因此,滨海湿地在应对全球变暖方面有着更大的潜力,在保护生物多样性、调节气候变化等方面起着重要的功能[5—6],是应对全球气候变化效应的关键生态系统,在全球碳循环中占据重要位置[7—8]。
光合作用是植物重要的生理过程,是生态系统碳循环和能量流动必不可少的一个环节[9]。植物的光合特性直接影响植物的生存、竞争能力、生产力以及碳汇能力,而温度是影响植物光合作用的主要环境因素[10]。通常,温度通过提高参与植物光合作用的酶的活性和含量来提高植物的光合作用,增加植物对养分的吸收[11]。但由于植物本身特性,加上不同地区的地理环境因素,增温对植物光合特性的影响不尽相同,已有大量研究证明了这一观点,具体表现为增温使植物光合作用增加[12]、下降[13]或无变化[14]。相关研究主要进展和争议可总结为以下几点:(1)不同的物种对增温有不同的响应。C4植物生长的适宜温度一般要高于C3植物[15],前者植物的光合作用在一定的温度范围内会随着温度的升高而提升,而后者植物在同样的条件下可能会超出植物生长的适宜温度,光合作用受到抑制[16]。例如在相同增温条件下C3植物本氏针茅光合作用无显著变化,C4植物白羊草光合能力提升[17]。(2)植物的生长环境对增温有不同的响应。实验室条件下光合速率随着温度的升高而提升,但在田间原位实验却得出相反的结果[18]。(3)植物气孔关闭影响光合作用的强度[19],温度过高,植物气孔关闭,气孔导度和蒸腾速率下降,气体交换能力减弱,降低植物的光合作用[20]。(4)植物对增温响应有一定的适应范围[21]。植物的生长温度在最低温度和最适温度之间时,植物的光合速率随着温度升高而升高,当环境温度超过最适温度时,继续升温就会抑制植物的光合作用[17]。不同物种在生态系统中因为不同的生态位和对外界环境的适应能力不同,必然导致对环境变化的响应程度不同,最终将改变植物种间关系和群落内部结构[22]。因此,研究生态系统对增温的响应已经成为全球变化研究的热点之一[23]。
模拟植被在气候变暖影响下的响应实验正日趋完善。20世纪末期,人们广泛利用开顶箱(Open-top chamber,OTC)来模拟生态系统中植被在增温条件下的响应试验,并取得了许多成果[24—25]。国外科学家利用OTC装置在高纬度和高海拔地区进行了模拟增温实验。例如Havström等讨论了三种植被在高北极和亚北极地区模拟增温条件下的生长反应[26];Chapin等讨论了北极苔原对OTC实验和观测到的气候变化的响应[27];Laine等利用OTC装置讨论了增温对芬兰北方湿地温室气体通量交换的影响[28];我国科学家在崇明岛东滩湿地、青海湖湖滨湿地、青藏高原等多地设置了OTC观测样地,开展了一系列有关模拟增温的科学研究[29—31]。虽然植被形貌特征对增温响应的研究已有很多,但由于技术方法的限制,植被光合特性在增温条件响应下的研究却很少,有关滨海湿地入侵植物如何改变光合作用特征参数的微观研究更是鲜有报道,限制了植物对增温的响应机制的深刻理解。江苏盐城滨海湿地作为陆海相互作用的集中地带和我国首个湿地类型的世界遗产,在生态方面的作用更是重中之重,但由于外来物种互花米草的入侵,其光合特性不仅直接影响了本地物种芦苇的生长,更直接影响了湿地生态系统的碳汇功能。因此,开展江苏滨海湿地芦苇和互花米草光合特性对增温的响应及其响应机制的研究具有一定的科学意义。
本研究以江苏盐城滨海湿地芦苇(Phragmitesaustralis)和入侵植物互花米草(Spartinaalterniflora)为研究对象,采用野外开顶箱原位模拟增温,在九月份监测芦苇和互花米草的光合特性和环境因子的变化,探究植物光合特性在增温条件下的响应模式,预测在未来气候变暖情况下植物的竞争力和碳汇能力演变,为未来气候变化下该区域植被的管理和利用提供理论基础。
1 材料与方法
1.1 研究区域概况
研究区位于江苏省盐城市滨海湿地(图1),距盐城市中心约50 km,是暖温带季风气候与亚热带季风气候的过渡地带,是淤泥质海岸带湿地最具特色的自然景观。区域内有盐城国家级珍禽自然保护区和大丰麋鹿国家级自然保护区,有“东方湿地之都”的美誉。研究区自然条件较好,夏季高温多雨,冬季温暖湿润。年平均气温为13—16℃,受台风和梅雨的影响,雨季较长,雨量充沛,年降水量为1100 mm左右。日照充足,年平均日照时间时数为2300 h。季风性气候特征明显,四季分明,雨热同期。滨海湿地全球增温观测网络(Coastal-wetland Research On Warming Network,CROWN)盐城站主要包括两个观测站点:互花米草增温站(CROWN Ⅰ)(33°22′28.4″N,120°43′21.6″E)位于大丰区东北部,四卯酉河以北,距黄海海边不足五公里的互花米草生境内;芦苇增温站(CROWN Ⅱ)(33°36′55.8″N,120°32′22.4″E)位于射阳县东南部,新洋港河以南,盐城国家级珍禽自然保护区的芦苇生境内。
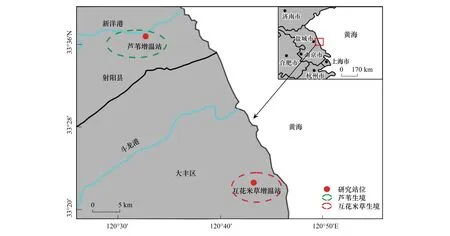
图1 研究区概况Fig.1 Overview of the study area
1.2 OTC实验
开顶箱(OTC)属于被动增温装置,具有不需要电力支持、建设和维护成本低、操作方便等优点[32—33]。玻璃设计可以降低内部空气流动,阻隔大气红外线辐射,内部热量不容易散失,是目前在国内外应用较为广泛的一种增温装置。2018年年初在江苏盐城新洋港芦苇生境和四卯酉互花米草生境分别选取地势均匀、植物生长状况较为一致的地块设置野外科学观测实验站,每个实验样地共设置6个开顶箱增温装置(OTC)和6个对照组。开顶箱呈八边形,铝合金框架构成,侧面为高透钢化玻璃(透光率>92%),边长1.07 m,高2.7 m,占地面积5.54 m2。开顶箱固定于钢制基板上,与土壤表面相距10 cm间隙,保证了装置内外水文通透性。紧邻增温装置设置相同地面形状的样地作为对照组。为避免科研过程中湿地土壤破坏和方便监测,增温站位和对照站位之间设有可供人员活动的木栈道。每块样地中的第一个增温站位建有一个环境因子在线监测系统(图2),该系统由太阳能电池板和铅酸蓄电池组成的供电系统供电,加上在线传输系统,能够实时监测气温、相对湿度、土壤电导率和土壤温度等关键环境因子。
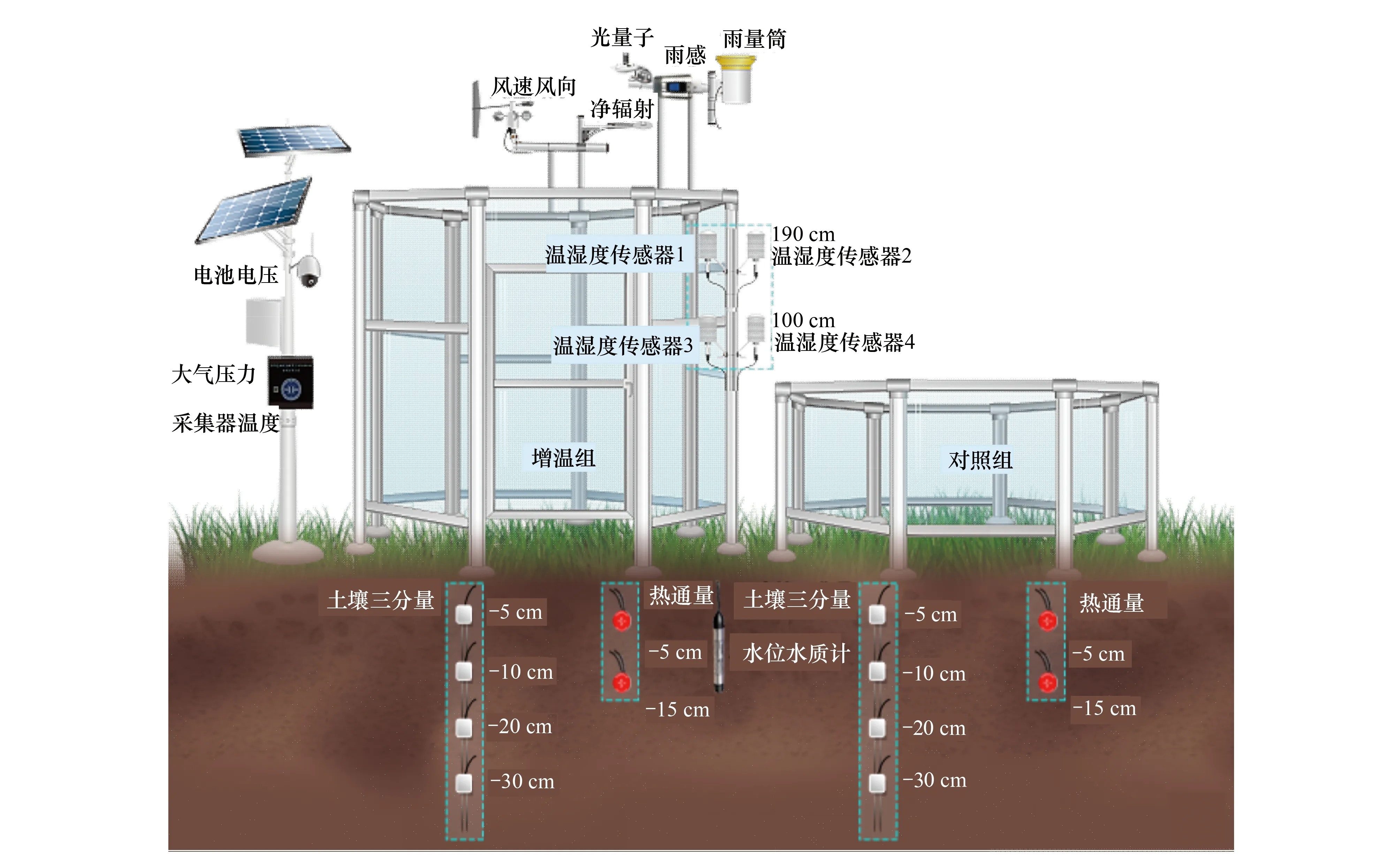
图2 增温观测站传感器布设(左图为开顶箱(OTC)增温样地,右图为对照样地)Fig.2 Sensor layouts of the observation station for warming experiments (the warming plot with an open-top chamber (OTC) (left panel); the control plot (right panel))
1.3 植物光合生理指标的测定
1.3.1瞬时气体交换参数的测定
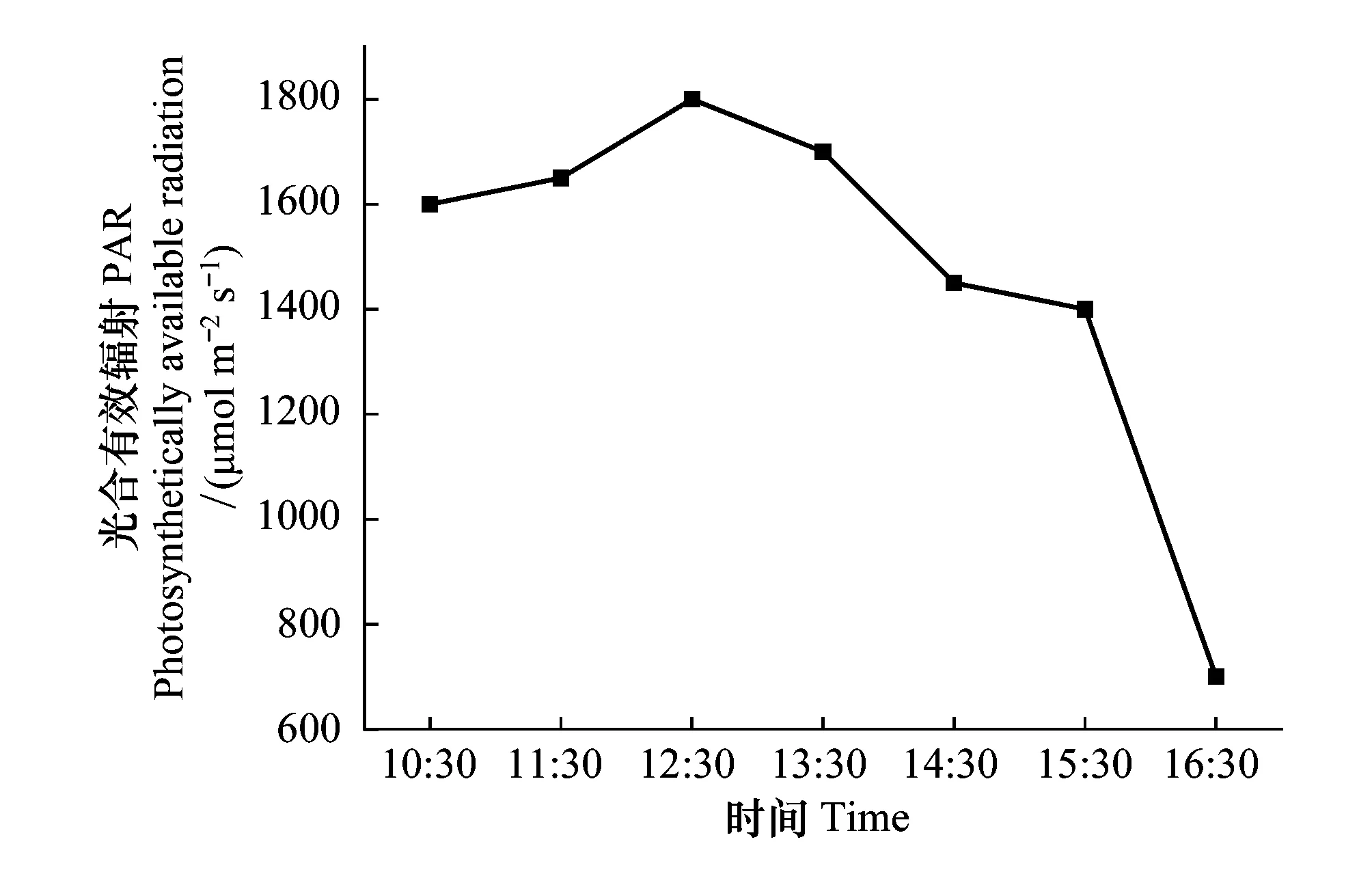
图3 研究时段的光合有效辐射(PAR)变化Fig.3 Changes of PAR during the study period
本研究以江苏盐城滨海湿地芦苇(Phragmitesaustralis)和入侵植物互花米草(Spartinaalterniflora)为研究对象,采用开顶箱模拟增温,监测植物的光合特性变化。2020年9月中旬,在6个OTC增温箱中随机选取3个增温组和相应的对照组作为实验样地。依次在每个增温组和对照组中随机选取长势良好且植物形貌特征基本一致的互花米草/芦苇1株,每株选定由顶端向下的第3个充分伸展的完整叶片为测试对象。利用Li- 6800便携式光合系统(LI-COR,USA)对选定叶片的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)和水分利用效率(WUE)等参数进行日调查动态测定。测定时段为10:00—16:30,每隔1 h测定一次,连续测定3次,每次测定采集5 min。泵流量采用仪器默认值500 μmol/s。大气CO2浓度控制在400 μmol/mol。为确保实验的准确性,将晴朗天气获取的各时段光强作为后期测定时人工光源控制的光合有效辐射(PAR)值来探讨植物在增温条件下的光合特性(图3)。
1.3.2光响应曲线的测定
相同测定条件下于9:00—11:00、14:00—16:00进行植物光响应曲线的测定,手动设置光合有效辐射(1800、1500、1400、1200、900、600、300、150、70、50、30、0 μmol m-2s-1)。在具体测定时,尽可能地将每一对(一条增温组和一条对照组光响应曲线为一对)光响应曲线测定时间安排在最接近的时段内完成(如各对平行样同时在上午或下午)。用光响应模型拟合所测量的数据可以获得植被的表观量子效率(AQY)、最大净光合速率(Pnmax)、光饱合点(LSP)、光补偿点(LCP)和暗呼吸速率(Rd)等重要的光合参数。目前常用的光响应模型有直角双曲线模型[34]、非直角双曲线模型[35]、指数模型[36]和直角双曲线修正模型[37]四种。本文将采用直角双曲线修正模型来拟合两种植物的光响应曲线。模型表达式如下:
式中,Pn(I)为净光合速率;I为光合有效辐射,α为I=0时光响应曲线的初始斜率即初始量子效率;β,γ是与光合有效辐射无关的系数;,Rd为植物的暗呼吸速率。
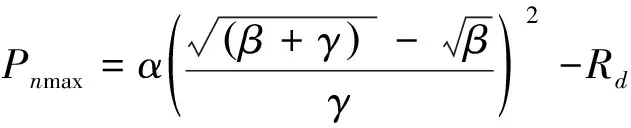
1.3.3数据处理与统计分析
用Excel 2010对实验原始数据进行整理、汇总。用SPSS 23.0软件对光合速率与主要影响因素进行相关性分析,统计显著性水平为P<0.05。极显著水平为P<0.01。图件采用origin 9、CorelDRAW 2018和Sufer 15等绘图软件完成。
2结果2.1OTC的增温效应
本次提取监测站2020年5月份到12月份土壤表层(5 cm)和深层(20 cm)土壤温度以及OTC内气温和相对湿度数据进行分析(由于在线设备故障,CROWN Ⅰ全年数据和CROWN Ⅱ 5月份土壤表层温度丢失)。此外为了更加直观体现增温变化,特将9月16日研究时段的相关数据进行分析。
从表1可以看出,OTC增温箱的土壤平均温度都要高于对照组,其中研究时段的5 cm土壤平均温度最高,为24.01℃,日平均季节的20 cm土壤温度最低,为19.15℃。OTC增温箱和对照组的5 cm土壤温度都要高于20 cm土壤温度。由图4可以看出,不管表层(5 cm)还是深层(20 cm),增温组和对照组土壤温度都具有明显的季节性特征。增温组表层土壤温度比对照组平均高0.13℃,深层土壤温度比对照组平均高0.35℃。10月份土壤深层温度变化幅度明显高于土壤表层,最高温度增幅为1.6℃。从研究时段看,不同时段的增温幅度存在明显差异(图4)。
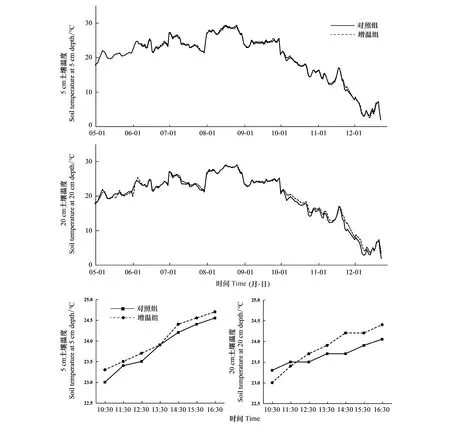
图4 芦苇增温站(CROWN Ⅱ)增温组和对照组土壤温度日平均季节变化和典型晴朗天气研究时段日变化 Fig.4 Average daily seasonal variation of soil temperature in the warming group and the control group and the diurnal variation in the study period of typical fine weather at CROWN II
从表1可知研究时段的气温显著高于日平均季节,其中研究时段OTC增温箱气温最高,均值为30.06℃,日平均季节对照组最低为19.46℃。图5显示增温组和对照组5—12月份的气温同样具有明显的季节性特征变化。增温组和对照组的气温变化幅度不大,增温组平均温度比对照组高0.51℃,且最高气温都出现在八月份(增温组为(28.91±0.06)℃,对照组为(28.51±0.05)℃)。从研究时段看(图5),气温和相对湿度均呈现良好的变化规律。增温组的平均气温比对照组高2.06℃,表明OTC具有明显的增温效果,但相对湿度正好相反,增温组的平均相对湿度比对照组低7.03%。
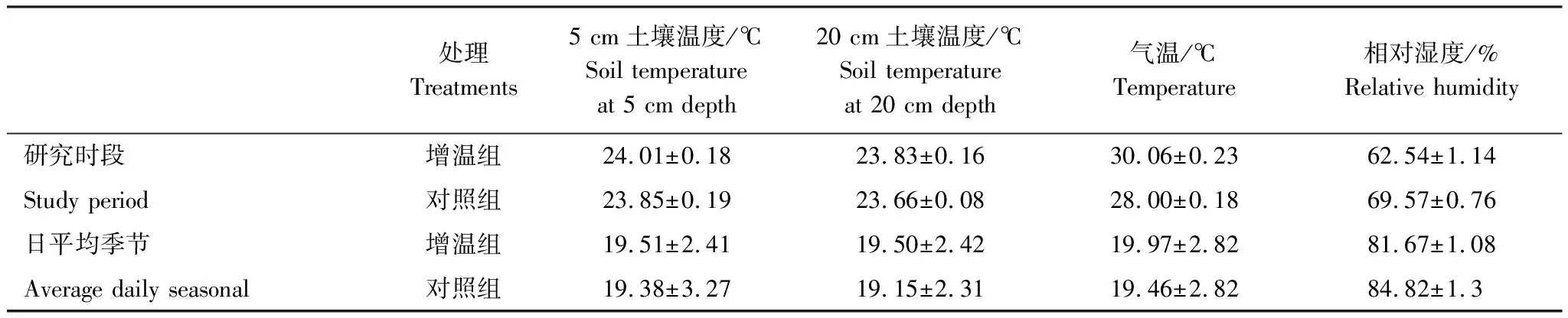
表1 增温组和对照组环境因子对比(平均值±标准误)
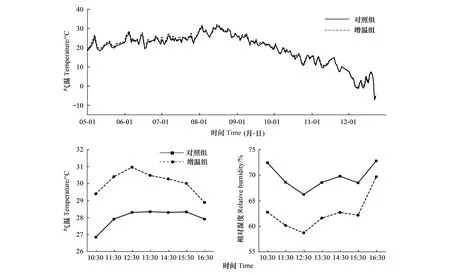
图5 CROWN Ⅱ增温组和对照组气温日平均季节变化和典型晴朗天气下研究时段日变化Fig.5 Diurnal mean seasonal variation of air temperature of the warming group and the control group and diurnal variation in the study period under a sunny day at CROWN II
2.2 增温对瞬时气体交换参数的影响
从图6看出,互花米草和芦苇的Pn日变化趋势大体相似,都表现为双峰型。在上午10:30左右四组植物的净光合速率达到最大,其中互花米草增温组最大,达到22.83 μmol m-2s-1。净光合速率除了芦苇增温组在上午11:30略有上升之外,其他都是逐渐下降,随后小幅度的增加后又逐渐下降。除了芦苇增温组,其他三组植物的气孔导度Gs都是在10:30达到一天最大值,其中芦苇对照组的Gs最大,达到0.32 mol m-2s-1。互花米草Gs要远小于芦苇,且变化幅度较小。 芦苇蒸腾速率Tr的日变化幅度相对较大,对照组和增温组的变化趋势大体相似,除了增温组的Tr在上午11:30时小幅度增加外,两者都是逐渐下降到最小值之后稍微升高后又开始下降。但对照组的Tr始终大于增温组。互花米草的Tr变化相对平稳,总体呈下降趋势。与芦苇不同的是,互花米草增温组的Tr和Gs始终大于对照组。芦苇的胞间CO2浓度Ci在一天中未呈规律性变化,且均显著大于互花米草(P<0.05),但互花米草的Ci规律性较好,增温组的Ci较稳定,保持在150 μmol/mol上下波动,在上午11:40之前高于对照组之外,其他时间都低于对照组。芦苇的水分利用率WUE显著低于互花米草,并且变化幅度不大,在2.28—2.48 mmol/mol之间波动,对照组的WUE显著高于增温组(P<0.05)。互花米草增温组和对照组的WUE变化规律均呈不规则的“W”形变化。增温组的WUE整体上高于对照组,两者最大值都出现在14:30,分别为6.27 mmol/mol和5.75 mmol/mol。
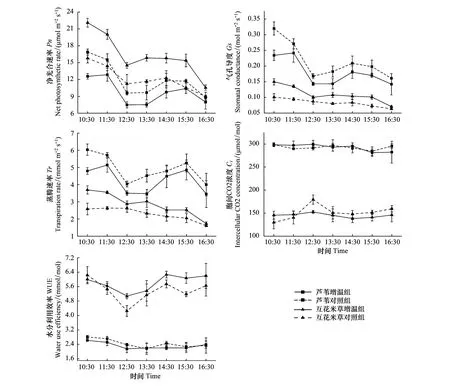
图6 日调查下芦苇和互花米草各参数比较Fig.6 Comparison of photosynthetic characteristic parameters between Phragmites australis and Spartina alterniflora
2.3 瞬时气体交换参数间相关性分析
从表2得知,除芦苇对照组的WUE外,Pn与Tr和Gs显著相关(r>0.77,P<0.05),但与WUE和Ci相关性不显著(r=0.74,P>0.05)。
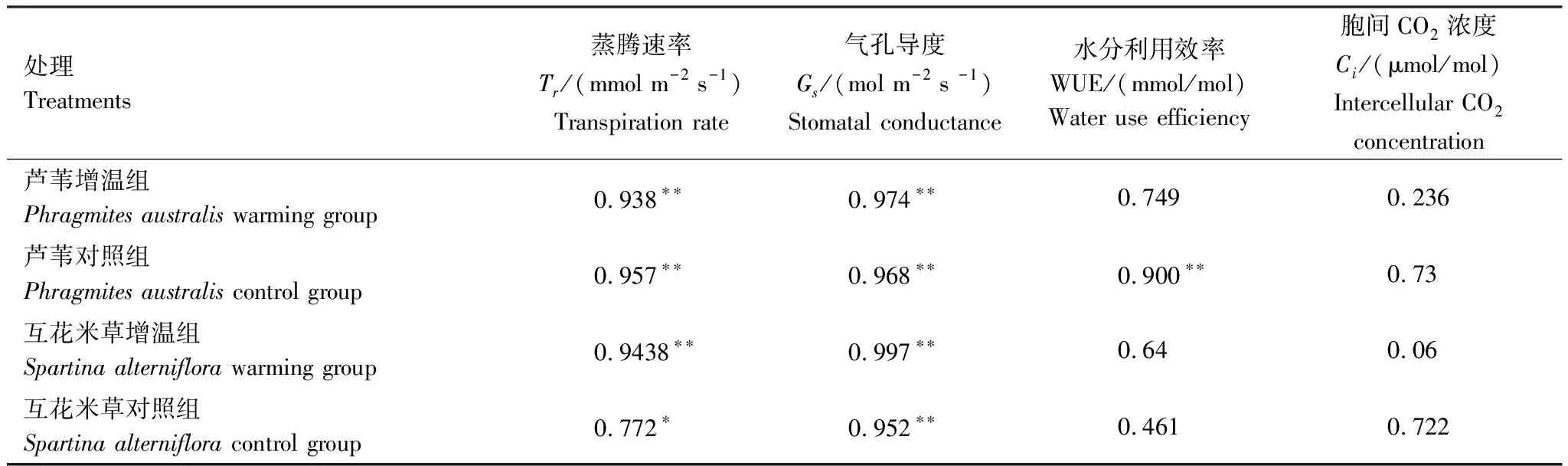
表2 净光合速率(Pn)与其他光合参数相关系数
2.4 增温对光响应曲线及响应参数的影响
图7显示增温显著地影响了芦苇和互花米草的光合响应过程。增温组互花米草的光响应曲线高于对照组,且在光合有效辐射较低时互花米草的净光合速率要明显高于对照组,而芦苇增温组的光响应曲线则低于对照组,但在光合有效辐射较低时两者的净光合速率变化幅度相似,当光合有效辐射达到200 μmol m-2s-1时,表现为增温组和对照组之间的显著差异。
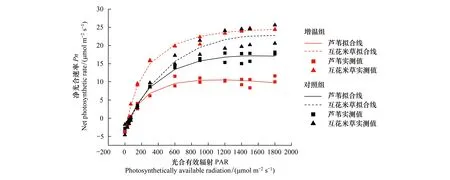
图7 芦苇和互花米草的光响应曲线Fig.7 Light response curves of Phragmites australis and Spartina alterniflora
增温条件下,芦苇的各项光合参数都有所降低,其中Pnmax和LSP变化幅度较大,分别下降了35.2%和41.6%。但是互花米草除了LCP降低了45.3%外,其他各项参数都有了不同程度的提升,其中AQY增加了114%(表3)。
3 讨论
植物的光合生理特性对植物的生长发育至关重要,并且与所处环境温度密切相关。一方面相同的植物生理特性对不同的温度表现出不同的响应特征;另一方面,不同的植物光合生理特征对相同的温度可能存在不同的响应规律,造成这种差异的原因可能是植物对所处环境温度的敏感性不同[10]。上述研究结果表明该区滨海湿地芦苇和互花米草对增温响应程度和方向存在显著的差别,特别是不同植物响应的方向存在相反的现象,其具体情况讨论如下。
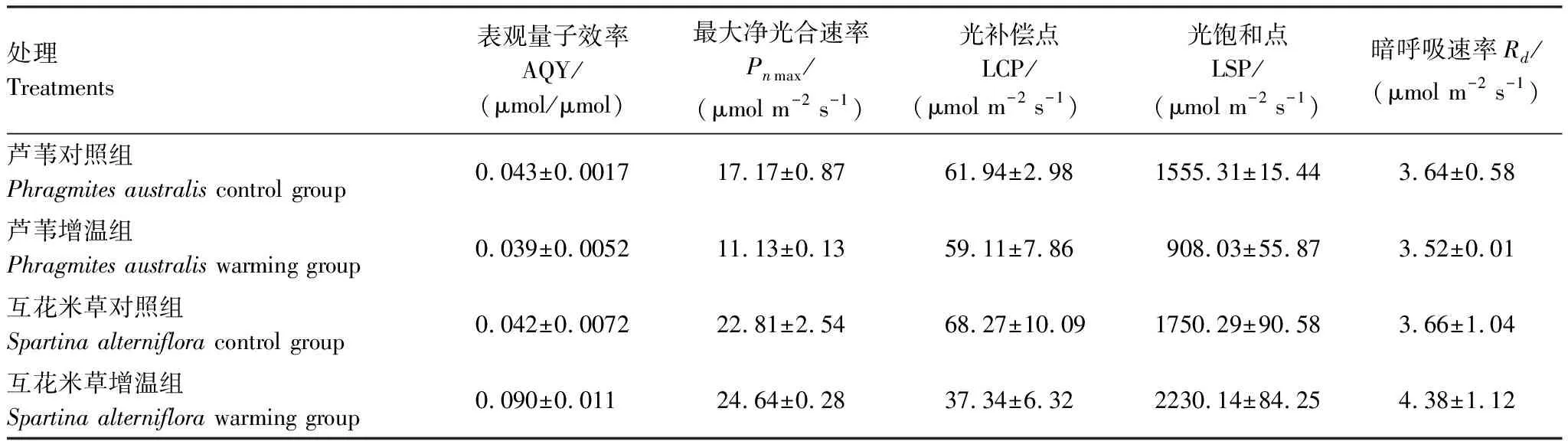
表3 不同生长环境下芦苇和互花米草主要光合参数对增温的响应
3.1 开顶箱的增温效果及其对湿度影响
OTC开顶箱可以降低内部空气流动,保持住内部热量,由此起到增温效果。从日平均季节变化来看,OTC内空气温度平均升高了0.51℃,而研究时段内OTC内空气增高幅度更大,为2.06℃,增温效果与先前文献报道的类似装置相似[29,31,38]。
深层(20 cm)土壤温度平均增温幅度为0.35℃,显著高于表层(5 cm)土壤0.13℃的增温幅度(表1),该增温幅度略小于青海小泊湖的同类增温效果[29]。从研究时段统计数据来看,深层土壤增温效果亦略优于表层土壤(表1)。这可能是由于靠近海边,研究站在潮汐的影响下,间歇性的淹水降低了OTC对表层土壤的增温效果。
相对湿度主要与空气中水蒸气含量和温度有关,空气中水蒸气含量差异不大,所以空气相对湿度主要受温度控制[39],从日平均季节变化和研究时段统计结果发现OTC内湿度分别下降了3.15%和7.03%(表1)。由于OTC开顶箱可以有效保持光照强度幅射下来的热量,使箱内水蒸气蒸发,因此PAR在一定程度上也影响了OTC内气温和相对湿度的变化。
3.2 芦苇和互花米草瞬时气体交换参数对模拟增温的响应
温度是影响植物光合作用的主要因素,控制着生态系统中许多生物化学反应速率,以及绝大部分生物学过程[40—41]。本研究芦苇和互花米草气体交换参数Pn、Gs、Tr和WUE等光合性能存在相似性,但对模拟增温的响应方向是相反的(图6),表现出一定程度的复杂性和不确定性。净光合速率Pn是指光合作用产生的糖类减去呼吸作用消耗的糖类的速率,或者说净光合速率是指植物光合作用积累的有机物,是总光合速率减去呼吸速率的值。芦苇和互花米草光合性能相似性可从图6结果观测到,即表现为在测量时间段内,芦苇和互花米草瞬时净光合速率Pn曲线都为双峰曲线型,其原因是中午温度过高,植物体内缺水,植物为实现自我保护,气孔导度关闭导致植物光合作用下降,进入“午休”状态,下午气温和光合有效辐射下降后,又达到了植物的适宜环境,气孔打开,光合作用又开始增加。Pn曲线Gs分布形状相吻合(图6),以及两者相关性很高(r>0.95,P<0.01)(表2)均进一步证明了上述推断。马加芳等的研究也证明了这一现象[42]。本研究在模拟增温3年后的观测结果发现互花米草各项光合参数Pn、Gs、Tr和WUE对增温的响应符合一般性认识,其各项生理参数值呈上升趋势(图6),生理机能增强,促进了光合作用。该现象的主要驱动机制有二:其一是增温可通过加速新陈代谢增加植物的光合能力或通过较高的矿物质分解作用增强植物对矿物营养的吸收,从而促进植物的生长[43];其二是,增温也可能促进植物细胞分裂,增强植物从土壤获取营养物质的能力,以此促进植物生长,提高植物净光合速率。然而,不同于互花米草对增温的响应,在为期3年的增温处理样地中芦苇的光合参数Pn、Gs、Tr和WUE等均呈显著下降趋势(图6),表明增温抑制了芦苇的光合生理活动。这一结果有悖于光合性能对增温响应的一般性认识,可能的原因是Rubisco酶含量和活性在长期持续的增温条件下降低[44],为保持光合作用动态平衡,减少气孔张度,致使Tr下降、WUE下降,从而导致Pn下降。特别是在长期增温条件下,加速了植物对土壤中的氮的吸收,使土壤氮含量耗尽[45],由此导致叶氮含量及光合氮素利用效率下降,降低植物的比叶重,进而影响了植物的光合作用[10]。杨淑慧等于2012年对比了崇明东滩围垦湿地芦苇增温1年和2年后的结果发现增温第一年净光合速率增高,而第2年却下降[46]。亦即是说植物光合性能对增温响应存在短期效应和长期效应的不同,因此持续不断的观测研究是必要的。需要特别指出的是,互花米草生长过程中消耗土壤氮及矿物质营养的速率较芦苇慢,且互花米草作为C4植物,增温条件下光合作用酶对温度耐受性要强于芦苇,因此增温提高了互花米草酶的活性,使互花米草光合作用进一步加强。
WUE是Pn与Tr的比值,指生态系统单位重量水所制造的干物质量,是表示植被水分吸收过程利用效率的一个指标。图6显示增温处理样地中互花米草的WUE指标升高,而芦苇的WUE指标下降,进一步说明互花米草在水分缺失的情况下仍能保持较高的生产力,这与樊同在崇明滨海湿地实验中得到相同的结论[47]。这说明增温条件下互花米草对环境的适应能力和节水能力要强于芦苇。气孔是植物叶片与外界气体交换的主要通道,它是影响光合作用,植物呼吸作用及蒸腾作用的主要因素[48]。芦苇和互花米草的Pn和Gs、Tr呈现高度相关性,说明温度通过影响气孔导度进而影响了植物的光合作用,但不能单纯的将气孔导度作为判断Pn气孔限制因素的唯一标准,许大全认为可以将胞间CO2浓度Ci的下降变化方向作为气孔因素是否导致植物净光合速率变化的重要依据,许大全特别强调指出胞间CO2分压增加是光合作用非气孔限制的最可靠的判据[49]。其规则是:当Pn下降,Ci下降,此时气孔因素是限制净光合速率的主要原因;当Pn下降,Ci升高或保持不变,此时非气孔因素是限制净光合速率的主要原因。从本研究日变化观测来看,在上午10:30—12:00时间段观测的四组不同处理的植物净光合速率Pn与胞间CO2浓度Ci对比结果可以发现,当Pn下降时,Gs也下降,但其相应的Ci有所增高(如图6的互花米草的控制处理)或不变(如图6的芦苇)。如果从增温和控制处理对比来看,一方面增温处理的互花米草Pn增加,Gs增加,而Ci是减少的;另一方面芦苇的Pn下降,但Gs是增高的,且Ci未呈显著的变化。这些均说明叶片的叶肉细胞活性比气孔导度更重要。因此,通过本研究观测的结果,可以比较可靠地判定非气孔限制因素是Pn下降的原因,即叶肉细胞光合活性酶的降低可能是Pn下降的主要驱动机制[50]。
3.3 光响应曲线及响应参数对模拟增温的响应
光响应曲线反应了植物光合作用随着光合有效辐射(PAR)变化而变化的趋势。表观量子效率AQY和光补偿点LCP反映了植物对弱光的利用情况。其中AQY是光合生理生态研究中广泛使用的参数,是指在不考虑叶片的光反射和投射损失(一般为15%左右)、不按照光合机构实际吸收的光量子数,而是按照入射的光量子数计算得到的量子效率,即为表观量子效率,其度量值为光响应曲线在弱光阶段的斜率。AQY越小,表示植物利用弱光能力越弱。而LCP是指当植物通过光合作用制造的有机物质与呼吸作用消耗的物质相平衡时的光照强度,LCP越大则表示植物利用弱光能力越弱。增温条件下,互花米草的AQY值增高,而芦苇的同名参数保持微小的变化,但两种植物的LCP对增温的响应均表现为下降(表3),说明增温增强了芦苇和互花米草利用弱光的能力,芦苇和互花米草的耐阴性都有所加强。
在一定的光强范围内,植物的光合速率随光照强度的上升而增大,当光照强度上升到某一数值之后,光合速率不再继续提高时的光照强度值为光饱和点(LSP)。LSP表征植物对强光的适应能力。芦苇和互花米草的LSP在增温条件下发生了相反的变化,可能是由于互花米草属于C4植物,芦苇属于C3植物,使得在相同环境下,互花米草对PAR有更好的适应性,具有更高的光合速率和LSP,与赵广琦等的研究结果相似[51],这是互花米草能成为入侵物种的另一个重要因素。从图7和表2可以看出,增温降低了芦苇的Pnmax和LSP,意味着芦苇适应强光的能力下降,导致光合作用能力下降,而在增温条件下,互花米草的Pnmax和LSP均有大幅度增加,说明互花米草更容易适应强光,即使在中午时段互花米草增温组的净光合速率高于其他三组的主要原因。
暗呼吸速率Rd指示着植物叶片新陈代谢作用的强弱,是指植物叶片在光照强度为零时,单位时间、单位叶面积通过呼吸作用消耗氧气和有机物生成的二氧化碳的量。Rd数值越大指示植物新陈代谢越强,对贮存的碳化物的消耗加快。本研究发现增温处理芦苇的Rd下降(表3),碳化物消耗减慢,新陈代谢功能减弱的现象从另一个侧面也证明了芦苇存在明显的增温胁迫,从而限制了芦苇光合性能和碳汇能力。而增温处理的互花米草的Rd上升(表3),即增温使碳化物消耗显著增加,这一现象合乎植物暗呼吸速率对增温响应的一般规律。虽然芦苇和互花米草暗呼吸速率对增温响应表现为前者消耗有机质速率减小,而后者增大,但其暗呼吸量不足以抵消光合速率变化的速率,这点可从净光合速率对增温响应得到清楚的解释(图6)。因此,可以肯定地判断芦苇受到增温的胁迫,在减缓了新陈代谢过程同时也减弱了光合作用,而作为C4植物互花米草对增温有较好的适应性,且表现出更好的光合作用能力和固碳能力,同时不可避免地需消耗更多的光合作用产物。因此,从本研究结果可以推论,植物较高的暗呼吸速率,可以指示较强的光合性能,反之亦然。然而,要从理论上深入的解释暗呼吸速率对光合性能指示意义还有待进一步的研究工作。
4 结语
植物的光合特性是由众多的过程和因子交织在一起共同调控的,对增温的响应机制比较复杂,仅从某一方面,某一时刻研究植物在增温条件下的响应存在片面性。特别是生态系统植物光合性能对增温响应存在短期效应和长期效应的区别,同时也存在植物在不同生长季节的区别,利用短期、瞬时的光合数据去预测未来生态系统光合性能演化趋势需十分谨慎。本研究的不足之处有:(1)在日调查只选取了三片叶子作为重复实验,重复性较少;(2)测光合日变化一般应在一早光强较低时就开始,本研究的光合日变化数据缺了早晨一块,不够完善;(3)光响应曲线的测定时间未考虑上下午环境因素的影响,没有做到时间统一;(4)本研究测试的时候是生长季偏后期,光合作用的大小也可能与植物或土壤的营养元素变化相关,而非仅仅是与增温相关。这些都使研究结果的外延推测存在局限性。此外,很多研究已证明植物的生理特性对增温的响应会随着增温作用时间的变化而变化,并且在植物不同生长阶段植物对温度的敏感不同。为揭示滨海湿地芦苇和互花米草光合特性以及碳汇功能在未来气候持续变暖的情况下的响应,研究其响应机制,有必要设计更完善的实验方案,特别是要加强季节变化和年际变化等不同时间尺度的连续观测,这也是下一步需要注意的问题。只有这样,才能为未来气候变暖情况下预测植物的竞争力、适应性以及生态系统碳汇功能的变化提供更为可靠的科学依据。
猜你喜欢
农业工程学报(2022年4期)2022-04-24
海外文摘·文学版(2022年4期)2022-04-14
黄河之声(2021年19期)2021-02-24
农业与技术(2020年12期)2020-07-04
岁月(2018年2期)2018-02-28
中国诗歌(2017年12期)2017-11-15
新课程·下旬(2017年7期)2017-08-14
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
